Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 3
Aim: The role of alkaline phosphatase in cell metabolism and regulation is essential and cuts across cell proliferation, division, cell death, membrane transport and DNA cleavage. This study addresses the role and importance of alkaline phosphates (ALP) and Placenta Alkaline phosphatase (PLAP) in the cells of germinal epithelium of male Rat testes following induced lead (Pb) toxicity and rejuvenation by Selenium (Se) and Zinc (Zn) treatment.
Method: Adult Sprague-Dawley Rats (males) were divided into 4 groups of 15 animals each and were treated as thus; Group 1: Normal saline and serves as the control, Group 2: 100 mg/Kg BW of Pb only, Group 3: 100 mg/Kg BW of Pb and 2.25 mg/Kg each of Se and Zn and group 4 received 2.25 mg.Kg each of Se and Zn only. The duration of treatment was 56 days following which the animals were sacrificed by the 57th day and testes fixed in Bouin’s fluid.
Result: Pb induced toxicity could be apoptotic involving ROS activation of NO-dependent apoptotic pathway or necrotic involving a wide range damage by ROS induced lipid peroxidation. Selenium and zinc treatment ameliorated the damage induced by lead toxicity in the germinal epithelium.
Conclusion: ALP is involved in intrinsic regulation of DNA cleavage in apoptosis by functioning similar to endonucleases and also regulates membrane transport in perioxidated biomembranes. PLAP expression is minimal as tumorgenesis was not recorded, although Pb treatment showed signs of epithelium of endothelial cancer signalling, which was ameliorated by Se+Zn treatment.
<An important feature of the phosphatases is that they are capable of removing phosphate groups from biomolecules, a function similar to those of endonucleases and opposite that of kinases. There are four isozymes of humans’ Alkaline Phosphatase (ALP); these include lower ALP (L-ALP), corticosteroids ALP (C-ALP), Bone ALP and placenta- ALP (P-ALP). They can be detected by isoelectric point focusing on the germ cells. ALP has also been described as an important enzyme in membrane transport [1,2]. The isoform of ALP includes lower ALP (L-ALP), corticosteroids ALP (C-ALP), Bone ALP and placenta-ALP (P-ALP) [3]. It is well known that testicular germ tumor cells are capable of expressing early embryonic and extra-embryonic proteins. The gene coding for ALP and PLAP are usually not active until turned on in tumorgenesis [4,5]. The PLAP are products of early migrating primordial germ cells during development of the human embryo, whereas in adult this protein is expressed in trace amount in the testes and Thymus [6-8]. Recently, PLAP has been suggested to be appropriate for marking tumorgenic cells, and like the other species of ALP. It can be produced by cells of the seminiferous tubules during induced proliferation and degenerative processes such as those involving DNA damage [9].
The primary product of Pb toxicity is cell death, which could be either apoptotic or necrotic depending on the extent of the assault and bioactivation of radical scavengers such as Vitamin C and Vitamin E, Selenium and Zinc. Two different pathways have since been identified as routes to cell death in Pb toxicity: (1) Mitochondria signalling pathway involving Cathepsin D and production of Nitric Oxide (NO) which is capable of inducing DNA breakage [10] (2) Cytoplasmic pathway involving p53 signalling and apoptosis [11].
A necrotic pathway is obtainable in high concentration involving formation of oxygen radicals. The studies of Favilla et al. [12] and Koshida et al. [13] suggest that a similar event of DNA cleavage precedes apoptosis and necrosis. Two patterns of DNA cleavage have been described using a multi technique approach of In situ Endo Labelling (ISEL) to identify necrosis as 3’OH-5’P cleavage and In situ Nick Translation (ISNT) to identify apoptosis as 3’P-5’OH cleavage [14]. The DNA cleavage described above has been identified to be a function of 10 (Ten) different type of endonucleases and ALP/ PLAP [15]. In a normal tissue, ALP expression is active in membrane transport, and is regulatory in the tissue of the body by inducing DNA cleavage while PLAP is regulatory in embryonic systems [16]. Not all cleavage patterns are regulated and some may lead to tumorgenesis. Several studies have described tumorgenesis in Pb toxicity, thus PLAP is expressed in such tumorgenic cell lines [17]. Se+Zn, if in trace amount, can alter the toxicity of Pb by activating radical scavengers and vitaminsas well as inhibiting several cell death and degenerative pathways. This study examines the expression level of PLAP and ALP in Pb treated germ cells and possible ameliorative or cytoprotective effects of Se+Zn treatment.
