Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2015) Volume 6, Issue 4
The study reports the results of molecular characterization of the Internal Transcribed Spacer (ITS) of ribosomal DNA of 3 Monogenean species using polymerase chain reaction (PCR), nucleotide sequencing and construction of phylogenetic trees from different fish hosts of Kashmir. The present study shows that the size of the amplified product is 873bp long for D. kashmirensis, 1120bp long in D. aegyptensis and 687bp long in D. guptai revealing that there are intraspecific differences in their base pair lengths. Guanine and Cytocine (G+C) content of three Diplozoon species was found nearly constant for three species i.e., 47% (D. kashmirensis); 47% (D. aegyptensis) and 48% (D. guptai), this GC richness contributes to physical attributes of RNA structures, as there is correlation between GC content and optimal growth temperature. An important observation during the present study has been noticed that Schizothorax niger is infected by all the three species of Diplozoidae; D. kashmirensis; D. aegyptensis and D. guptai, but when all six fishes were collected simultaneously, parasitism by all the parasite species was never observed. Phylogenetic trees Maximum Parsimony (MP), Maximum Likelihood (ML) and Neighbor Joining (NJ) showed that D. kashmirensis and D. aegyptensis share a common host Carassius carassius and S. niger.
Keywords: Diplozoon, Ribosomal DNA, Schizothorax, Kashmir,Phylogenetic trees.
Monogeneans belonging to the Diplozoidae are common parasites on the gills of cyprinid fish. The life cycle is direct, including freeswimming oncomiracidia, larval stage (diporpa) and adult. Two larvae (diporpae) permanently fuse into a pair to form the sexually maturated adult. In the adult, the vitellaria and almost all the internal organs are situated in the anterior part of the body. The female and male reproductive organs and terminal part of the gut are situated in the posterior part. The attachment apparatus of adults consist of four pairs of clamps and a pair of small central hooks situated on the ventral side of the opisthaptor. Due to the complicated determination of several groups of monogenean parasites, molecular markers based on species-specific variability in the ribosomal DNA region (rDNA) their cytogenetics have been designed and shown to be useful for precise species identification [1-5]. The interspecific nucleic acid variability of Internal Transcribed Spacers of rDNA (ITS) has also been used to distinguish diplozoid parasites [6-11].
From the available data, it has been concluded that morphological and metrical differences in the clamp size, pharynx size, prohaptoral length, opisthohaptoral length, sucker distance, testis, ovary and egg size were the major criteria for species determination. Species deterimination of trematodes is difficult and demands great skill and experience. As the structures of taxonomic importance (central hooks, clamps etc.,) grow gradually and the measurements of sclerotized structures are variable, species determination of trematodes in different developmental stages is not always clear. There are still some unclear descriptions of trematode species that differ only by host species, and some studies that did not employ recommended criteria [12,13]. Molecular biology techniques have been used as objective methods to distinguish between parasite species. The rDNA genes, particularly the 28S gene, have been found generally useful in molecular taxonomy and phylogeny of parasites [14-16]. However, there are no published molecular studies of trematode genomes from the Kashmir valley. The present study reports the results of molecular analysis of the Internal Transcribed Spacer (ITS) of ribosomal DNA of 3 Monogenean species namely Diplozoon kashmirensis Kaw, Diplozoon aegyptensis Fischthal et Kuntz, Diplozoon guptai [17] using Polymerase Chain Reaction (PCR), nucleotide sequencing and construction of phylogenetic from different fish hosts of the Kashmir valley.
Parasite material
Parasite specimens of Diplozoon spp. were collected from the Carassius carassius; Cyprinus carpio communis; C. c. specularis; Schizothorax niger; S. esocinus; S. curvifrons and S. plagiostomus of Kashmir and were used for DNA extraction. Samples were immediately fixed in 70% alcohol after collecting from the gills, gill cover, mouth cavity, eyes & fins of host fish. These samples were remained in alcohol until the present study.
DNA isolation
Parasite specimens of three Diplozoon species were collected from fish hosts of Carassius carassius; Cyprinus carpio communis; Schizothorax curvifrons; Schizothorax esocinus; Schizothorax niger and Schizothorax plagiostomus from Wular lake, Anchar lake, Dal lake, Manasbal lake, River Jhelum and River Sindh of Kashmir valley preserved in 100% ethanol for genomic DNA extraction and stored at-200C for good quality of DNA. For DNA extraction ethanol was removed from parasites as per the protocol given by [18] and as such, these specimens were air dried to remove ethanol. The resultant DNA was examined on 1.5% agrose-TAE gels, stained with ethidium bromide (EtBr) and visualized under UV light.
Morphological characterization
The present specimens having rectangular opisthaptor with four pairs of clamps and two individuals in form of a cross belongs to genus Diplozoon Nordmann, 1832. When compared D. kashmirensis with D. aegyptensis, they showed similarity in comparative size of clamps, size of eggs, form of oral suckers but differs in the proportion of body length to its breadth, shape of ovary, shape and position of testis. D. kashmirensis resembles D. guptai in egg size, absences of sticky glands in the anterior part, position of testis with respect to ovary, extent of vitellaria and in the arrangement of intestine in hind portion of the worm but shows strong variations as regards total body length, size ratios, clamp size, the shape & size of testis and also prepharynx size. The variations of the three species can be regarded as intraspecific variations due to geographical isolation and are not sufficient for the creation of new species and thus the present specimens are described as D. kashmirensis Kaw, 1950; D. aegyptensis Fischthal et Kuntz, 1963 and D. guptai [17-24] (Table 1).
