Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2016) Volume 5, Issue 1
Keywords: Ellman methods; Non-competitive inhibition; kinetic methodology
Ellman methods; Non-competitive inhibition; kinetic methodology
Acetylcholinesterase enzyme (AChE; EC 3.1.1.7) is a large glycoprotein whose basic chemistry was worked out in the 1960s by Lawler, Leuzinger and Baker [1]. AChE belongs to the family of hydrolases whose active site is characterized by a catalytic coordinated triad of three essential amino acids: histidine, serine, and glutamic acid [2,3]. This enzyme is one of the best studied of all enzymes from the point of view of its mechanism of action, the nature of its active site, its distribution and localization in tissue, and its physiologic functions [1]. It is mainly found at neuromuscular junctions and cholinergic synapses in the central (CNS), where its activity terminates the synaptic transmission [4]. AChE has a very high catalytic activity; each molecule of AChE degrades approximately 25,000 molecules of the neurotransmitter acetylcholine (ACh) per second into choline and acetic acid. At the CNS level the produced choline is transported back into the nerve terminals which reuse it in synthesizing new ACh molecules [5-7].
AChE activity can be inhibited by a variety of chemicals, such as organophosphates (OP), carbamates and nerve agents [8] leading to ACh accumulation, resulting in cholinergic receptors over-stimulation and disturbance of many body functions and finally in respiratory arrest and death [9]. Nevertheless, others AChE inhibitors such as physostigmine, pyridostigmine, procycline, donepezil, galantamine, huperezine [10], tacrine and rivastigmine [11] are clinically used against OP poisoning and neurodegenerative diseases like Alzheimer´s (AD). However, all this inhibitors differ in bioavailability, half-life and effectiveness.
N-acetylcysteine (NAC) is a thiol (SH) containing antioxidant, which has been investigated for many pathologies treatment, such as liver failure, inflammation, nephropathy and brain disorders [12]. It is a naturally occurring compound found in several vegetables, including garlic, onion [13], peppers and asparagus [14]. This compound is rapidly absorbed after oral administration in both wild animals and humans [15]. The greatest plasma concentration is reached 2-3 h after administration [16]. NAC has been widely used as an in vitro and in vivo antioxidant in several experiments [17]. NAC exerts its antioxidant action by facilitating reduced glutathione and scavenging the reactive oxygen species formed during oxidative stress [18]. Many are the reports of beneficial effects of NAC such as renoprotective [19], antiangiogenic [20], anticancer [21], hepatoprotective [22], antifibrotic [23], as chelating agent in metal poisoning treatment [24], and as neuroprotective [25]. However, few are the reports about NAC direct effects on cerebral AChE activity.
It is well known that the most common assay used in AChE determination is based on Ellman’s method, which is a photometric method for determining enzyme activity by following the increase of yellow color produced from thiocholine when it reacts with dithio-bisnitrobenzoate ion [26]. In this context, we here performed a kinetic in vitro study of NAC on mice brain AChE activity, in the attempt to get access to a method that uses the 5,5´-dithio-bis-2-nitrobenzoic acid (DTNB) as chromogen compound functionality and to better understand the potentially direct effect of NAC on this enzyme activity.
Animals
Adult male Swiss albino mice from our own breeding colony (20- 30 g) were housed in cages and kept on a 12 h light/dark cycle, at a room temperature of 22 ± 2°C, with free access to food (Guabi, RS, Brazil) and water. All the experiments here described are in accordance with the standards of the Committee on Care and Use of Experimental Animal Resources, the Federal University of Santa Maria, Brazil (Project process number 088/2014). All efforts were made to minimize the number of animals used and their suffering.
Chemicals
Acetylthiocholine iodide (98%) (ATCh), 5,5´-dithio-bis-2- nitrobenzoic acid (DTNB), N-acetyl-L-cysteine (98%) (NAC), were purchased from Sigma-Aldrich (São Paulo-Brazil). All other reagents used in the experiments were of analytical grade and of the highest purity.
