International Journal of Physical Medicine & Rehabilitation
Open Access
ISSN: 2329-9096
ISSN: 2329-9096
Research Article - (2022)
Purpose: Architectural properties of the muscles are the prime predictors of functional attributes and force-generating capacity of the muscles. This data is vital for musculoskeletal modelling and selecting the appropriate muscle-tendon units for tendon transfers. Cadaveric data for architectural properties is the gold standard and primary input for musculoskeletal modelling. There is a paucity of these datasets, especially in the leg muscles.
Methods: Sixty muscles of the anterior & lateral compartments from twelve formalin-fixed lower limbs were studied for gross architecture including the peculiar fibre arrangements and architectural properties of muscles. Muscle weight, Muscle length, Fibre length, Pennation angle & Sarcomere length were measured. Normalised fibre length, Fibre length to muscle length ratio (FL/ML Ratio), and the Physiological Cross-Sectional Area (PCSA) were calculated from the obtained data.
Results: Muscles displayed a combination of architectural strategies and were partly fusiform and partly pennate. The tibialis anterior and peroneus longus were the heaviest muscles in their respective compartments and showed more extensive origin from the nearby deep facial sheets.
Long fibre length and less pennation angle were seen in muscles of the extensor compartment. Muscle power was highest in the tibialis anterior and peroneus longus and least in the Extensor hallucis longus.
Conclusion: Arching of the foot and eversion are peculiar to humans and recent in evolution. Due to the functional demand of maintaining the medial longitudinal arch and eversion, the Tibialis Anterior and Peroneus Longus have more muscle weight, and longer PCSA and hence are more powerful.
Extensor compartment muscles were architecturally more suited for excursions because of the long fibre length and less pennation angle. This study contributes baseline normative data for musculoskeletal modelling platforms and simulation tools- an emerging area in biomechanics and tendon transfers.
Medial longitudinal arch; Musculoskeletal modelling; Pennation angle; Sarcomere length; Tendon transfer
Structure-function relationships in skeletal muscles have been examined quite extensively microscopically. However, less attention has been paid to the macroscopic arrangement of muscle fibres known as muscle architecture. Muscle architecture can be defined as, “the arrangement of muscle fibres within a muscle relative to its axis of force generation [1]. Architectural properties of muscles are considered good indicators of force generation and potential for the excursion [2]. Architectural differences are thus the best predictors of force generation [1].
Gans emphasised that the arrangement of fibres in a muscle reflects its function. Parallel fibered muscles are primitive and simplest, providing maximum excursion. The pennate arrangement of fibres allows more fibres to be packed in a muscle, so also increases the cross-sectional area and potential force production [3].
Principles of tendon transfers have been refined over time. It is now understood that properties of muscles like expendability, adequate strength and excursion of the transferred unit are of prime importance [1]. When choosing an adequate muscle-tendon unit to be transferred, the treating physician has to make sure that the unit is expendable, meaning there must be another muscle- tendon unit that can perform the donor unit’s original function. The muscle-tendon unit being transferred should have adequate strength and excursion to be effective [4].
In the last two decades, attempts have been made to study the architectural properties of muscles which are frequently used in tendon or muscle transfers. However, most of the available studies have focussed on the muscle architecture of forearm muscles [5-9], and relatively few studies have focussed on leg muscles [10-12]. Computational modelling and in silico studies is an emerging field of biomechanics. Cadaveric and experimental data is the primary input for such modelling.
Human muscle architecture is also investigated using ultrasound and MRI [13-15]. However, due to limitations of current generation imaging facilities, the gold standard for muscle architecture data remains the dissection of a cadaver [12]. Therefore, the present study intends to describe the gross architecture including the peculiar fibre arrangements and architectural properties of muscles in the anterior and lateral compartments of the leg.
The following parameters were measured in architectural analysis: a) Muscle weight, b) Muscle length, c) Fibre length, d) Pennation angle and e) Sarcomere length. After removing the surrounding fascia and fat tissues, the muscles were harvested intact (from the most proximal origin to the most distal tendon attachment) and were stored in 1x phosphate-buffered saline for 24 to 48 hours before architectural measurements. Muscle specimens were removed from the buffer and gently blotted dry. 1x PBS was formed by the following constituents are 800 ml of distilled water, 8 g of NaCl, 0.2 g of KCl, 1.44 g of Na2HPO4, 0.24 g of KH2PO4, pH adjusted to 7.4 with HCl and distilled water added to a total volume of 1 litre [16].