60 F1 generation adult Sprague-Dawley rats were used. The animals were procured and kept in the animal holding facility of the Osun State University and allowed to acclimatize. The animals were then divided into 4 groups of 15 animals each. The animal handling protocol followed the Helsinki Convention on animals use for research. Group 1 (control) received 10 ml/kg normal saline, group 2 received 100 mg/Kg BW of lead acetate (Pb), group 3 received 100 mg/Kg of Pb, 2.25 mg/Kg of Se and 2.25 mg/Kg of Zn. The duration of treatment was 56 days for all groups (56 days represents the complete cycle for spermatogenesis; to ensure that each of the stages have been affected by the treatments). The animals were sacrificed on the 57th day and the testes were dissected to obtain the seminiferous tubules. The tissues were immediately fixed in Bouin’s fluid (histology) and Formolcalcium (Immunohistochemistry) for 24 hours. Tissues for histology were later transferred into a change of Bouin’s fluid for another 24 hours. Tissue processing was done to obtain paraffin wax embedded sections using the method of Ng et al. [18] [Se: Sodium Selenium and Zn: Chelated Zinc].
Histology
The embedded tissues were sectioned to obtain 7 μm thick sections for routine histological staining in Hematoxylin and Eosin (H&E) using the method of Denison et al. [19].
Immunohistochemistry
The paraffin wax embedded sections were mounted on a glass slide in preparation for antigen retrieval where the slides were immersed in urea overnight and then placed in a microwave for 45 minutes to re-activate the antigens and proteins in the tissue sections. Primary antibody treatment involved treating the sections with biotinylated goat serum for one hour following which the sections were transferred to 1% bovine serum albumin (BSA) to block non-specific protein reactions. Secondary treatment involved the use of diluted anti-ALP and Anti- PLAP on the pre-treated sections for one hour. The immunopositive reactions were developed using a polymer 3’3’ Diaminobenzidine Tetrachloride (DAB) with colour intensification involving the use of mathenamine silver kit. The sections were counterstained in Hematoxylin and treated in 1% acid alcohol (freshly prepared).
Transformation
Methenamine silver intensification was used on the immunoperoxidase preparation after the peroxidase/H2O2/DAB reaction has been carried out to give a brown deposit. The sections were then counterstained in Hematoxylin. The counterstained sections were washed in running tap water, thoroughly rinsed in distilled water, and placed in preheated methenamine silver solution at 60°C for five minutes. Although it could be occasionally longer if the intensification had been carried out at room temperature. In this study, to further increase the clarity, Hematoxylin was removed from counterstained nuclei with 1% acid alcohol before the silver intensification was carried out. The composition of the stock solution was 0.125% silver nitrate in 1.5% hexamine. The solution was stored at 4°C. Prior to use, 2 ml of 5% tetraborate was added to 50 ml of the stock silver solution giving a pH of 8.0, which was then filtered into a coupling jar and protected from sunlight.