| Species Particulars | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Particulars | Total Body Length | Forebody | Hind body | Length ratio between fore & hind body | Clamp size | Testis | Egg size | Anterior suckers | Prepharynx | Pharynx | Host | Site | Locality |
| D. kashmirensis Kaw, 1950 |
2.3-4.32 | 1.4-2.64 x 0.71-1.51 |
0.9 x 1.72 x 0.5-0.69 |
1:0.646 | 0.15 x 0.075 0.166 x 0.076 0.154 x 0.075 0.140 x 0.074 |
0.16 x 0.27-0.29 | 0.27 x 0.29 x 0.07-0.09 |
0.063-0.074 x 0.045-0.063 |
0.065 | 0.065-0.075 | Schizothorax sp. | Gills | Dal Lake, Kashmir |
| D. aegyptensis Fischthal et Kuntz, 1963 |
4.529 (3.62-5.77) | 2.665 x 0.558 (1.879-3.452 x 0.299-0.836) | 1.128 x 0.178 (0.867-1.871 x 0.130-0.245) | ----- | 0.070 x 0.097 0.065-0.079 x 0.092-0.102 |
0.136 x 0.08 (0.103-0.19 x 0.063-0.093) | 0.292 x 0.107 (0.524-0.313 x 0.081-0.132) | 0.038 x 0.07 (0.029-0.046 x 0.065-0.079) | 0.027 (0.020-0.034) |
0.062 x 0.044 (0.051-0.075 x 0.040-0.050) | Labeo forskalii | Gills | Giza Fish Market, Giza Fish Market, Egypt |
| D. aegyptensis Fayaz et Chishti 1993 | 4.2 (3.95-4.25) | ----- | ----- | ----- | 0.114 (0.10-0.124) x 0.045 (0.04-0.048) | 0.155 (0.14-0.17) x 0.105 (0.10-0.11) | 0.25 (0.22-0.28) x 0.082 (0.076-0.088) | 0.048 (0.032-0.064) | 0.054 (0.041-0.068) | 0.064 (0.056-0.072) x 0.047 (0.044-0.05) | Schizothorax niger | Gills | Dal lake, Kashmir |
| Diplozoon guptai Fayaz et Chishti 1999 | 1.873 (1.28-2.55) | 1.66 x 0.604 (0.755-0.144 x 0.44-0.65) | 0.603 x 0.395 (0.46-0.76 x 0.289-0.48) | 1:0.523 (1:0.4-0.639) | 0.102 x 0.045 0.106 x 0.032 0.093 x 0.038 0.081 x 0.035 |
0.813 x 0.0786 (0.076-0.12 x 0.052-0.128) | 0.245-0.07 (0.228-0.268 x 0.06-0.08) | 0.056 x 0.05 (0.04-0.08 x 0.032-0.072) | 0.0356 (0.03-0.042) | 0.057 x 0.025 (0.044-0.069 x 0.02-0.028) | Schizothorax niger; S. esocinus; Labeo sp. and Carasius carassius | Gills | Dal and Anchar Lake, Kashmir |
Table 1: Comparative Morphological characteristics of Diplozoon species (measurements in mm).
PCR amplification
The PCR amplified products of ITS regions of rDNA were successfully obtained using the primers (Table 2). PCR amplification was carried out to amplify ITS region of Diplozoon species (Table 3). The size of the amplified product was found to be 873bp long for D. kashmirensis; 1120bp long in D. aegyptensis and 687bp long in case of D. guptai (Figure 1). In BLAST search of these sequences, they showed similarity with other Diplozoon spp. (Table 3). In bioinformatics analysis, the results tallied with those of the earlier study; hence, the same are not repeated here in. Based on morphological studies, these species were identified as belonging to three Diplozoon species. The present results of the molecular analysis corroborate the species identification of these forms. Therefore, it can be assumed that the present species recovered from the different fish hosts of water bodies of Kashmir valley is D. kashmirensis Kaw, Diplozoon aegyptensis Fischthal et Kuntz, and Diplozoon guptai [24].
| Species | Primer Designed | GenBank Accession Number | Author and Year |
|---|---|---|---|
| Diplozoon kashmirensis Kaw, 1950 | Forward | AF 369758 to AF 369761 | Sicard et al., 2001 |
| Cer5.8S 2249:5/GCTCACGTGACGATGAAGAG3/ | |||
| Diplozoon aegyptensis Fischthal et Kuntz, 1963 | Reverse | ||
| Cer28S 3116 :5/TTCGCTATCGGACTCGTGCC3/ | |||
| Diplozoon guptai Fayaz and Chishti, 1999 |
Table 2: Primers used for Trematodes.
| Monogenea | Initial Denaturation | Denaturation for 30 cycles | Annealing | Extension | Final extension |
|---|---|---|---|---|---|
| Diplozoon kashmirensis; D. aegyptensis and D. guptai | 95°C for 10 minutes | 30 cycles at 95°C for 30 seconds | 55°C for 30 seconds | 72°C for 75 seconds | 72°C for 10 minutes |
Table 3: PCR assay of Monogeneans which were carried out in a thermocycler (Eppendorf Mastercycler Personal) under different conditions.