Methods
Enzyme source: Six Swiss albino mice brains were homogenized 1:10 (w/v) in cold ice phosphate potassium buffer (KPB) 0.1 M, pH=7.4 with glass/teflon homogenizer. The homogenates were centrifuged at 3000 × g for 10 min at 4°C to yield the salt soluble fraction (SS). The pellet was re-dissolved in KPB 0.1 M, pH=7.4 containing 1% Triton-× 100 and centrifuged at 15000 × g for 10 min at 4°C to yield the detergent soluble fraction (DS), fraction that was used in further assay. Brain mouse choice as enzyme source was based in the perspective to evaluate NAC effects in vivo in futures studies.
Enzyme activity and protein determination: Enzyme activity measurement was performed according to Ellman et al. [26], with some modifications as described in the following steps.
Step 1: In a 96 well plate was added KPB + ethopropazine 20 μM (dissolved in 12 μM hydrochloric acid) and an aliquot of brain DS homogenate (enzyme).
Step 2: DTNB (prepared in KPB) was added at the mixture and allowed to stabilize (stabilization time) at 25oC (for stabilization time determination see point 2.3.3.4).
Step 3: Proceeded to the ATCh (prepared in KPB) addition to start enzymatic reaction.
The reaction velocity was measured by increasing absorbance as a result of yellow anion formation (5-thio-2-nitrobenzoate, TNB-) at 412 nm with an absorption coefficient of 0.0136 μM-1 cm-1, for 2 min and expressed in μmol.h-1.mg prot-1 (Scheme 1).
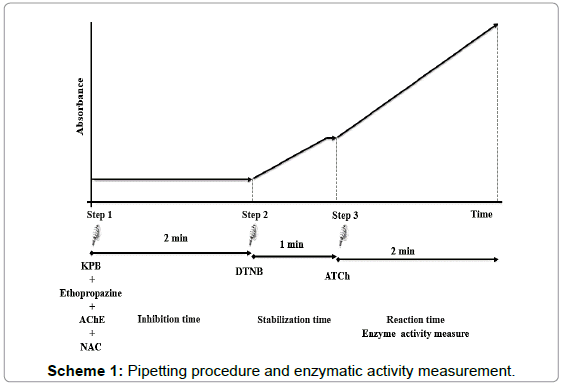
Scheme 1: Pipetting procedure and enzymatic activity measurement.
Protein concentration was measured by the method of Bradford [27], using bovine serum albumin as the standard.
Assay modifications: NAC is a SH containing molecule [22] that can directly interact with the DTNB added to the mixture and so providing false positives results. However, few studies were made with the aim to improve the methodology based in this propriety of NAC, previously to enzyme activity inhibition assay. All experiments were conducted under standard laboratory temperature (25°C).
DTNB concentration and pH determinations: Assays with different concentrations of DTNB were performed. The DTNB final concentrations were of 0.0125, 0.025, 0.05, 0.1, 0.2 and 0.3 mM. After, a pH curve (ranging from 6.8 to 8.0) was performed. From these results, a linearity evaluation was developed, where the 0.3 mM concentration of DTNB and pH=7.4 were chosen for further analysis.
Enzyme activity linearity: For enzyme activity linearity evaluation, enzymatic reaction was performed using different ATCh concentration (0.025, 0.05, 0.1, 0.15 and 0.45 mM).
Stabilization time: Immediately after DTNB addition into the medium containing NAC, an absorbance increase is observed resulting of the formation of the NTB- which ionizes to the NTB2- dianion that has yellow color. The absorbance increase time for each NAC concentration was measured to determine the required time for the NTB2- formation. For this procedure no enzyme was needed, only NAC was added to a tube containing KPB and 0.3 mM DTNB, the absorbance increase was measured for 2 min.