Muscle Weight (MW)
It was recorded using a Digital Weighing machine with measurements of up to 2 decimals. Muscle was weighed, but the weight was not corrected for formaldehyde fixation. The external tendons, connective tissue and fat were removed before weighing.
Muscle Length (ML)
It was measured as the distance from the origin of the most proximal muscle fibers to the insertion of the most distal muscle fibers by thread and centimeter scale.
Pennation Angle (PA)
It was measured with a goniometer, for more precision, a protractor was kept over it. PA was obtained from the proximal, middle, and distal muscle portions and averaged to yield one value per muscle.
Depending on the morphology of the muscle PA was calculated from different pennate parts of the muscles. It was measured from the unipennate and bipennate parts of the muscle. Circumpennate portions were not included as calculation of pennation was not possible with the methodology used. The mean of PA of different parts of muscles was considered as the final value.
Muscle Fibre Length (MFL)
The muscles were placed in 20% nitric acid to partially digest the connective tissue surrounding the muscles and allow separation of approximately 0.1 mm thick bundles. Average digestion time was 48 hours. Thin fibre bundles were separated, kept straight without stretching and length was measured with a digital vernier calliper. The muscle was divided into three equal parts. Three readings were taken from the centre of each part. The average of the three readings was taken as the final value.
Sarcomere Length (SL)
After acid digestion, individual fibers were removed with forceps and with the aid of microscope. Sections of fiber approximately 5 mm in length were mounted on a micrometry slide using isotonic saline. The muscle was divided into three equal parts. Three sections were taken from the center of each part. Digital images of the fiber sections were taken from a light microscope at 100x magnification. Two readings were recorded from one section, so a total of six readings were recorded from one muscle.
Fiber length was normalised to compensate for natural fiber length variation simply because muscles were fixed at different joint angles. For normalization of fiber length, the sarcomere length within a specimen was measured and a standard sarcomere length (2.7 µm) was selected. Then all raw fibre lengths were normalized using the following equation;
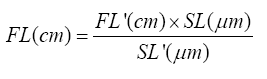
FL: Normalized Fibre Length.
FL’: Raw Fibre Length
SL: Standard Sarcomere Length (2.7 µm)
SL’: experimentally, Measured Sarcomere Length (9)
Fibre length to muscle length ratio (FL/ML Ratio) was calculated by dividing fibre length by muscle length.
The Physiological Cross-Sectional Area (PCSA) was calculated using the following equation:
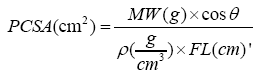
Where ϴ is surface pennation angle and ρ is muscle density (1.056 gcm−3).
Fibre length/ Muscle weight ratio and PCSA/Muscle weight ratio were also calculated. Findings of all study parameters of architectural properties were subjected to descriptive statistics by SPSS version 21.
Gross architecture
The gross architecture in the muscles of the anterior and lateral compartments of the leg varied within and across the compartments. The usual pattern as observed was that the muscle had a uniform fusiform pattern in their proximal fourth or proximal third. Here the muscle fibre ran almost parallel to the long axis of the muscle. In the distal two-thirds or three-fourths, the muscles assumed a pennate pattern (unipennate/bipennate/circumpennate) arranging themselves around an aponeurosis which was initially in the form of broad aponeurosis and when traced distally thickened to form a tendon. The gross architecture of individual muscles as observed in the present study was as follows.
Muscles of lateral compartment: The upper third to an upper fourth of each muscle was fusiform and the remaining portion was variably pennate (Figure 1).

Figure 1: Muscles of the anterior and lateral compartments as visualised from their superficial surface.
Peroneus longus: The lower three-fourths were unipennate where the fibres arising from bony attachment as well as fascial covering of the peroneus longus converge towards the intramuscular aponeurosis. The bulk of muscle fibres gradually reduced distally.
Peroneus brevis: In the middle one-third, the fibres arising from bony/fascial attachment of muscle assume a bipennate form and are unipennate in the lower one-third.
Muscles of anterior compartment
Tibialis anterior: The muscle was triangular in cross-section with the base directed superficially. The intramuscular aponeurosis was leaf-like, which is expanded superiorly and narrowed down inferiorly to ultimately continue as the tendon of the muscle. In the middle half, fibres converge on to the intramuscular aponeurosis from all aspects making it circumpennate. In the lower one-fourth, the muscle was unipennate.