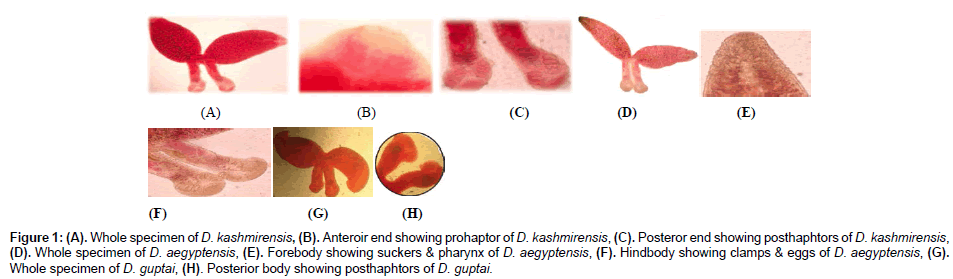
Figure 1: (A). Whole specimen of D. kashmirensis, (B). Anteroir end showing prohaptor of D. kashmirensis, (C). Posteror end showing posthaphtors of D. kashmirensis, (D). Whole specimen of D. aegyptensis, (E). Forebody showing suckers & pharynx of D. aegyptensis, (F). Hindbody showing clamps & eggs of D. aegyptensis, (G). Whole specimen of D. guptai, (H). Posterior body showing posthaphtors of D. guptai.
[Reagents for PCR: Taq DNA polymerase 3 U/μl, dNTP mixture 100 mM, primers 20 pmols, 10 × TaqDNA Polymerase buffer (Genei), PCR water (Sterile milli-Q)].
Sequences deposited in GenBank
GenBank: AF973616; Diplozoon kashmirensis, complete sequence.
GenBank: AF973617; Diplozoon aegyptensis, complete sequence.
GenBank: AF973618; Diplozoon guptai, complete sequence.
The three monogenean species of Trematodes viz., Diplozoon kashmirensis Kaw, 1950; Diplozoon aegyptensis Fischthal et Kuntz, 1963 and Diplozoon guptai [17] which were recovered during the present study are used for molecular study for the first time.
Nucleotide sequences
PCR products were visualized and documented, and the sizes of the sequences were estimated. The sequence obtained from three different Diplozoon species were submitted to GenBank and their accession number acquired were AF973616; AF973617 and AF973618 (Table 4). Sequences were compared with other sequences of monogenean species from GenBank. When the BLAST search was performed, the query sequence showed maximum similarity with 28S rDNA sequence of Diplozoon spp. The nucleotide sequences obtained and shown in (Figures 2-5) are as raw sequences (Table 5).
| S no | Monogenean Species | Host |
GenBank Accession No. | Family | Base pair length | Authors | Country | Year |
|---|---|---|---|---|---|---|---|---|
| 1. | D. kashmirensis Kaw, 1950* | Carassius Carassius, Cyprinus carpio cummunis, Schizothorax niger, S. esocinus, S. curvifrons | AF973616 | Diplozoidea | 873 bp | Present study | India | 2015 |
| 2. | D. aegyptensis Fischthal et Kuntz, 1963* | Carassius Carassius, Schizothorax niger; | AF973617 | Diplozoidea | 1120 bp | Present study | India | 2015 |
| 3. | D. guptai Fayaz and Chishti, 1999* | Schizothorax niger | AF973618 | Diplozoidea | 687 bp | Present study | India | 2015 |
| 4. | D. bliccae (Glaser, 1965) | Blicca bjoerkna | AF369761 | Diplozoidea | 988 bp | Sicard et al. | France | 2001 |
| 5. | D. paradoxum Nordmann, 1832 | Abramis brama | AF369759 and AJ563372 | Diplozoidea | 769 bp | Matejusova | Czech Republic | 2004 |
| 6. | D. homoion Bychowsky & Nagibina, 1959 | Rutilus rutilus, Scardinius erythrophthalmus | AF369760 | Diplozoidea | 996 bp | Sicard et al. | France | 2001 |
Table 4: Monogenean trematode species used for molecular comparison of ITS rDNA sequences along with their hosts, country and GenBank accession numbers for corresponding sequences (*Query sequence).
| Diplozoon kashmirensis | Diplozoon aegyptensis | Diplozoon guptai | |
|---|---|---|---|
| Length | 873 bp | 1120 bp | 687 bp |
| A | 177 | 237 | 123 |
| C | 191 | 224 | 148 |
| G | 226 | 312 | 188 |
| T | 279 | 345 | 228 |
| G+C | 47% | 47% | 48% |
| Total No. of Amino Acids | 280 | 353 | 219 |
| Molecular Weight | 30827 Da | 38825 Da | 24323 Da |
Table 5: Summary of base pairs and amino acids of Diplozoon kashmirensis Kaw, 1950, Diplozoon aegyptensis Kuntz, 1963 and Diplozoon guptai Fayaz et Chishti, 1999.
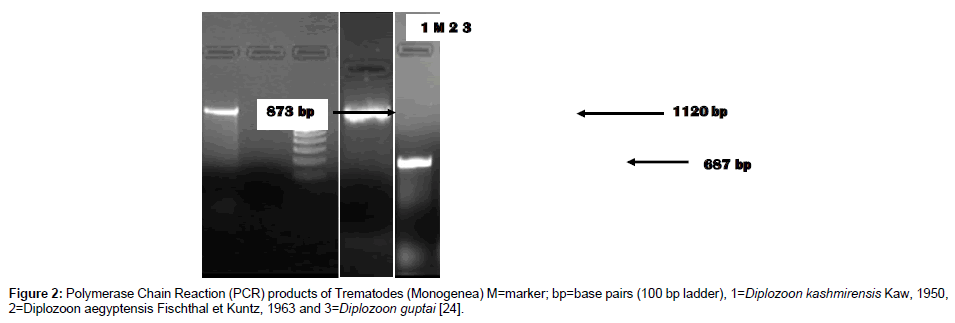
Figure 2: Polymerase Chain Reaction (PCR) products of Trematodes (Monogenea) M=marker; bp=base pairs (100 bp ladder), 1=Diplozoon kashmirensis Kaw, 1950, 2=Diplozoon aegyptensis Fischthal et Kuntz, 1963 and 3=Diplozoon guptai [24].
