Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2013) Volume 2, Issue 6
Keywords: Zingiber officinale; Callus; Regeneration, 2,4-D, BAP, SDS-PAGE; RAPD-PCR
BAP: 6- Benzylaminopurine; 2,4-D: 2,4-dichlorophenoxyacetic acid; SDS-PAGE: Sodium Dodecyl Sulfate-Poly Acyrlamid Gel Electrophoreses; RAPD-PCR: Random Amplification Polymorphisms DNA- Polymerase Chain Reaction; MS-medium: Murashige and Skoog medium
Ginger (Zingiberofficinale Rosco) belongs to the family Zingiberaceae is one of the world’s most important spices and produces a pungent, aromatic rhizome that is valuable all over the world not only as a spice but also as herbal medicine for its health promoting properties. Based on several reports [1-3] ginger is constrained severely can summarized as following: (I) ginger normally propagates by its rhizome, with a low proliferation rate, and the reproducing part (the rhizome) is also the economically used part of the ginger plant, which restricts the availability of ginger seeds needed for cultivation, (II) easily infected by soil-born pathogens such as bacterial wilt (Pseudomonas solanacearum), soft rot (Pythium aphanidermatum), and nematodes (Meloidogyne spp.), which cause heavy losses in yield, (III) ginger rhizomes show variations and degeneration under longterm vegetative propagation and (IV) normal breeding of ginger is a real problem due to poor flowering and seed set. Callus induction and plantlets regeneration from different parts of ginger has been reported by several authors [4-6]. Moreover, Abdul Ghafoor [7] who summarized that, among biochemical techniques, SDS-PAGE is most widely used due to its validity and simplicity for describing genetic structure of crop germplasm. Furthermore, Rout et al. [8] demonstrated that RAPD analysis could apply to access the genetic fidelity of ginger plants derived in vitro on an industrial scale as part of crop improvement programs. Further, DNA profile by RAPD techniques has been reported by several workers [9,10]. The present investigation aimed to establishment of reproducible and promising protocols for callus production from primordial leaf explants and plantlets regeneration from ginger plant. Moreover, carrying out molecular characterization of protein and DNA profile (RAPD) of ginger regenerated compare to original materials.
Plant material
Healthy ginger rhizomes were secured from Institute of Medicinal and Aromatic Plants, Dokki, Cairo, Egypt. Rhizomes were carefully cleaned with soap and tap water and kept under darkness condition. After a few days, the induced bud sprouts excised, and then surface sterilized by immersion in 70% ethanol for 2 min, followed by three washes using sterile distilled water. Further, immersed in 50% (v/v) of commercial Clorox solution (5.25% sodium hypochlorite) containing a drop of Twin 20 for 15 min. Subsequently rinsed for several times with sterile distilled water. The obtained sterilized buds were cultured aseptically on MS-medium supplemented with 4.5 mg/l BAP for achievement of shootlets multiplication.
Callus induction and shoots regeneration
Callus induction: The multiplication in vitro plantlets were obtained previously as described by Abbas et al. [11]. Callus cultures were initiated from in vitro primordial leaves of 8-weeks-old plantlets cultures. The basal primordial leaves were cut to small segments (0.5-1.0 cm) nearly uniform in size and placed in glass jars (50 ml) containing MS [12] solid medium, 3% sucrose and 0.7% agar supplemented with various concentrations of 2,4-D and BAP as follow:
| CB | MS-Free growth regulators |
| C1 | MS+2 mg/l 2,4-D |
| C2 | MS+3 mg/l 2,4-D |
| C3 | MS+4 mg/l 2,4-D |
| C4 | MS+2 mg/l 2,4-D+0.2 mg/l BAP |
| C5 | MS+3 mg/l 2,4-D+0.2 mg/l BAP |
| C6 | MS+4 mg/l 2,4-D+0.2 mg/l BAP |
Cultures of all treatments were divided to two groups. The first group incubated and maintained under 16-h photoperiod at intensity of 3000 Lux from cool white light of fluorescent lamps (Phillips). The second group maintained under darkness condition. All cultures incubated at 26 ± 1°C for 8 weeks (two subcultures) and five replicates were used. The obtained observations were recorded as follow:
• Percentage of callus production (CP%) was evaluated according to the following relation: CP%=(explants forming calluses/total number of explants)×100
• Fresh and dry weights (g/jar).