Enzyme inhibition and kinetics parameters: NAC effect on brain AChE activity was conducted using NAC concentrations of 0, 25, 50, 75, 100, 150 and 200 μM. NAC was added to the mixture containing the KPB and the enzyme and allowed to react for 2 min. Procedure was continued with the addition of DTNB as described in step 2 of section 2.3.2. The kinetic parameters Michaelis-Menten constant (Km) and maximum velocity (Vmax), were estimated by assaying the enzyme activity in increasing ATCh concentrations from 0.025 to 0.45 mM.
Time dependence for AChE inhibition and positive control: NAC at the concentration of 169 μM (IC50) was incubated with AChE at different intervals, ranging from 2 to 60 min before the addition of the substrate. After that, the procedure was continued as described in step 2 of section 2.3.2. A positive control (physostigmine 5 μM) was also assayed at the same intervals. The substrate concentration used in this assay was of 0.45 mM.
Statistical analysis: All experimental results are given as the mean (s) ± S.E.M. The statistical analysis was performed using Oneway (ANOVA) followed by the Newman-Keuls when appropriate. p values <0.05 were considered significant. The activities were plotted on Michaelis-Menten and Linewever-Burk plots using nonlinear and linear regression respectively. Dixon and Hill plots were performed to determine the inhibition constant (Ki) and to confirm the enzyme inhibition type induced by NAC [28] using software GraphPad Prism version 5.00 for Windows, GraphPad Software, San Diego, CA, USA (www.graphpad.com).
DTNB concentration and pH
As shown in Figure 1, AChE activity was lower when used DTNB in a concentration of 0.0125 mM (p=0.0002). No statistical difference was observed in DTNB ranging from 0.025 to 0.3 mM. However, the highest enzyme activity was reached with the concentrations of 0.2 and 0.3 mM. Ideal medium pH was of 7.4 (Figure 2). No alteration in pH after NAC addition in the medium was observed. AChE activity was linear (r2=0.9996; p=0.0001), using DTNB 0.3 mM and pH=7.4, along the protocol time (Figure 3).
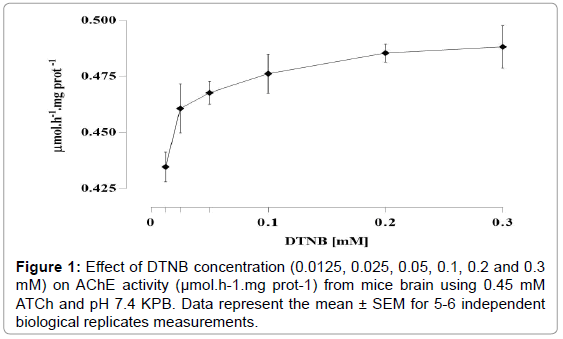
Figure 1: Effect of DTNB concentration (0.0125, 0.025, 0.05, 0.1, 0.2 and 0.3 mM) on AChE activity (μmol.h-1.mg prot-1) from mice brain using 0.45 mM ATCh and pH 7.4 KPB. Data represent the mean ± SEM for 5-6 independent biological replicates measurements.
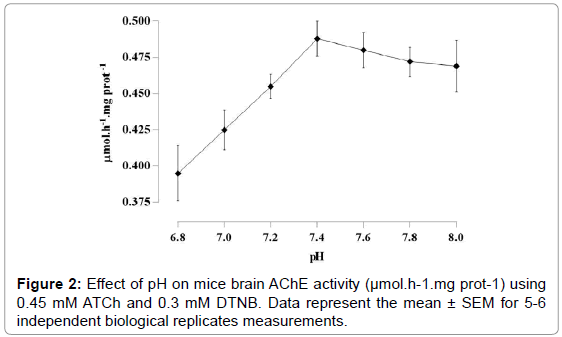
Figure 2: Effect of pH on mice brain AChE activity (μmol.h-1.mg prot-1) using 0.45 mM ATCh and 0.3 mM DTNB. Data represent the mean ± SEM for 5-6 independent biological replicates measurements.