Figure 3: Raw nucleotide sequences of Diplozoon kashmirensis Kaw, 1950.

Figure 4: Raw nucleotide sequences of Diplozoon aegyptensis Fischthal et Kuntz, 1963.

Figure 5: Raw nucleotide sequences of Diplozoon guptai [24].
Pairwise alignment
Pairwise alignments of Diplozoon species were made by using different softwares such as Gene Runner, DNA Dynamo, Chromas Pro. D. kashmirensis showed maximum similarity with those of D. bliccae where as D. aegyptensis showed maximum similarity with D. paradoxum and in case of D. guptai that showed maximum similarity to D. homoin (Tables 6-8).
| D. kashmirensis D. bliccae |
6 6 | ACTGCCTTGAGCATCGACTTCTTGAACGTAAATTGCGGCATTAGGCTCTGCTGATGCCAC ||| ||||||||||| ||||||||||| ||||||||||||| |||| ||||||||| GCTGACTTGAGCATCGATTTCTTGAACGTGAATTGCGGCATTACCCTCTAATGATGCCAC |
65 65 |
| D. kashmirensis D. bliccae |
66 66 |
GCCTAGCCGAGTGTCGGCATTAAATCTATCACGACGCTTAATTGGTCGTGGCTTAGTTTG |||||||||||| ||||||||||||||| ||||||||||| |||||||||||||| | GCCTAGCCGAGTATCGGCATTAAATCTAGCACGACGCTTATTTGGTCCTGGCTTAGAAAG |
125 125 |
| D. kashmirensis D. bliccae |
126 126 |
TTGTCAGCCGTCGTGTTGTACT---CAACGTGTTGTTCAGTTGTCAAGTCGACGGTATTA |||||||||||||||||||||||||||||||||||||| |||||||||||||||||||| TTGTCAGCCGTCGTGTTGTACTTGGCAACGTGTTGTTCTTTTGTCAAGTCGGCGGTATTA |
185 185 |
| D. kashmirensis D. bliccae |
186 186 |
TTGACGCTTGCCAAATGTAATGGAGAGTTTGTATATGC--AATATCTGCCGGTAGCCTGT ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| TTGACGCTTGCCAAATGTAATGGAGAGTTTGTATATGCGAAATATCTTCCGGTAGCCTGT |
245 245 |
| D. kashmirensis D. bliccae |
246 246 |
TGGTGTTGGCTACGCTGCCCCGTGTATGGTTTATTTGCATTTTTGTGCATACCGATGGGG ||||||||||||||||||||||||||| ||||||||||||||||||||||||||||||| TGGTGTTGGCTACGCTGCCCCGTGTATTTTTTATTTGCATTTTTGTGCATACCGATGGGG |
305 305 |
| D. kashmirensis D. bliccae |
306 306 |
TGGTTAGCTTCTCGTCATCAGTGCGTGTTTGCCGGTGG-GTCGTGGCGTGGGAATTTCAA ||||||||||||||||| |||||||| |||||||||||||||||| |||||||||||| TGGTTAGCTTCTCGTCAGCAGTGCGTCCTTGCCGGTGGTGTCGTGGAATGGGAATTTCAA |
365 365 |
| D. kashmirensis D. bliccae |
366 366 |
TAAGCATTTCTGAATGGTAATTGTGAAATTGTCAT---ATGTGCTGTTCTCTTGAGCCTT ||||||||||||||| ||||||||||||||||||||||||||||||||||||||||| TAAGCATTTCTGAATCCTAATTGTGAAATTGTCATTTTATGTGCTGTTCTCTTGAGCCGC |
425 425 |
| D. kashmirensis D. bliccae |
426 426 |
TTGGCCCACGGGTTGTGCGGTGACCAGTGTTGCTTTGAATGCGAGCGCATGCATGCCAGG |||||||| |||||||| ||||||||| |||||||||||||| |||||||||||||| ACGGCCCACTTATTGTGCGATGACCAGTGACGCTTTGAATGCGAGTGCATGCATGCCAGG |
485 485 |
| D. kashmirensis D. bliccae |
486 486 |
TCGCAGCCTATTTGTGATCGCGAC-GTGCTTTGCTTGTGTTCTGCGTTTAATTTTTGTCA ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| TCTCAGCCTATTTGTGATCGCGACAGTGCTTTGCTTGTGTTCTGCGTTTAATTTTTGTCA |
545 545 |
| D. kashmirensis D. bliccae |
546 546 | CTGTTTCTTGCGAATGAGCGAGTCTGGCCCGAGACGAGATTATGTGCCCATGTCGTGCTG ||||||| |||||||||||||||||||||||||||||| ||||||||||||||||||| CTGTTTCCCGCGAATGAGCGAGTCTGGCCCGAGACGAGAGCATGTGCCCATGTCGTGCTG |
605 605 |
| D. kashmirensis D. bliccae |
606 606 |
TGCAGACATTACTACTCCATTGGTCGCTAAGTGCATATCGGTGTC--CCGTATTCTACTG ||||||||||||||||||||| |||||||||| ||||||||||||||||||| ||||| TGCAGACATTACTACTCCATTCTTCGCTAAGTGTGTATCGGTGTCACCCGTATTTTACTG |
665 665 |
| D. kashmirensis D. bliccae |
666 666 |
TACTGCTGTGGTGTGTGCACCTGACCTCGGATTAGGCGTGATTACCCGCTGAACTTAAGC |||| ||||||||| ||||||||||| |||||||||||||| ||||||||| ||||||| TACTTCTGTGGTGTATGCACCTGACCAAGGATTAGGCGTGATCACCCGCTGAGCTTAAGC |
725 725 |
| D. kashmirensis D. bliccae |
726 726 |
ATATCAATAAGCGGAGGAAAAGAAACTAACCAGGATTCCCTT-GTAACGGCGAGTGAACA |||||||| |||||||||||||||||||||| ||||||||||||||| |||||||||| ATATCAATGGGCGGAGGAAAAGAAACTAACCACTATTCCCTTAGTAACGTCGAGTGAACA |
785 785 |
| D. kashmirensis D. bliccae |
786 786 |
GGGATTAGCCCAGCACCGAAGCTGCGGTC--TTGGCCGTTCGGCAATGTGGTGTTTAGGT ||||||| |||||||||||||||||||||||||||||||||||| ||||||||||| CCGATTAGCAAAGCACCGAAGCTGCGGTCTTTTGGCCGTTCGGCAATCCGGTGTTTAGGT |
845 845 |
| D. kashmirensis D. bliccae |
846 846 |
TGGCATACTCAGGCGATGTACTGTGTAG | |||||||||||||||||||||| TATCATACTCAGGCGATGTACTGTGCCC |
873 873 |
Table 6: Pairwise alignments of the 28S rDNA ITS consequences of Diplozzon kashmirensis and Diplozoon bliccae, numbering refers to ITS sequences.