• Dry matter content (%).
Fresh weights of the different calli cultures were dried using oven air drier at 40°C for three days to constant the weight. The percentage of dry matter contents was estimated according the following equation:
Dry matter content (%)=(dry weight×100)/fresh weight
Shoots regeneration: In this part of study, regenerated shootlets obtained using calli cultures previously induced from the first experiment. Whereas, calli cultures were transferred to solidified MS medium supplemented with different concentrations of BAP to study its efficiency on shoots regeneration as follow:
| GB | MS-Free growth regulators |
| G1 | MS+1 mg/l BAP |
| G2 | MS+2 mg/l BAP |
| G3 | MS+3 mg/l BAP |
| G4 | MS+4 mg/l BAP |
The obtained results were recorded as follow:
• Regeneration percentage (%).
• Number of regenerated shoots.
Biochemical markers
SDS PAGE (Sodium Dodecyl Sulfate-Poly Acrylamide Gel Electrophoreses): Biochemical genetic monitoring of SDSpolyacrylamide gel electrophoresis (PAGE) procedure was carried out according to the described method by Laemmli [13] as follow:-
Half gram of in vivo rhizome parts or in vitro micro rhizomes, root, shoot and regenerated shoots was used. The percentages of separating and staking gels used in this experiment were 10% and 3% acrylemide, respectively. The molecular weight of the polypeptide bands were calculated from calibration curve of low molecular weight marker kit of Pharmacia (Uppsala, Sweden).
Isolation of genomic DNA and RAPD (Random Amplification Polymorphisms DNA) analysis: DNA extracted from in vivo rhizomes or in vitro regenerated shoot lets micro rhizomes. The obtained DNA extracts were purified and subjected for random amplified polymorphic DNA (RAPD) analysis using polymerase chain reaction (PCR) technique as described by Doyle and Doyle [14].
RAPD-PCR analysis: PCR amplification was performed in 20ul reaction mix containing 20 ng genomic DNA, 1 unit Taq polymerase (Gibco), and 200 uM each of dATP, dCTP, dGTP, dTTP, 20 p mole random primer (Operon) and appropriate amplification buffer. The mixture was assembled on ice, overlaid with a drop of mineral oil. Amplification was performed according to Williams et al. [15] for 45 cycles, using Biometera Uno thermal cycler, as follows: One cycle at 92°C for 3 min and then 45 cycles at 92 °C for 30 sec, 35°C for 60 sec and 72°C for 2 min (for denaturation, annealing and extension, respectively). Reaction finally incubated at 72°C for 10 min and further 10 min at 62°C.
Electrophoresis: The amplification products were analyzed by electrophoresis according to Sambrook et al. [16] in 0.8% agarose in TAE buffer (for each litre of 50X TAE Stock solution: 242 g Tris Base, 57.1 ml Glacial Acetic Acid and 100 mL 0.5 M EDTA), stained with 0.2 μg/ ml ethidium bromide and documented with gel documentation system to reveal band polymorphism. Nucleic acids bands were photographed and detected under short wave of UV light. 5 primers were tested, three of them (AM-6, AM-7 and AM-8) produced amplification products (Table 1) and their sequences as following:
| Primer | Sequence 5'--------- 3' |
|---|---|
| AM-6 | TCT GCG GCT GTA GTT CCA GT |
| AM-7 | CTT CGG CAG CAT CTC TTC AT |
| AM-8 | CAG TGT GGA AGC CGA TTA TG |
Table 1: Primers used and sequencing.
Resulted data were subjected to statistical analysis, employing F-test for significance at P ≤ 0.05 and computing of L.S.D values to separate means in different statistical groups according to described method by Gomez and Gomez [17].
Callus induction and shoots regeneration
Callus induction: After six weeks of culture, sections of primordial leaves formed yellow/white calli along their edges (Figure 1 a-c). Primordial leaflet sections produced calli under darkness conditions while; the other group incubated under light condition gave no response.