Extensor Digitorum Longus and Peroneus Tertius (EDL and PT)
The muscle belly of EDL and PT was conjoint in the upper part in all the specimens studied. The distal two-thirds of the muscle are separated into five intramuscular aponeuroses with unipennate fibres getting inserted into them. The medial four continued as the tendon of EDL and the lateral most continued as the tendon of PT.
Architectural analysis of muscles of anterior and lateral compartments
Twelve samples were taken for the measurements of each muscle. Average values for all the architectural parameters of individual muscles are compiled in Table 1. Peroneus Tertius was considered as a part of extensor digitorum longus, as the muscle was inseparable from extensor digitorum longus.
| MUSCLE WEIGHT | MUSCLE LENGTH | PA | SL | FL | NFL | FL/ML RATIO | PCSA | FL/MW RATIO | PCSA/MW | |
|---|---|---|---|---|---|---|---|---|---|---|
| Tibialis Anterior | 76.18 ± 24 | 31.03 ± 3 | 13.75 ± 2 | 2.1 ± 0.4 | 8.25 ± 1.2 | 11.16 ± 3.5 | 0.26 ± 0.04 | 8.44 ± 1.9 | 0.11 ± 0.02 | 0.11 ± 0.02 |
| Extensor Hallucis Longus | 18.73 ± 4 | 26.7 ± 4 | 10.16 ± 2 | 1.94 ± 0.2 | 8.82 ± 1.7 | 12.49 ± 3.1 | 0.33 ± 0.06 | 2.06 ± 0.7 | 0.50 ± 0.17 | 0.10 ± 0.02 |
| Extensor Digitorum Longus and Peroneus Tertius | 38.65 ± 9.8 | 33.31 ± 4.8 | 10.71 ± 2.2 | 3.23 ± 0.9 | 7.66 ± 1.2 | 6.80 ± 1.78 | 0.23 ± 0.03 | 4.78 ± 1.37 | 0.20 ± 0.05 | 0.12 ± 0.02 |
| Peroneus Longus | 50.35 ± 16 | 28.95 ± 3 | 18.87 ± 5 | 3.4 ± 0.2 | 4.08 ± 0.6 | 3.17 ± 0.38 | 0.14 ± 0.03 | 11.29 ± 4 | 0.08 | 0.22 ± 0.03 |
| Peroneus Brevis | 23.96 ± 7.2 | 23.08 ± 2.8 | 18.09 ± 5.9 | 3.87 ± 0.75 | 3.54 ± 0.6 | 2.53 ± 0.62 | 0.15 ± 0.02 | 6.11 ± 1.8 | 0.15 ± 0.05 | 0.25 ± 0.03 |
Note: MW: Muscle Weight; ML: Muscle Length; PA: Pennation Angle; SL: Sarcomere Length; FL: Fibre Length; NFL: Normalized Fibre Length; PCSA: Physiological Cross-Sectional Area.
Table 1: Architectural parameters of muscles of extensor and lateral compartments.
Architectural analysis: Peroneus longus measured twice as heavy compared to peroneus brevis. In the anterior compartment, the TA is twice as heavy as EDL and four times that of EHL. Extensor digitorum longus was the longest muscle closely followed by TA. Pennation angle for anterior compartment muscles was comparatively less than muscles of lateral compartment ranging from 10- 14 degrees. In the lateral compartment, PA for PL and PB is similar around 18 degrees.
Muscles of the anterior compartment have long fiber length owing to more excursions produced by the muscles. The FL of these muscles was almost double as compared to the lateral compartment. Tibialis Anterior has a high PCSA, whereas EHL and EDL have low PCSA among leg muscles. Sarcomere length for TA and EHL was lower, whereas for EDL, PL and PB it was on the higher side.
Architectural properties of the muscles are the best indicators of muscle function. The muscle power and excursion are determined by properties like muscle weight, fibre length, PA and PCSA [1,3,17,18]. The muscles of the anterior and lateral compartments seem to exhibit a combination of architectural strategies to optimise the function ascribed to them. This was evident in all the muscles as they had a combination of fusiform (parallel) and pennate arrangement. The parallel fibres determine the excursion of the muscles whereas the pennation added the number of muscle fibres and thus the resultant force.