| D. aegyptensis D. paradoxum |
1 4 | TGCAAACTGCCTTGAGCATCGACTTCTTGAACGTAAATTGCGGCATTAGGCTCTG-CTGA ||||||||||||||||| |||||||| ||||||||||| ||||||||||||||| |||| TGCAAACTGCCTTGAGCCTCGACTTCCCGAACGTAAATTACGGCATTAGGCTCTGCCTGA |
59 63 |
| D. aegyptensis D. paradoxum |
60 64 | TGCCACGCCTAGCCGAGTGTCGGCATTAAATCTATCACGACGCTTAATTGGTCGTGGCTT |||| |||||||||||||||||||||||||||||||||| |||||||||||||| TGCCCGACCTAGCCGAGTGTCGGCATTAAATCTATCACGACATAATATTGGTCGTGGCTT |
119 123 |
| D. aegyptensis D. paradoxum |
120 124 | AGTTTGTTGTCAGCCGTCGTGTTGTACTTGGCAACGTGTTGTTCAGTTGTCAAGTCGACG |||||||| |||||||||||||||||| |||||||||||||||||||||||| |||| AGTTTGTTAAAAGCCGTCGTGTTGTACTTAACAACGTGTTGTTCAGTTGTCAAGTAGACG |
179 183 |
| D. aegyptensis D. paradoxum |
180 184 | GTATTATTGACGCTTGCCAAATGTAATGGAGAGTTTGTATATGCGAAATATCTGCCGGTA ||||||||||||||||||||||||||||||||||| | |||||||||||| || |||| GTATTATTGACGCTTGCCAAATGTAATGGAGAGTTAG-NDATGCGAAATATCCGCTGGTA |
239 242 |
| D. aegyptensis D. paradoxum |
240 243 | GCCTGTTGGTGTTGGCTACGCTGCCCCGTGTATGGTTTATTTGCATTTTTGTGCATACCG |||||||||||||||| |||||| ||||||||||||||| |||||||||||||||||||| GCCTGTTGGTGTTGGCAACGCTGTCCCGTGTATGGTTTACTTGCATTTTTGTGCATACCG |
299 302 |
| D. aegyptensis D. paradoxum |
300 303 | AT-GGGGTGGTTAGCTTCTCGTCATCAGTGCGTGTTTGCCGGTGGTGT----C-GTGGCG || |||| |||||||||| ||||||||| ||||||||||||||||||| | |||||| ATGGGGGCGGTTAGCTTCGCGTCATCAGAGCGTGTTTGCCGGTGGTGTATTGCAGTGGCG |
353 362 |
| D. aegyptensis D. paradoxum |
354 363 | TGGGAATTTCAATAAGCATTTCTGAATGGTAATTGTGAAATTGTCATTTTATGTGCTGTT ||||||||||||| ||||||| |||||||||||||| |||||| |||||||||||||||| TGGGAATTTCAATGAGCATTTGTGAATGGTAATTGTTAAATTGCCATTTTATGTGCTGTT |
413 422 |
| D. aegyptensis D. paradoxum |
414 423 | CTCTTGAGCCTTTTGGCCCACGGGTTGTGCGGTGACCAGTGTTGCTTTGAATGCGAGCGC ||||||||||||||||| ||||||||||||||||||||||||||||||||||| |||| CTCTTGAGCCTTTTGGCTTTCGGGTTGTGCGGTGACCAGTGTTGCTTTGAATGCGTGCGC |
473 482 |
| D. aegyptensis D. paradoxum |
474 483 | ATGCATGCCAGGTCGCAGCCTATTTGTGATCGCGACAGTGCTTTGCTTGTGTTCTGCGTT |||||||||||||||||||||| ||||||||||||||||||||||||||||||||||||| ATGCATGCCAGGTCGCAGCCTA-TTGTGATCGCGACAGTGCTTTGCTTGTGTTCTGCGTT |
533 541 |
| D. aegyptensis D. paradoxum |
534 542 | TAATTTTTGTCACTGTTTCTTGCGAATGAGCGAGTCTGGCCCGAGACGAGATTATGTGCC ||||||||||||||| |||||| ||| |||||| || ||||||| |||||| |||||| TAATTTTTGTCACTGCCGCTTGCGTATGTGCGAGTGTGACCCGAGATGAGATTGTGTGCC |
593 601 |
| D. aegyptensis D. paradoxum |
594 602 | CATGTCGTGCTGTGCAGACATTACTACTCCATTGGTCGCTAAGTGCATATCGGTGTCACC |||||| |||||||| ||||||||||||||| |||||||||||||||| ||||||||| | CATGTCATGCTGTGCTGACATTACTACTCCACTGGTCGCTAAGTGCATGTCGGTGTCATC |
653 661 |