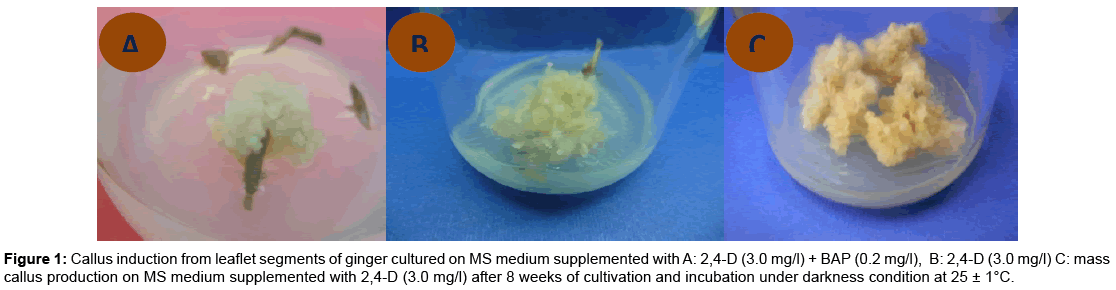
Figure 1: Callus induction from leaflet segments of ginger cultured on MS medium supplemented with A: 2,4-D (3.0 mg/l) + BAP (0.2 mg/l), B: 2,4-D (3.0 mg/l) C: mass callus production on MS medium supplemented with 2,4-D (3.0 mg/l) after 8 weeks of cultivation and incubation under darkness condition at 25 ± 1°C.
Percentage of callus production: Explants were excised from the innermost leaf tissues of two-months-old ginger plantlets then, cultured on MS medium incorporated with various levels of 2,4- D alone or in combinations with BAP and incubated under light or darkness conditions for callus production (Table 2 and Figure 1). Varied significantly, between explants and percentage of callus production across treatments were 27.61%, and all the treatments produced averages of callugenesis did not higher than 50%. MS medium supplemented with 2,4-D at 3 mg/l resulted the highest percentage of callus production 50%. However, the moderate of callus percentage (33.3%) was recorded with 2 or 3 mg/l 2,4-D combined with 0.2 mg/l BAP. Contrary, data showed a little callugenesis at high concentration of 2,4-D alone or in combination with BAP. Accordingly the lowest callus percentage (25 and 10%) was recorded with high concentration of 2,4- D (4 mg/l) alone or in combination with BAP (0.2 mg/l). In addition, there no callus formation was recorded on MS media free growth regulators.
| Code | MS medium supplemented with: | CP% | FW(g/jar) | DW(g/jar) | DM% |
|---|---|---|---|---|---|
| CN | Free growth regulators | 0 | 0.00 | 0.00 | 0.00 |
| C1 | 2 mg/l 2,4-D | 41.66 | 3.35 | 0.12 | 3.92 |
| C2 | 3 mg/l 2,4-D | 50 | 3.87 | 0.14 | 3.66 |
| C3 | 4 mg/l 2,4-D | 25 | 0.96 | 0.06 | 5.91 |
| C4 | 2 mg/l 2,4-D + 0.2 mg/l BAP | 33.3 | 2.47 | 0.11 | 4.39 |
| C5 | 3 mg/l 2,4-D + 0.2 mg/l BAP | 33.3 | 3.77 | 0.12 | 3.07 |
| C6 | 4 mg/l 2,4-D + 0.2 mg/l BAP | 10 | 0.74 | 0.03 | 3.78 |
| LSD0.05 | --- | 1.49 | 0.06 | 1.65 | |
Table 2: Effect of MS-medium supplemented with 2,4-D and BAP at different Concentrations on calli growth parameters induced from primordial leaves of Zingiber officinale after 8 weeks of cultivation under darkness condition at 25 ± 1°C. Where: CP%: Callus Percentage,FW: Callus fresh weight, DW: Callus dry weight DM: Dry matter contents (%), Darkness: 24 h/day
In this respect, Saingproal and Kanchanapoom [18] reported that callus culture of Zingiber officinale was established using young rhizome explants on MS medium supplemented with 23-18.1 μM 2,4-D. Furthermore, Ma and Gang [5] reported that, the presence of 2,4-D at 1.5–5.0 mg/l in the culture medium resulted in callus growth for ginger explants. Moreover, Nayak et al. [19] indicated that, callus of turmeric was initiated from the shoot base segments on MS basal medium supplemented with 1, 3 and 5 mg/l of 2,4-D. In addition, Jamil et al. [6] initiated callus from shoot tip of young ginger plants on MS-media supplemented with NAA (0.1 mg/l), Kin (1-2 mg/l), IAA (0.l mg/l) and BAP (1-2 mg/l). Moreover, Taha et al. [20] established promising protocol for callus production from Helianthus Tuberosus nodal stem and leaf explants by augmentation of MS-medium with 1 mg/l each of NAA and BAP. Further, Wani et al. [21] reported that, leaf and apical bud explants of Tridax procumbens showed maximum response in terms of callus using MS medium with the combination of 2,4-D (0.5 mg/l) and BAP (0.5mg/l).