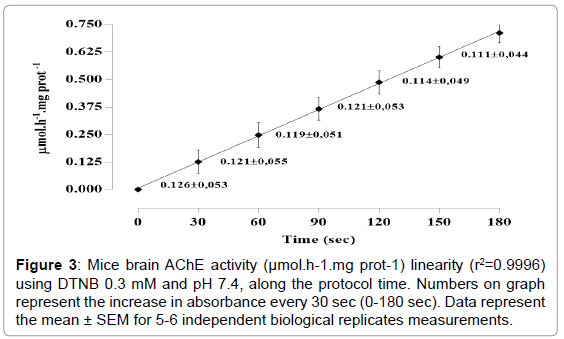
Figure 3: Mice brain AChE activity (μmol.h-1.mg prot-1) linearity (r2=0.9996) using DTNB 0.3 mM and pH 7.4, along the protocol time. Numbers on graph represent the increase in absorbance every 30 sec (0-180 sec). Data represent the mean ± SEM for 5-6 independent biological replicates measurements.
Ache activity linearity
Linear regression demonstrated AChE linear activity for all ATCh concentrations here tested; 0.025 mM (r2=0.9970), 0.05 mM (r2=0.9985), 0.10 mM (r2=0.9975), 0.15 mM (r2=0.9963) and 0.45 mM (r2=0.9978) (Figure 4) along the 2 min of enzyme activity measurement.
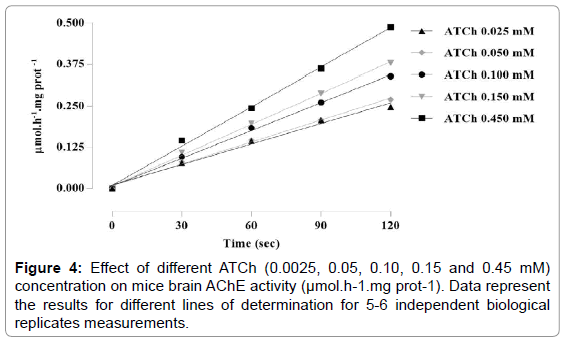
Figure 4: Effect of different ATCh (0.0025, 0.05, 0.10, 0.15 and 0.45 mM) concentration on mice brain AChE activity (μmol.h-1.mg prot-1). Data represent the results for different lines of determination for 5-6 independent biological replicates measurements.
Stabilization time
Direct time reaction between DTNB and NAC was concentration dependent. For the lower NAC concentration (25 μM) the stabilization time was 17 ± 0.0011 sec, while for the highest concentration (200 μM) was 54 ± 0.0049 sec. In view of these results the time of 60 sec was concluded to be enough to avoid any interfere of this interaction (Table 1). This 60 sec were applied to the method assay in step 2 (and so called: stabilization time).
| NAC [μM] | Stabilization time (sec) |
|---|---|
| 25 | 17 ± 0.0011 |
| 50 | 20 ± 0.0021 |
| 75 | 25 ± 0.0026 |
| 100 | 35 ± 0.0049 |
| 150 | 47 ± 0.0051 |
| 200 | 54 ± 0.0059 |
Data are expressed as mean ± S.E.M.
Table 1: DTNB-NAC direct reaction time.
Enzyme inhibition and kinetics parameters
Statistical analysis demonstrated that no significant difference was observed in Km (mM) for all NAC tested concentrations (Figure 5). Nevertheless, significant differences in Vmax (μmol.h-1.mg prot-1) were observed at started from the NAC concentrations of 75 μM (p=0.0369) (Table 2). Absence of Km alteration and decrease of Vmax presented by AChE activity induced by NAC featuring an inhibition of non-competitive type, with a Ki of 147 ± 0.82 μM (Figure 6). The Hill coefficient (h) was < 1.0 which indicates that once one ligand molecule of NAC is bound to the enzyme, its affinity for other ligand molecules decreases (Figure 7).