The general morphology of the testes in Hematoxylin and Eosin staining shown in Figure 1A (control), Figure 1B (Pb treatment), Figure 1C (Se+Zn treatment) and Figure 1D (Se+Zn treatment following Pb treatment). This shows that Pb treatment (Figure 2B) induced degeneration in the spermatogenic cells while treatment with Se+Zn reduced the extent of such degeneration. Immunohistochemical localization involves the antigen retrieval method and an immunoperoxidase reaction involving the Peroxidase Anti Peroxidase (PAP) method. Brownish immunodeposition (positivity) resulting from DAB were observed to be heavy around the luminal surface of the seminiferous tubules (arrow head and ‡) (Figure 2A and 2C). Histocomparative analysis reveals that ALP expression was highest in Pb+Se+Zn treatment group (2C), followed by the control (2A). The Se+Zn only (Group 4) showed expression in its thick luminal surface (Figure 2D) but lower than those observed in the control, the least ALP expression was observed in the Pb treatment group (2B). This is evident that ALP expression was increased in the Pb+Se+Zn (Figure 2C) treatment due to the effect on membrane transport and exchange rather than cell regulation. The reduced ALP immunodeposition in Se+Zn synergy treatment only Figure 2D showed that cell proliferation observed in this group was due to Se+Zn treatment irrespective of ALP production. Hence, ALP is linked to cell survival in Pb+Se+Zn treatment rather than cell proliferation in Se+Zn treatment. PLAP expression was minimal and is seen only in few endothelial cells of the control (‡ in Figure 3A). The PLAP immunodeposition was entirely limited to the region around the basement membrane, most likely characteristic of the dividing spermatogonia at the base of the Sertoli cell projections. This is an indication that Pb treatment can signal tumorgenesis in germ cells of the epithelium. PLAP positivity was not observed in Se+Zn treated group (Figure 3D), thus implying Se+Zn does not trigger tumorgenesis and holds the possibility of suppressing germ cell carcinogen. This can be further elucidated by comparing Figure 3C (Pb+Se+Zn treatment) and Figure 3B (Pb treatment only); the entire immunopositive region around the basement membrane of the Pb treated group is absent in the Pb+Se+Zn treatment which indicates that Se+Zn synergy can hold certain tumor suppressor effects.
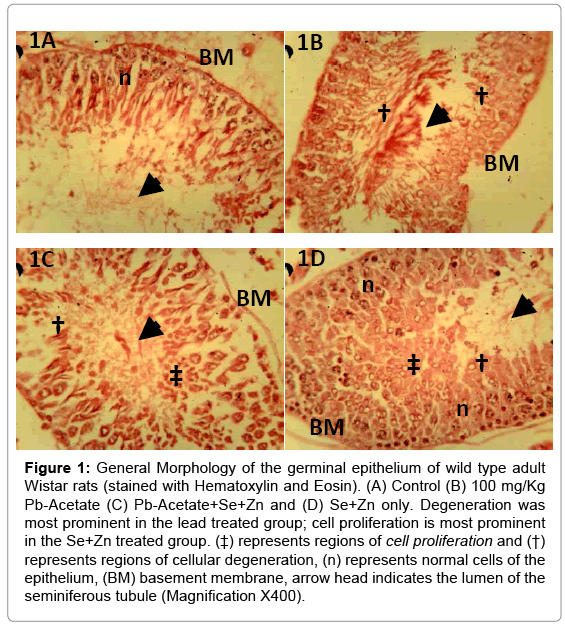
Figure 1: General Morphology of the germinal epithelium of wild type adult Wistar rats (stained with Hematoxylin and Eosin). (A) Control (B) 100 mg/Kg Pb-Acetate (C) Pb-Acetate+Se+Zn and (D) Se+Zn only. Degeneration was most prominent in the lead treated group; cell proliferation is most prominent in the Se+Zn treated group. (‡) represents regions of cell proliferation and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Magnification X400).