Calli fresh and dry weights (g/jar): Overall results showed significant differences at the 5% level among different treatments applied. However, data in Table 2 and Figures 1-3 revealed that, MS medium fortified with 2,4-D in concentration of 3 mg/l alone or in combination with 0.2 or 2 mg/l BAP gave the greatest value of fresh and dry mass of calluses (3.87, 3.77, 3.35 and 0.14, 0.12 and 0.12 g/jar, respectively) without significant difference between them. Conversely, the high concentration of 2,4-D (4 mg/l) alone or in combined with 0.2 mg/l BAP gave the lowest value of fresh and dry mass of callus which recorded 0.96, 0.74, 0.03 and 0.06 (g/jar), respectively without significant difference between them. The obtained results were in accordance with those reported by Prakash et al. [22] cleared that, MS medium supplemented with 2,4-D favored for callus initiation from leaf sheath explants of Curcuma amada and the maximum amount of callus was obtained with MS medium supplemented with 9.0 μM 2,4-D. While, Al-Jibouri and Al-Shamarri [23] reported that, plant parts of sugarcane cultured on the MS medium consist of 4 mg/l of 2,4-D was superior in fresh and dry weights of callus. Their results indicated that the fresh and dry weights of callus was decreased as concentration of 2,4-D increased more than 4 mg/l in the medium. In accordance of this result reached by Memon et al. [24] who cleared that, when the amount of 2,4-D was increased (than 3 mg/l )in the cotton cultures medium the rate of callus formation was reduced. Aazami and Hassanpouraghdam [25] indicated that, 1 mg/l 2, 4-D and 2 mg/l BAP gave the highest dry weight of calli in Raphanus sativus cultures.
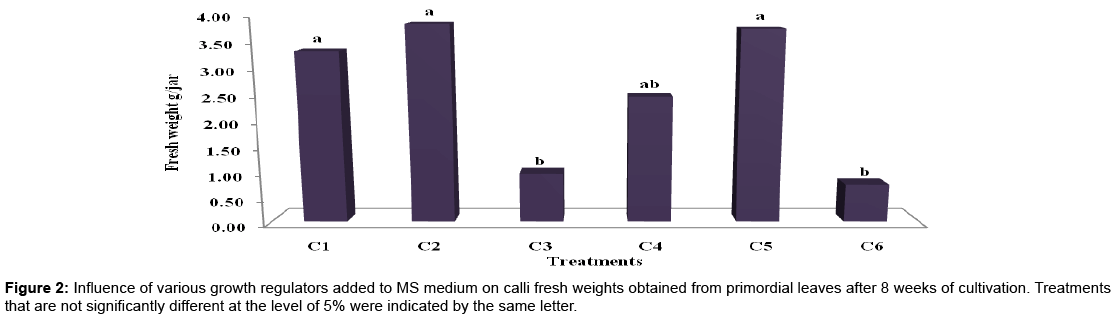
Figure 2: Influence of various growth regulators added to MS medium on calli fresh weights obtained from primordial leaves after 8 weeks of cultivation. Treatments that are not significantly different at the level of 5% were indicated by the same letter.