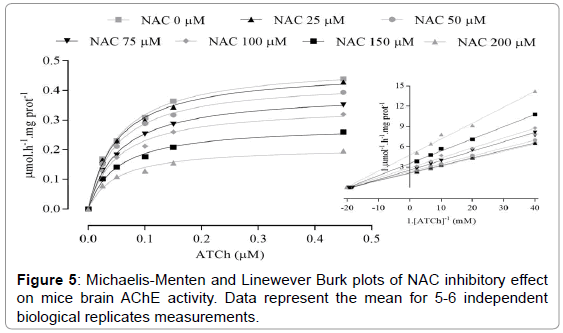
Figure 5: Michaelis-Menten and Linewever Burk plots of NAC inhibitory effect on mice brain AChE activity. Data represent the mean for 5-6 independent biological replicates measurements.
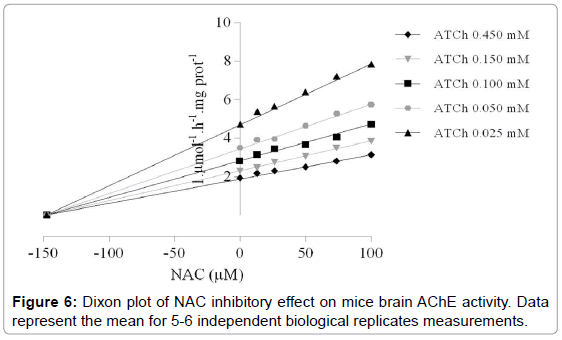
Figure 6: Dixon plot of NAC inhibitory effect on mice brain AChE activity. Data represent the mean for 5-6 independent biological replicates measurements.

Figure 7: Hill plot of NAC inhibitory effect on mice brain AChE activity, showing the slope of the curve providing apparent dissociation constant. Data represent the mean for 5-6 independent biological replicates measurements.
| NAC [μM] | Km [mM] | % decrease | Vmax (μmol.h-1.mg prot-1) | % decrease |
|---|---|---|---|---|
| 0 | 5.33 x 10-3 ± 0.0042 | - | 48.79 x 10-3± 0.0120* | - |
| 25 | 5.19 x 10-3± 0.0041 | 2.63 | 46.97 x 10-3± 0.0116* | 3.73 |
| 50 | 5.02 x 10-3± 0.0030 | 5.81 | 43.21 x 10-3± 0.0080* | 11.43 |
| 75 | 5.46 x 10-3± 0.0032 | 3.19 | 39.19 x 10-3 ± 0.0072* | 19.68 |
| 100 | 5.09 x 10-3 ± 0.0073 | 5.81 | 34.71 x 10-3 ± 0.0155* | 28.86 |
| 150 | 5.09 x 10-3± 0.0068 | 5.81 | 28.21 x 10-3 ± 0.0117* | 42.18 |
| 200 | 4.96 x 10-3± 0.0084 | 6.94 | 21.01 x 10-3 ± 0.0109* | 56.94 |
Data are expressed as mean ± S.E.M. * p<0.05, compared with NAC 0 μM
Table 2: NAC effect on kinetic parameters of AChE activity.
Time dependence and positive control
As shown in figure 8 NAC inhibit AChE activity in a manner independent of time. After the incubation with the IC50 of NAC the remaining enzyme activity was approximately of 50.33%, showing no significant difference between 2 and 60 min. The same did not occur for physostigmine, statistical analysis showed that the incubation time significantly affected the remaining enzyme activity (F: 33.47, p: 0.0001).
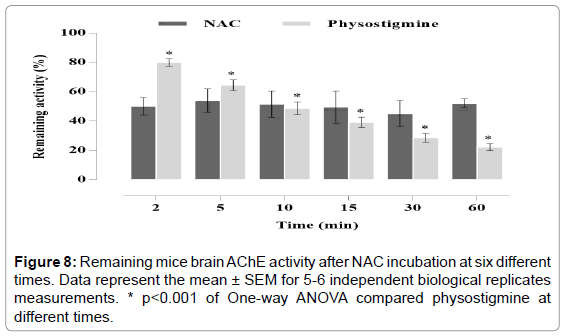
Figure 8: Remaining mice brain AChE activity after NAC incubation at six different times. Data represent the mean ± SEM for 5-6 independent biological replicates measurements. * p<0.001 of One-way ANOVA compared physostigmine at different times.