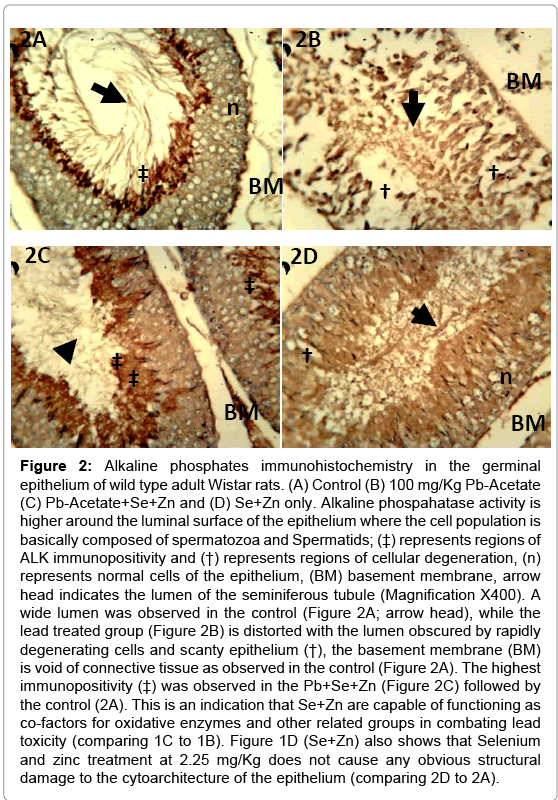
Figure 2: Alkaline phosphates immunohistochemistry in the germinal epithelium of wild type adult Wistar rats. (A) Control (B) 100 mg/Kg Pb-Acetate (C) Pb-Acetate+Se+Zn and (D) Se+Zn only. Alkaline phospahatase activity is higher around the luminal surface of the epithelium where the cell population is basically composed of spermatozoa and Spermatids; (‡) represents regions of ALK immunopositivity and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Magnification X400). A wide lumen was observed in the control (Figure 2A; arrow head), while the lead treated group (Figure 2B) is distorted with the lumen obscured by rapidly degenerating cells and scanty epithelium (†), the basement membrane (BM) is void of connective tissue as observed in the control (Figure 2A). The highest immunopositivity (‡) was observed in the Pb+Se+Zn (Figure 2C) followed by the control (2A). This is an indication that Se+Zn are capable of functioning as co-factors for oxidative enzymes and other related groups in combating lead toxicity (comparing 1C to 1B). Figure 1D (Se+Zn) also shows that Selenium and zinc treatment at 2.25 mg/Kg does not cause any obvious structural damage to the cytoarchitecture of the epithelium (comparing 2D to 2A).
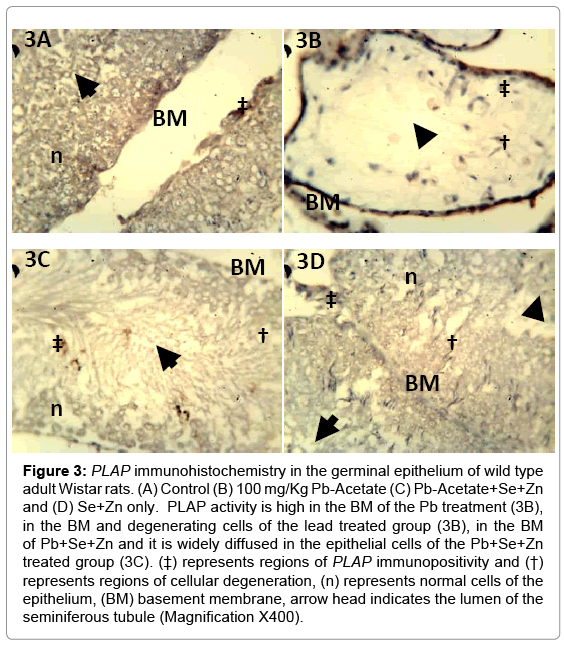
Figure 3: PLAP immunohistochemistry in the germinal epithelium of wild type adult Wistar rats. (A) Control (B) 100 mg/Kg Pb-Acetate (C) Pb-Acetate+Se+Zn and (D) Se+Zn only. PLAP activity is high in the BM of the Pb treatment (3B), in the BM and degenerating cells of the lead treated group (3B), in the BM of Pb+Se+Zn and it is widely diffused in the epithelial cells of the Pb+Se+Zn treated group (3C). (‡) represents regions of PLAP immunopositivity and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Magnification X400).