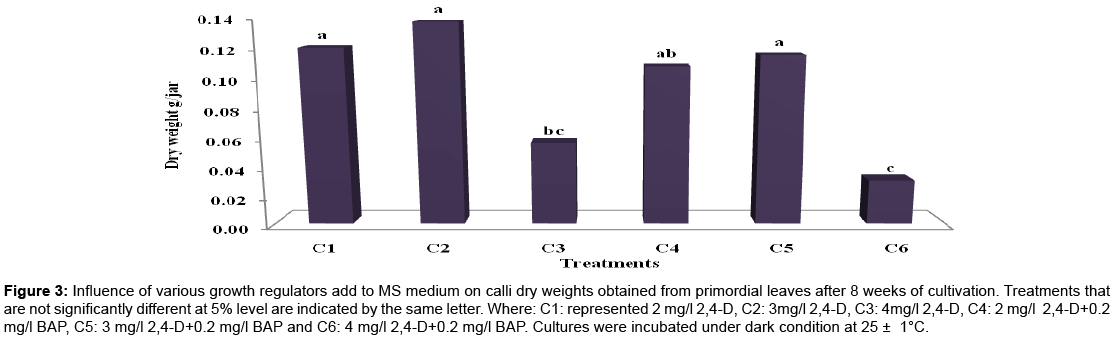
Figure 3: Influence of various growth regulators add to MS medium on calli dry weights obtained from primordial leaves after 8 weeks of cultivation. Treatments that are not significantly different at 5% level are indicated by the same letter. Where: C1: represented 2 mg/l 2,4-D, C2: 3mg/l 2,4-D, C3: 4mg/l 2,4-D, C4: 2 mg/l 2,4-D+0.2 mg/l BAP, C5: 3 mg/l 2,4-D+0.2 mg/l BAP and C6: 4 mg/l 2,4-D+0.2 mg/l BAP. Cultures were incubated under dark condition at 25 ± 1°C.
Dry matter content (%): Dry matter content significantly changed at the 5% level between different treatments, data in Table 2 showed that direct relation between increasing the concentrations of 2,4-D alone and dry matter content (%) in callus. The highest percentages of dry matter contents (5.91 and 4.39%) were recorded with MS medium supplemented with either 4 mg/l 2,4-D or 2 mg/l 2,4-D+0.2 mg/l BAP, respectively. However, the lowest percentages of dry matter contents (3.66 and 3.07 %) were recorded with 3 mg/l 2,4-D alone and in combination with 0.2 mg/l BAP. The obtained results were in accordance with those reported by Lal [26] who cleared that, increasing of the 2,4-D content in the culture media resulted in rapid enhancement of dry matter content of sugarcane calluses and reached to maximum at 5 mg/1 2,4-D level. In addition, Hussein [27] found that, the highest dry matter content of calli derived from seed, leaf, stem and root plants of Thevetla sp were attained on MS medium supplemented with 1 mg/l, 2,4-D+3 mg/l Kin at the 3rd week of cultivation.
Plants regeneration
Regeneration percentage: For achievement of shootlets regeneration, callus derived from leaf tissues was further sub-cultured onto a series of revised MS media supplemented with varying levels of BAP for proliferation of callus and organogenesis as shown in Table 3 and Figure 4. Careful observation revealed that there was a reverse relation between increasing of BAP concentrations and regeneration percentage. The maximum percentage of shootlets initiation (40%) was recorded with MS medium supplemented with 1.0 mg/l BAP while the lowest percentage of shootlets initiation (10%) was recorded with 4.0 mg/l BAP.
| Code | MS medium supplemented with: | Regeneration (%) | Number of regenerated shoots |
|---|---|---|---|
| GB | Free growth regulator | 0 | 0.00 |
| G1 | 1 mg/l BAP | 40 | 1.67 |
| G2 | 2 mg/l BAP | 20 | 0.67 |
| G3 | 3 mg/l BAP | 20 | 0.67 |
| G4 | 4 mg/l BAP | 10 | 0.33 |
| LSD at ( p ≤ 0.05 ) | --- | NS | |
Table 3: Effect of MS medium supplemented with different concentrations of BAP on regeneration percentage and number of regenerated shoots of Zingiber officinaleafter
5 weeks of cultivation under light condition at 25 ± 1°C.
Light: 16h/day
NS: Non- Significant

Figure 4: In vitroshoots regeneration from leaflet segment calli cultures after 5 weeks of cultivation on MS medium supplemented with 1.0 mg/l BAP and incubated under light condition at 25 ± 1°C.