This study attempted to determine in vitro cerebral AChE activity and the NAC inhibitory effect on this enzyme. The method used was Ellman’s using ATCh as alternative substrate and DTNB as color reagent resulting in production of 5-thio-2-nitrobenzoate (TNB) that has yellow color due to the shift of electrons to the sulfur atom. This method is particularly limited for testing antidotes against AChE inhibitors or for measuring this enzyme activity in samples of such treated individuals, once some antidotes contain reactive oxime group splitting DTNB and providing false positive reaction [29]. In the same way, NAC can also directly react with DTNB, so in this case, it was mandatory to perform some previous experiments in order to assess the method functionality. We here determinate the DTNB concentration and pH required for a linear enzymatic AChE activity in mouse brain and also added a stabilization time to the method in the attempt to avoid probably false positive reaction (absorbance increase). All ATCh concentrations here tested showed linear activity. Finally, the results here achieved show that NAC behaves as a reversible non-competitive AChE inhibitor, by reducing enzyme Vmax in a concentration dependent way without interfering in the Km.
The most common assay used in AChE determination is based on Ellman’s method. Over the last years different chromogens have been tested [30], nevertheless, in this work we choose to use DTNB. The present results demonstrated that DTNB at concentration ranging from 0.025 to 0.3 mM were enough to provide similar AChE activity reading (46.08 × 10-3 and 48.83 × 10-3 μmol.h-1.mg prot-1, respectively) along the protocol time. Thus, we here use the 0.3 mM final concentration to avoid any hypothesis of chromogen consumption by directly interact with NAC. In fact this chromogen has acid pH and can directly interfere with enzyme activity. Here, assays were performed with DTNB solutions in a range from 6.8 to 7.6. As showed by the present results the higher activity was achieved with pH=7.4. Besides, knowing the ideal pH is critical when working with inhibitors. The enzymes inhibition may be greatly affected by pH, in the irreversible no less than the reversible case. The rate reaction of the inhibitor with the enzyme may be affected by pH because the different ionic forms of the enzyme may react at different rates with the inhibitor, or because the inhibitor ionizes itself [28].
For all ATCh concentrations here used the enzyme activity was linear, thus demonstrating that this decrease was not due to lack of substrate. Moreover, substrate inhibition is a kinetic property of AChE [31]. However, the highest substrate final concentration (ATCh 0.45 mM) tested was based on previously studies of our laboratory [32], which demonstrate to be a non-inhibitory substrate concentration. The 1 min waiting time addition (stabilization time) was established to avoid interference of the direct reaction of DTNB with NAC, which occurs immediately after the DTNB addition in the medium, causing an immediate increase in absorbance.
Enzyme kinetics is the study of the chemical reactions catalyzed by enzymes. Through this, we can show the catalytic mechanism of an enzyme, its role in metabolism, how its activity is controlled or how a drug or an agonist might interfere in the enzyme activity. Our results showed that NAC can decrease in vitro AChE activity of mice brains. In 1966, Dixon and Webb proposed that a single enzyme involved in a main metabolic chain inhibition will render the whole chain inoperative and will have a profound or even fatal effect upon the organism. However, some exceptions to this rule apply [28]. To date, the use of AChE inhibitors is one of the few therapies that show consistent positive results in the AD treatment [33]. Here, NAC at 75 μM significantly decrease in vitro AChE activity in approximately 20%, when compared with the normal enzyme activity being the IC50 of 169 μM. As can be seen by the Vmax, the enzyme activity decrease was dependent on the NAC used concentration. However, by varying the substrate concentration the Km remained unchanged, characteristic of a non-competitive inhibition, where the inhibitor molecule binds to an enzyme somewhere other than the active site reducing the enzyme activity or binds equally well to the enzyme whether it has already bound the substrate.