Membrane damage and DNA cleavage are both products of oxidative stress; resultant of the ability of Pb to inhibit metallo-enzyme Cytochrome C Oxidase (CcOX) - a membrane bound mitochondria enzyme and a member of the Cytochrome P450 super family that forms the Complex V of the electron transport chain [20]. When such inhibition occurs, it will generate ROS at Complex I and IIIa [21]. The ROS thus formed reacts with nitrogen containing compound to form NO and itself induces peroxidation of lipids in biomembranes. This tilt the cells in a path of apoptosis hanging the balance of tumorgenesis should in case it escapes the Go phase of cell death [22]. The role of phosphatases are important in membrane dysfunction as the enzyme itself is membrane bounded and required for membrane transport; so also in apoptosis or nuclear cleavage as ALP function like the endonucleases to induce DNA cleavage (3’OH-5’P or 3’P-5’OH) [23,24]. Lead, being a mitochondria poison is capable of inducing cell death by oxidative stress and has been described as tumorgenic in other studies. Testicular disorders are very rare, thus because of the rarity and diverse morphology they present a diagnostic challenge. Some of the effects of lead exposure include male infertility; our ultimate aim is to describe ALP and PLAP as markers for spermatogenic linage dysregulation and how they can be used as indicators for membrane transport at the luminal surface (ALP) and basement membrane cell signalling (PLAP) both in toxicity and Se-Zn synergy treatment. The expression of ALP in this study represents two important cellular processes. The first is the role of ALP in membrane transport which can be seen as the intensity of brownish deposit on the luminal surface of the germinal epithelium. Our study shows that membrane transport is highest in Se+Zn treatment (Figure 2D), higher than that observed in the control group (Figure 2A). The lowest ALP immunopositivity was seen in lead (Pb) treated group (group 1) (Figure 2B) while some ameliorative effects were equally observed in group 3 (Figure 2C). This shows a reversal effect of Se+Zn synergy treatment on Pb toxicity. Thus, the effect of Pb on membrane transport in the spermatogenic cell line can be said to be detrimental as it reduces the ALP expression level, while Se+Zn treatment can relief such toxicity by increasing ALP activity at the luminal cell layer. Based on our initial discussion on the role of lead induced oxidative stress on inducing DNA cleavage, a second role of ALP can be depicted as it acts like the endonucleases by removing phosphate groups from the phosphate sugar backbone of DNA. The process of such dephosphorylation causes DNA cleavage which then leads to apoptosis. The limitation of this study is that no specific staining was done for DNA cleavage to determine its exact pattern in this study; however, the expression pattern can be described as phasic such that a first line of defence is to regulate membrane transport, followed by a second line of DNA cleavage to prevent irregular cell division and tumorgenesis.
A balance was seen in toxicity of Pb and counter toxicity by Se+Zn although the activities of Se+Zn synergy are seen to involve the inhibition of pathways involved in ROS activation, and improving the viability of metalloenzymes like ALP and CcOX as observed in the ALP (Figure 3A and 3C) where membrane activity increased following Se+Zn in Pb toxicity similar to the findings of Choudhury et al. [25]. The studies of Jeong et al. [26] suggested selenium attenuated the cadmium induced testicular damage in rats. Biochemical evaluation from the studies showed increased level of ALP in the treatment with Se and a reduction in the Pb treated group; although no immunolabelling was done. Other studies involving the use of Se+Zn synergy demonstrated the efficacy of this synergy against Mercury induced toxicity, the result showed increased activities of AST, ALT and Alkaline phosphate, the effects was also consistent for hepatic, kidney and neural tissues [27,28]. Zn and Se are complementary in function, aside having synergistic effects when co-administered, as selenium supplementations have been seen to improve zinc levels and anti-oxidant value of blood [29-31] and counter oxidative stress better [16]; primary marker is improved membrane transport and metalloenzymes activity (CcOX & ALP) as selenium inhibits oxidative stress, improves signal transport and differentiation in bone marrow cells. Se+Zn will also generally improve the blood metabolic profile [32].
Pb induced toxicity could be apoptotic involving ROS activation of NO-dependent apoptotic pathway or necrotic involving a wide range damage by ROS induced lipid peroxidation. In both cases ALP is involved in intrinsic regulation of DNA cleavage in apoptosis by functioning similar to endonucleases and also regulates membrane transport in perioxidated biomembranes. PLAP expression is minimal as tumorgenesis was not recorded. Although Pb treatment showed signs of epithelium of endothelial cancer signalling, this was ameliorated by Se+Zn treatment.