| D. aegyptensis D. paradoxum |
654 662 | CGTATTCTACTGTACTGCTGTG--GTGTGTGCACCTGACCTCGGATTAGGCGTGATTACC ||||||||||||||||||||| |||||||||||||||||||||||||||||||||||| AGTATTCTACTGTACTGCTGTGTTGTGTGTGCACCTGACCTCGGATTAGGCGTGATTACC |
711 721 |
| D. aegyptensis D. paradoxum |
712 722 | CGCTGAACTTAAGCATATCAATAAGCGGAGGAAAAGAAACTAACCAGGATTCCCTTAGTA |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| CGCTGAACTTAAGCATATCAATAAGCGGAGGAAAAGAAACTAACCAGGATTCCCTTAGTA |
771 781 |
| D. aegyptensis D. paradoxum |
772 782 | ACGGCGAGTGAACAGGGATTAGCCCAGCACCGAAGCTGCGGTCTTTTGGCCGTTCGGCAA |||||||| |||||||||||||||||||||||||||||||||||||||||||||||| ACGGCGAG----CAGGGATTAGCCCAGCACCGAAGCTGCGGTCTTTTGGCCGTTCGGCAA |
831 841 |
| D. aegyptensis D. paradoxum |
832 842 | TGTGGTGTTTAGGTTGGCATACTCAGGCGATGTACTGTGCTAAGTCCATTCATGAATATG |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| TGTGGTGTTTAGGTTGGCATACTCAGGCGATGTACTGTGCTAAGTCCATTCATGAATATG |
891 901 |
| D. aegyptensis D. paradoxum |
892 902 | GCTAGCTATCTGGCCCAGAGAGGGTGAAAGGCCCGTGAGCATAGTGCGTCGTTCTGTCTT ||||||||||||||||||||||||||||||||||||||||||||| ||| |||||||||| GCTAGCTATCTGGCCCAGAGAGGGTGAAAGGCCCGTGAGCATAGTACGTTGTTCTGTCTT |
951 961 |
| D. aegyptensis D. paradoxum |
952 962 | AGTCAACCGTTGAGTCGGGTTGTTTAGGAATGCAGCC || |||||||||||||||||||||||| |||||||| AGCCAACCGTTGAGTCGGGTTGTTTAGTAATGCAGCA |
988 998 |
Table 7: Pairwise alignment of the 28S rDNA ITS consequences of Diplozoon aegyptensis and Diplozoon paradoxum, numbering refers to ITS sequences.
| D. guptai D. homoion |
9 6 | ACTGCCTTGAGCATCGACTTCT--AACGTAAATCGCGGTATTAGGCTCTGCCTGATGCCA |||| |||||||||||| ||||||||||| ||| |||| |||| |||| |||||||| ACTGACTTGAGCATCGATTTCTTGAACGTGAATTGCGGCATTACCCTCT-AATGATGCCA |
68 64 |
| D. guptai D. homoion |
69 65 |
CGCCTAGCCGAGTGTTGGCATTATATCTATCACGACGCTTAATTGGTCGTGGCTTAGTTT ||||||||||||| | ||||||| ||||| ||||||||||| |||||| |||||||| CGCCTAGCCGAGTATCGGCATTAAATCTAGCACGACGCTTATTTGGTCCTGGCTTAGAAA |
128 124 |
| D. guptai D. homoion |
129 125 |
GTTGTCAGCCGTCGTGTTTTACTTTGCAACGTGTTGCTCAGTTGTAAAGTCGACGGTATT |||||||||||||||||| ||||| ||||||||||| || ||||| |||||| ||||||| GTTGTCAGCCGTCGTGTTGTACTTGGCAACGTGTTGTTCTGTTGTCAAGTCGGCGGTATT |
188 184 |
| D. guptai D. homoion |
189 185 |
ATTGACGCTTGCCAAATGTAATGGAGAGTGTGTATATGCGAAATTTCTGCCGG-AGCCTG ||||||||||||||||||||||||||||| |||||||||||||| ||| ||||||||||| ATTGACGCTTGCCAAATGTAATGGAGAGTTTGTATATGCGAAATATCTTCCGGTAGCCTG |
248 244 |
| D. guptai D. homoion |
249 245 |
TTGGCGTTGGCGACGCTGCCCCGTGTATGGTTTACTTGCATTTTTGTGCATACCGATTGG |||| |||||| |||||||||||||||| |||| ||||||||||||||||||||| ||| TTGGTGTTGGCTACGCTGCCCCGTGTATTTTTTATTTGCATTTTTGTGCATACCGA-TGG |