Number of regenerated shoots: Overall results showed no significant differences at the 5% level among different treatments applied. However, datain Table 3 revealed that the highest number of regenerated shootlets (1.67) recorded with MS medium supplemented with 1 mg/l BAP. However, the lowest number of regenerated shootlets (0.4) recorded with 4 mg/l BAP. In vitro plant regeneration is often the most important step for successful implementation of various biotechnological techniques used for plant improvement programmes [28]. The optimistic role of BAP in shootlets regeneration has been reported in ginger and different species by Kackar et al. [29] reported that, efficient plant regeneration of ginger was achieved when embryogenic cultures were transferred to MS medium containing 8.9 μM BAP. In addition, Kim et al. [30] who found that plantlets regeneration of Zingiber offiinale was achieved on MS medium with BAP of 1-2 mg/l from 40 days old calluses. Moreover, Jain et al. [31] reported that, BAP (0.01 mg/l) was found to be essential for obtaining morphogenetic differentiation of the callus, as complete absence of BAP in the callus induction and maintenance medium, resulted in poor regeneration of Cynodon dactylon callus cultures. In addition, Guo and Zhang [4] reported that somatic shoots of ginger embryogenic callus were developed into complete plantlets on solid MS medium supplemented with 3.0 mg/l BAP and 0.1 mg/l NAA. Furthermore, Roopadarshini [32] indicated that, BAP was found to be the best cytokinin compared to Kin for initiation and multiplication of shoots from the compact creamish white callus obtained through vegetative bud explants of turmeric.
Biochemical markers
SDS-PAGE analysis: Total soluble proteins of four randomly selected tissue-culture derived ginger parts were extracted and subjected to protein electrophoresis. To compare the protein banding patterns of different parts produced from in vitro culture with original materials. SDS-PAGE profile of total proteins was extracted from the in vivo rhizomes (mother plant), in vitro microrhizomes, roots, shootlets multiplication and indirect regenerated shootlets of Zingiber officinale.
SDS-PAGE analysis revealed that 12 protein bands with different molecular weights ranged from 56 to 34 kD as shown in Figure 5 and Table 4. Among such protein bands, one band with 41 kD was not appeared in the in vivo rhizomes, while four other protein bands with 45, 41, 39 and 37 kD were disappeared between in vitro produced parts. As general, from Table 5, the regenerated shootlets showed more variable protein bands as compared with in vivo rhizomes. For instance, the directed shootlets revealed the highest protein bands along with all materials used in this study. Moreover, three variable bands with 45, 39 and 37 kD were not detected in indirect regenerated shootlets and two bands with 41 and 37 kD were not detected in the in vitro microrhizomes. This observation was similar to Bertozo and Valls [33] they concluded that the protein profiles of SDS-PAGE was useful for identification and classification of phenotypically specific groups (‘fingerprinting’) and particularly helpful to identify a likely hybrid between the Arachis pintoi accessions.
| Band No. | MW (KDs) | In vitro | In vivo rhizomes | |||
|---|---|---|---|---|---|---|
| Shoot lets multiplication | Indirect regenerated | Root | Micro rhizomes | |||
| 1 | 56 | + | + | + | + | + |
| 2 | 55 | + | + | + | + | + |
| 3 | 54 | + | + | + | + | + |
| 4 | 53 | + | + | + | + | + |
| 5 | 52 | + | + | + | + | + |
| 6 | 48 | + | + | + | + | + |
| 7 | 45 | + | - | + | + | + |
| 8 | 43 | + | + | + | + | + |
| 9 | 41 | + | + | + | - | - |
| 10 | 39 | + | - | + | + | + |
| 11 | 37 | + | - | + | - | + |
| 12 | 34 | + | + | + | + | + |
| Total No. | 12 | 9 | 12 | 10 | 11 | |
Table 4: SDS-PAGE analysis of total protein bands extracted from in vitro and in vivo parts of Zingiber officinale.
| In vivo rhizomes | In vitro | |||||
|---|---|---|---|---|---|---|
| Micro- rhizomes | Root | Indirect regenerated | Shoots multiplication | |||
| In vivo rhizomes | 1.000 | |||||
| In vitro | Micro rhizomes | 0.952 | 1.000 | |||
| Root | 0.957 | 0.909 | 1.000 | |||
| Indirect regenerated | 0.800 | 0.842 | 0.857 | 1.000 | ||
| Shoot lets multiplication | 0.957 | 0.909 | 0.957 | 0.857 | 1.000 | |
Table 5: Similarity index between in vitro and in vivo parts of Zingiber officinale.