AChE non-competitive inhibitors are clinically used in AD treatment, like Tacrine [34], Donepezil and Huperzine A [35] to name a few. Besides, despite the countless effects of NAC on different neurological disorders like AD by increase the glutathione levels or free-radical scavenging [36], few are the reports of its effect on cerebral AChE activity. Hill coefficients give information on the number of interacting sites. When this coefficient is smaller than one do not indicate anticooperative binding but shows that at least one ternary complex has enzymatic activity [37]. Here, Hill coefficients was smaller than one indicating a possible allosteric binding, where induced conformational changes cause loss of the active conformation. Such a denaturation mechanism is in stark contrast to the desired specificity of drugs.
NAC is structurally similar to cysteine amino acid which can easily form a dipolar ion so called zwitterion at neutral pH [38]. We believe that NAC can also form a zwitterion, a molecule with no overall electrical charge, but that contains separate positive and negative parts (N+ and O-, respectively). The cationic region (N+) present in the NAC zwitterion suffers an electrostatic attraction by the AChE anionic site, inducing a temporally modification in the enzyme esteric site and disabling the neurotransmitter bind (Scheme 2). The fact that NAC inhibitory effect over AChE activity did not vary according to the length of the preincubation period, suggests that the NAC effect is of reversible nature [39,40] similar to physostigmine, which also reversibly to inhibit the AChE, even though that this binds to the active site of the enzyme and not to the anionic one.
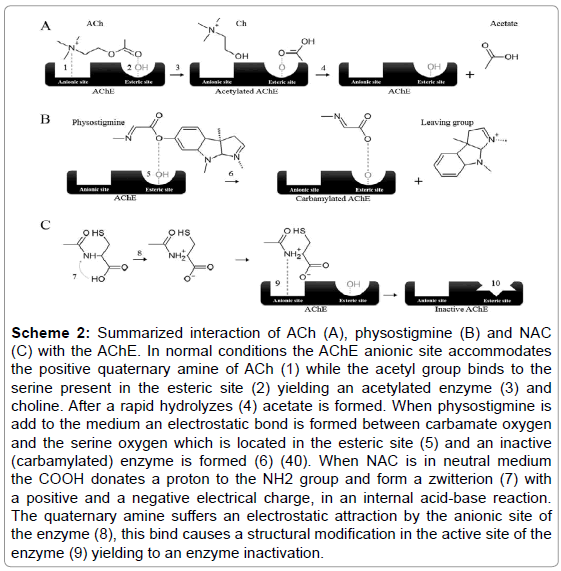
Scheme 2: Summarized interaction of ACh (A), physostigmine (B) and NAC (C) with the AChE. In normal conditions the AChE anionic site accommodates the positive quaternary amine of ACh (1) while the acetyl group binds to the serine present in the esteric site (2) yielding an acetylated enzyme (3) and choline. After a rapid hydrolyzes (4) acetate is formed. When physostigmine is add to the medium an electrostatic bond is formed between carbamate oxygen and the serine oxygen which is located in the esteric site (5) and an inactive (carbamylated) enzyme is formed (6) (40). When NAC is in neutral medium the COOH donates a proton to the NH2 group and form a zwitterion (7) with a positive and a negative electrical charge, in an internal acid-base reaction. The quaternary amine suffers an electrostatic attraction by the anionic site of the enzyme (8), this bind causes a structural modification in the active site of the enzyme (9) yielding to an enzyme inactivation.
Thus, we conclude that the Ellman method is reliable for the analysis of AChE activity when inhibited by compounds which contain SH groups. NAC inhibited non-competitively the enzymatic activity of AChE, been considered as an AChE inhibitor for further in vivo studies.
MEP is recipient of CNPq fellowships (503867/2011-0; 311082/2014-9); MC and TA are recipient of CAPES fellowships, LC is recipient of CNPQ/UFSM fellowships. Additional support was given by CNPq/FAPERGS/DECIT/SCTIE-MS/ PRONEM #11/2029-1.