308 303 |
| D. guptai D. homoion |
309 304 |
GGCGGTTAGCTTGTCGTCATCAGTGCGTGTTTGCCGGTGGTGATTTGTGGTGGCGTGGGA || ||||||||| |||||| |||||||| |||||||||| ||| |||| ||||| GGTGGTTAGCTTCTCGTCAGCAGTGCGTCCTTGCCGGTGG-----TGTCGTGGAATGGGA |
368 358 |
| D. guptai D. homoion |
369 359 |
ATTTCAATAAGCATTACTGAATGGTAATTAATAAATTGCCATTATATATGCTGTTCTCTT ||||||||||||||| |||||| ||||| |||||| |||| ||| |||||||||||| ATTTCAATAAGCATTTCTGAATCCTAATTGTGAAATTGTCATTTTATGTGCTGTTCTCTT |
428 418 |
| D. guptai D. homoion |
429 419 |
GAGCCTTTTGGCCCACGGGTTGTGCGGTGACCAGTGTTGCTTTGAATGCGTGCGCATGCA ||||| |||||||| |||||||| ||||||||| |||||||||||| | ||||||| GAGCCGCATGGCCCACTTGTTGTGCGATGACCAGTGACGCTTTGAATGCGAGTGCATGCA |
488 478 |
| D. guptai D. homoion |
489 479 |
TGCCAGGTCGCAGCCTA-TTGTGATCGCGACAGTGCTTTGCTTGTGTTCTGCGTTTATTT ||||||||| ||||||| ||||||||||||||||||||||||||||||||||||||| || TGCCAGGTCTCAGCCTATTTGTGATCGCGACAGTGCTTTGCTTGTGTTCTGCGT--AATT |
547 538 |
| D. guptai D. homoion |
548 539 |
GTTGTCACTGCTACTTGCATATGTGCGAGTGTGTACCCGGAATGAGATTTTGTGCCCATG ||||||||| | | || ||| |||||| || |||| | |||| |||||||||| TTTGTCACTGTTTCCCGCGAATGAGCGAGTCTGG-CCCGAGACGAGAGCATGTGCCCATG |
607 597 |
| D. guptai D. homoion |
608 598 |
TCATGCTGTGCTGACATTACTTCTCCACTGGTCGATAAGTGCATGTCGGTGTCACCAGTA || |||||||| ||||||||| ||||| | ||| |||||| | ||||||||||| ||| TCGTGCTGTGCAGACATTACTACTCCATTCTTCGCTAAGTGTGTATCGG-GTCACCCGTA |
667 657 |
| D. guptai D. homoion |
668 658 | CTTTGCTGTA-TT--GTG-T ||| ||||| || ||| | TTTTACTGTACTTCTGTGGT |
687 677 |
Table 8: Pairwise alignments of the 28s rDNA ITS consequences of Diplozzon guptai and Diplozoon homoion, numbering refers to ITS sequences.
Construction of phylogenetic tree
Phylogenetic trees were obtained by comparing the 28S rDNA sequences of the query parasite and other available sequences for related monogenean parasites. The E value was found to be zero up to the 100th sequence of BLAST search and the query coverage 95% and above. The species of D. kashmirensis and D. aegyptensis appeared to be the most closely related species, with well-supported clade by Neighbour joining and MP trees (Figures 6-8).
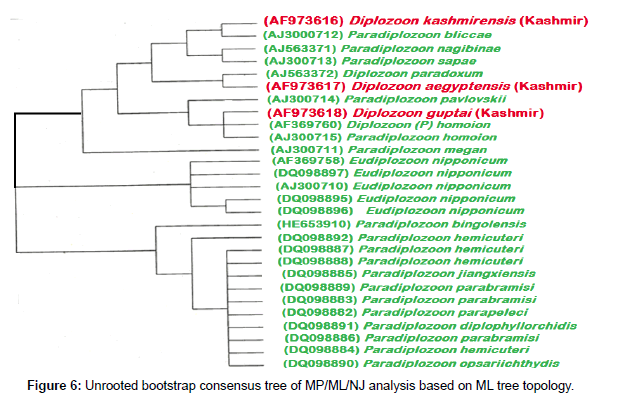
Figure 6: Unrooted bootstrap consensus tree of MP/ML/NJ analysis based on ML tree topology.
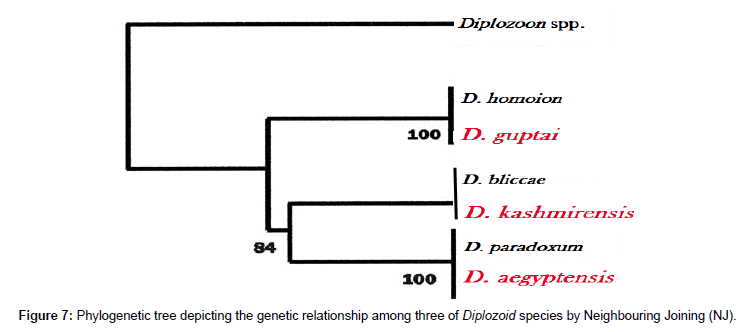
Figure 7: Phylogenetic tree depicting the genetic relationship among three of Diplozoid species by Neighbouring Joining (NJ).
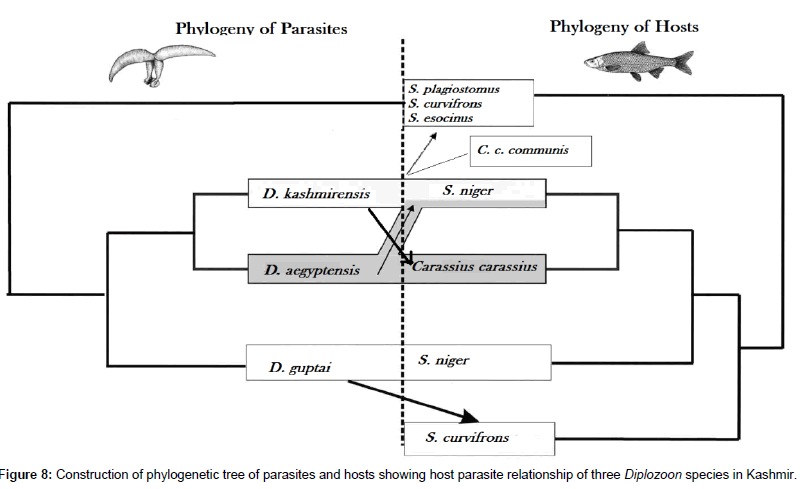
Figure 8: Construction of phylogenetic tree of parasites and hosts showing host parasite relationship of three Diplozoon species in Kashmir.