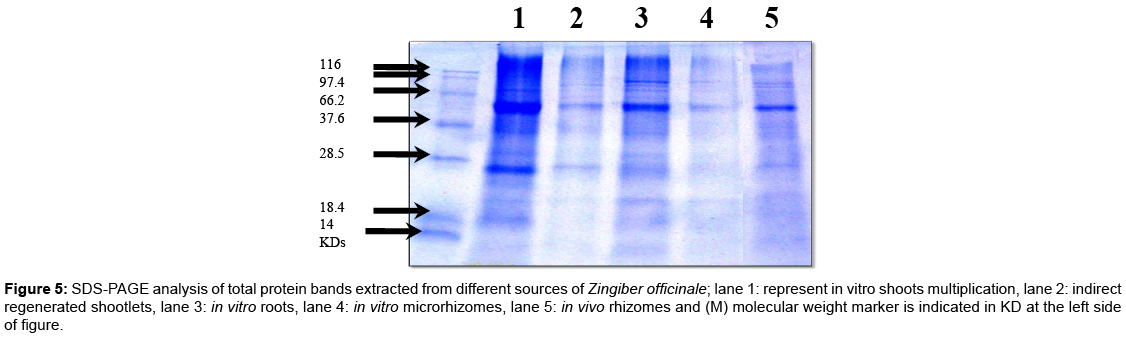
Figure 5: SDS-PAGE analysis of total protein bands extracted from different sources of Zingiber officinale; lane 1: represent in vitro shoots multiplication, lane 2: indirect regenerated shootlets, lane 3: in vitro roots, lane 4: in vitro microrhizomes, lane 5: in vivo rhizomes and (M) molecular weight marker is indicated in KD at the left side of figure.
In addition, Zhao et al. [34] used SDS–PAGE analysis in Polygonatum cyrtonema to determine the properties of newly regenerated rhizomes compared to wild rhizome and the result showed that the content and composition of proteins were similar in the wild and regenerated rhizomes.
Regarding similarity index (Table 5) represented a highly similarity in samples indicating a low variability between different parts produced from in vitro culture with original materials. Some samples got very low similarity indices and some got high similarity indices. Similarity index ranged between 0.800 and 0.957 with average 87.85. The highest value of similarity detected between in vitro and in vivo samples was formed in shoots and roots (0.957). In contrast, the lowest value of similarity was found in indirect regenerated shootlets (0.800). Similar electrophoretic migration rate was observed with four types of tissue cultures compared to original materials. However, high similarity between tested samples indicated that in vitro microrhizomes, root, shoot and indirect regenerated shootlets were similar in protein patterns with those in vivo rhizomes.
RAPD analysis: DNA of in vivo and in vitro different parts was prepared and amplified by PCR using random oligonucleotide primers. Amplification products were separated by agarose-gel electrophoresis to reveal band polymorphism. Out of the 5 random primers screened, only three primers produced clear reproducible bands (sequences presented in Table 6. The 3 primers yielded 104 scorable bands (with an average of 34.67 bands per primer), including seven polymorphic bands. The number of bands from each primer varied from 24 to 44. The primer AM-7 in particular, produced a large number of strongly amplified individual fragments (44), whereas, primers AM-8 produced the lowest number (24) of amplicons. On the other hand, primer AM-7 gave the highest percentage of polymorphism (9.09) while the lowest percentage (4.17) was obtained by using the primer AM-8. The number of amplification products generated by the 3 primers was 104 bands ranged from 24 in AM-8 to 44 in AM-7 (Table 6), with a size range of 130 bp in AM-7 to 652 in AM-6.
| Primer | Sequence 5'--------- 3' | Total Number of bands | Total Number of polymorphic bands | Polymorphism [%] |
|---|---|---|---|---|
| AM-6 | TCT GCG GCT GTA GTT CCA GT | 36 | 2 | 5.56 |
| AM-7 | CTT CGG CAG CAT CTC TTC AT | 44 | 4 | 9.09 |
| AM-8 | CAG TGT GGA AGC CGA TTA TG | 24 | 1 | 4.17 |
Table 6: RAPD-PCR amplification products of DNA extracted from in vivo and in vitro produced Zingiber officinale plants using three random primers.