Architecture properties showed a peculiar trend as follows
Tibialis Anterior was 2 to 4 times heavier than the other extensor compartment muscles. This indicates more powerful muscle action. The pennation angle of the extensor compartment muscles was less and fibre length & normalised fibre length were more, which are indicative of their predominant role in the excursion [2].
The physiological Cross Sectional Area is the summation of the force exerted by individual muscle fibres and determines the force generation capacity of the muscle-tendon units [3]. Pennation results in an increased number of muscle fibres, but there is a resultant loss of force which is compensated by increased PCSA to achieve a critical balance of enhancing the ultimate action of the muscle. While PCSA determines the force generation capacity and fibre length relative velocity of contraction. Muscle power is the product of force and velocity [18]. Hence, the muscle power in descending order was, TA followed by PL, EDL, and PT and the least powerful was EHA. This underscores their role in maintaining the arch and eversion of the foot. It was observed that the muscles TA, EDL, PL and PB also originated to a considerable extent from the nearest available deep facial sheets. This was most prominently observed in the case of TA and PL.
Architectural properties of the anterior and lateral compartments have not been extensively studied. Amongst the available cadaveric studies, Ward et al. have conducted on 21 limbs, while other studies are restricted to only 2-3 limbs [8,11,17-23]. The muscle weight of the TA and the pennation angle of PT showed high variability both in our study as well as that of Ward et.al. [11]. However, fibre length and PCSA were fewer variables. This may be because these parameters are independent of individual anthropometric data. Our findings conformed to those of Ward et.al. in those parameters [11]. A recent in-vivo study by Charles et al. have studied muscle architectural properties with the help of Diffusion Tensor Imaging (DTI) in young adults and has underscored the importance of architectural properties for muscular skeletal modeling [24]. They, however, have also accepted the limitations of this method in terms of its consistency and for measuring FL, SL and Pennation angle.
Cadaveric data is a gold standard for obtaining the normative values for the architectural properties of the muscles for musculoskeletal modelling. There seems to be a paucity of cadaveric data, as all other studies except for Ward et al. are limited to 2 to 3 cadavers [8,11,17-23]. The study will be an addition to the baseline normative data for musculoskeletal modelling and simulation, which is an emerging field with enormous potential to improve patient care and decrease treatment expenses It can clarify the pathophysiology of neurological and musculoskeletal derangements and is useful for predicting the success of novel and ongoing surgical and rehabilitation modalities [25].
The result of this study will aid plastic and orthopaedic surgeons in choosing the appropriate muscle-tendon unit as per requirement in cases of tendon and muscle transfers. When choosing an adequate muscle-tendon unit to be transferred, the treating physician has to make sure that the unit is expendable, meaning there must be another muscle-tendon unit that can perform the donor unit’s original function. The muscle-tendon unit being transferred should have adequate strength and excursion to be effective [4].
Besides acting as dorsiflexors, the extensor compartment muscles have additional functions eg, TA maintains the medial longitudinal arch & PT part of EDL acts as an evertor. Similarly, amongst the lateral compartment muscles, PL, besides being an evertor, also maintains the medial longitudinal arch. Arching of foot and eversion are peculiarly human traits and are still under evolution. Consequently, the muscles engaged in these actions are expected to be more powerful. The architectural properties of TA, PL, EDL and PT and their relative muscle power conform to the role ascribed to them.
• The architectural properties of TA make it suitable for force as well as excursion.
• Extensor muscles are more suited for the excursion.
• The muscle power in descending order was TA>PL>EDL>PB>EHL.
• Muscles supporting the medial longitudinal arch were more powerful followed by evertors
Citation: Kaur G, Lalwani R, Khan MM, Athavale SA (2022) Muscle Architecture of Leg Muscles: Functional and Clinical Significance. Int J Phys Med Rehabil.S20:003.
Received: 04-Nov-2022, Manuscript No. JPMR-22-19945; Editor assigned: 10-Nov-2022, Pre QC No. JPMR-22-19945 (PQ); Reviewed: 25-Nov-2022, QC No. JPMR-22-19945; Revised: 02-Dec-2022, Manuscript No. JPMR-22-19945 (R); Published: 09-Dec-2022 , DOI: 10.35248/2329-9096.22.S20.003
Copyright: © 2022 Kaur G, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.