Above Table shows that Diplozoon kashmirensis having GenBank accession number AF973616 mostly resembles with Diplozoon bliccae with an accession number AF369761.1. Out of 867 base pairs of Diplozoon kashmirensis, 807bp match with that of Diplozoon bliccae i.e., 93.08% similarity with 15 gaps (1.73%).
From the Table 6 it is clear that Diplozoon aegyptensis having GenBank accession number AF973617 shows 94.13% similarity with Diplozoon paradoxum with an accession number AF369759.1. Out of 988 base pairs of Diplozoon aegyptensis, 930 bp match with that of Diplozoon paradoxum with 11 gaps (1.11%).
The present observation shows that Diplozoon guptai having GenBank accession number AF973618 shows 86.16% similarity with that of Diplozoon homoion having GeneBank accession mnumber AF369760.1 (Table 7). 585 bp of Diplozoon guptai matches with Diplozoon homoion with 15 gaps, out of total 679 base pairs.
The rDNA second Internal Transcribed Spacer (ITS2) was amplified using primers Cer5.8S2249 and Cer28S3116 [7] for 3 species of diplozoids. Analysis of the ITS2 region following sequencing clearly allowed us discrimination at the species level and produced the same results as species identification made by using morphological structures. During the present study it was observed that the alignment of nucleotide sequences with those of other Diplozoon species of D. bliccae; D. paradoxum and D. homoion [2,6,7], clearly revealed the boundaries of the 5.8S and 28S rDNA genes, as the sequences in these species closely resembles to those of D. kashmirensis, D. aegyptensis and D. guptai. As noted in comparison of ITS2 sequences of Monogenean species, the first part of the ITS2 is also highly conserved, with only 6 variable sites in the first 65 nucleotides of the diplozoid sequences.
Species discrimination of diplozoids based on the shape of clamp sclerites and the length of the central hook can be difficult because of similarities in the shape of certain sclerites and overlapping ranges of central hook measurements. The PCR product of 3 species of diplozoids: D. kashmirensis; D. aegyptensis and D. guptai were clearly discriminated on the basis of nucleotide sequences which were different in their length of base pairs. The length of the PCR product could be useful to distinguish diplozoids from the genus Eu Diplozoon and Para Diplozoon from other diplozoids [2,6]. Length differences in the ITS2 have also been recorded in the genus Gyrodactylus [2,6] but are not generally as large as those found in the ITS1 region of Lamellodiscus and Gyrodactylus [2,6,19,20]. During the present study there are length difference of PCR products of three Diplozoon species i.e., D. kashmirensis contains 873bp; D. aegyptensis contains 1120bp and D. guptai contains 687bp of 28S rDNA genes, so on the basis of length of base pairs the three diplozoid species can be discriminated. ITS region have been found to be useful species markers for monogenean parasites [1,2-6] so, this method was performed to distinguish the diplozoid species. During the present study, the intraspecific variations within diplozoid species were studied and differences were detected in the ITS regions, but [2,6] studied that ITS region lacks intraspecific variation in groups of Monogenea which is due to the same species recovered from different hosts.
diplozoids are generally considered parasites of Cyprinid species but the host specificity differs and relates to geographical origin. In Eurasia, diplozoid occurrence is restricted to host fishes from the Cyprinidae and Perciformes families [2,6,8,10,21,22]. However, in Africa they also parasitize members of the Characidae [20,22]. All diplozoid species described in the present study are also host specific. ML, MP and NJ trees showed that D. kashmirensis; D. aegyptensis and D. guptai are closely related species, and this mirrors the close relationship of their hosts, thus all of these species are found in cyprinids from the same genus Schizothorax. These species have been described morphologically based on clamp shape, total body length, sucker, and pharynx length [17,24]. The present observations on molecular characterization demonstrate sufficient genetic variations between parasites from different hosts to confirm the validity of these species and that they appear to be host specific, as are many monogenean parasites. It may be speculated that the similarity of these species is a result of a relatively recent divergence of one from the other following a host-switching event. An important observation during the present study has been noticed that Schizothorax niger is infected by all the three species of Diplozoidae: D. kashmirensis; D. aegyptensis and D. guptai, but on all six fishes collected, simultaneous parasitism by all the parasite species was never observed. Two types of factors can be involved in the constitution of such a host-parasite system.
(a) Competition hypothesis: the installation of a first Diplozoon species prevents any other species from settling on the same gill. (b) Since natural hybridization has been reported between the two fishes, the introgression of genes from Carassius carassius into the genome of S. niger allows a host capture of the latter by D. aegyptensis and D. guptai but excludes the infestation by its natural parasite D. kashmirensis.
The present study has confirmed the existence of 3 species of diplozoids from 6 species of cyprinid fishes from the water bodies of Kashmir valley. All the species were clearly distinguished by differences in nucleic acid sequences within the second ribosomal DNA Internal Transcribed Spacer region (ITS2). Analysis of additional specimens from different cyprinid hosts by molecular methods may be helpful to clarify the systematics of this fascinating family Diplozoidae.
The authors extend their thanks to the authorities of the Department of Zoology and Biotechnology, University of Kashmir for providing laboratory facilities. TAS is also highly thankful to Prof. Fayaz Ahmad for giving valuable suggestions while compiling this paper.