Using primer AM-6, two polymorphic band, first band with a molecular mass 652 bp in shoots was absent and the other with a molecular mass 234 bp in in vivo rhizomes was absent (Figure 6 and Table 7), with the primer AM-7, two polymorphic bands with molecular mass 169 and 130 bp were absent in in vivo rhizomes, indirect regenerated and shootlets. However, for the primer AM-8 one polymorphic band with a molecular mass 189 bp in shootlets was absent. RAPD analysis of micro propagated plants, micro rhizomes and indirect regeneration shootlets of ginger showed a profile similar to that of the control indicating that no genetic variation had occurred in vitro. Further, RAPD technique could successfully apply to determine the genetic fidelity of ginger plants. RAPD variation has reported in many studies. For example, reports have indicated the occurrence of Rout et al. [8] who demonstrated that RAPD could apply to access the genetic fidelity of ginger plants derived in vitro and the monomorphic RAPD profiles in comparison with control plants. Moreover, Salvi et al. [35] concluded that genetic molecular markers were considered to be reliable in monitoring variability in the DNA sequences of plants. RAPD analysis of micro propagated plants of turmeric showed a profile similar to that of the control indicating that no genetic variation had occurred in vitro. On the other hand, Rady [36] reported that DNA (RAPD) analysis of Gypsophila using 9 primers, the total number of amplification products generated by (PCR) was 142 bands (15.7 bands per primer), of which 7.74 % showed polymorphism. The results indicated that very low variation at the DNA level occurred during in vitro culture. However, Tyagi et al. [37] assessed genetic stability of in vitro-conserved plants by RAPD analysis which showed no evidence of polymorphism between mother plants and the plants of Curcuma longa L.
| Primer | Size of polymorphic bands (bp) | Distribution of polymorphic bands | |||
|---|---|---|---|---|---|
| In vivo rhizomes | shoot letsindirect regenerated | shoot letsmultiplication | In vitro micro rhizomes | ||
| AM-6 | 652 | + | + | - | + |
| 234 | - | + | + | + | |
| AM-7 | 169 | - | + | + | + |
| 130 | - | - | - | + | |
| AM-8 | 189 | + | + | - | + |
Table 7: Distribution and size of polymorphic bands from in vivo and in vitro produced Zingiber officinale plants using 3 primers producing polymorphic bands.
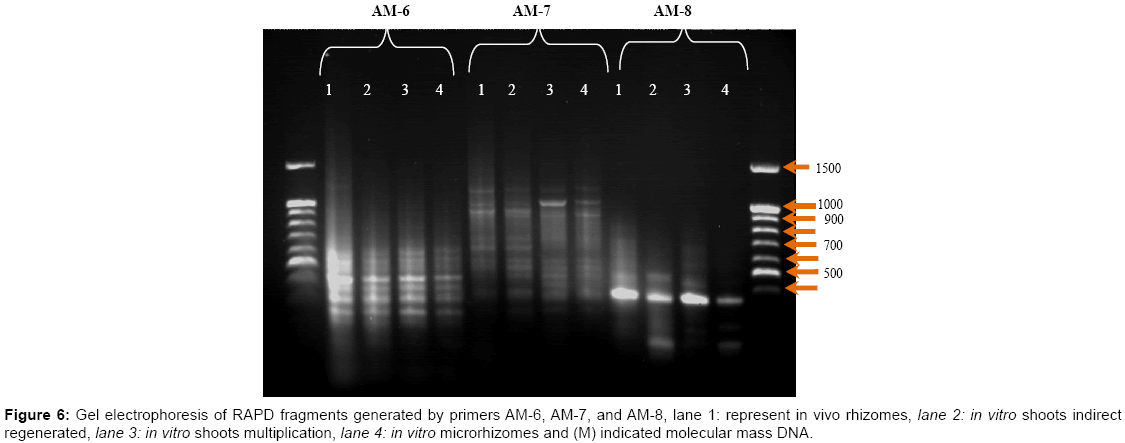
Figure 6: Gel electrophoresis of RAPD fragments generated by primers AM-6, AM-7, and AM-8, lane 1: represent in vivo rhizomes, lane 2: in vitro shoots indirect regenerated, lane 3: in vitro shoots multiplication, lane 4: in vitro microrhizomes and (M) indicated molecular mass DNA.