Journal of Cell Science & Therapy
Open Access
ISSN: 2157-7013
ISSN: 2157-7013
Research Article - (2015) Volume 6, Issue 6
Introduction: The lesser quantity products of reprogrammed stem cells and slower differentiation of stem cells into neurons have limited the advance of cell therapy in clinical applications. Neurotrophic factors BDNF, GDNF, FGF, and IGF are critical factors for further differentiation and proliferation of neuronal cells. However, the influence of the reprogramming genes on the neurotrophic factors is unclear.
Methods: Murine primary embryonic brain cells were transfected with cDNA constructs combining nonviral reprogramming genes and with/without complete length cDNA constructs of these neurotrophic factors. Reprogrammed iPSCs and progressive differentiated neural cells and controls were observed using methods of imaging and quantities.
Results: Our results suggested: 1) During time-courses from the transformation of iPSCs into progressively staged neuron cells, the non-viral reprogramming genes have been significantly accelerated formations of progenitor cells, neuron cells, and neuron network, respectively. 2) The non-viral reprogramming genes directly increased gene expressions of BDNF, GDNF, FGF and IGF at RNA levels. 3) cDNA BDNF plus reprogramming genes showed a robust induction of the immature neuronal marker doublecortin at the protein level. Reprogramming genes and neurotrophic factors fuel stem cells.
Conclusion: This study presents a high-efficiency approach for producing non-viral reprogrammed stem cells and auxiliary differentiated neuronal cells, which would potentially apply in the future clinical applications
Keywords: Nonviral; Reprogramming genes; Neurotrophic factors; Stem cell; Neuron cells
BDNF; Brain-Derived Neurotrophic Factor; GDNF: Glial Cell Line-Derived Neurotrophic Factor; FGF: Fibroblast Growth Factor; IGF: Insulin-like Growth Factor; iPSCs: Induced Pluripotent Stem Cells; Vector 20866, pCAG2LMKOSimO; TH: Tyrosine Hydroxylase
Cell transplantation therapies in clinical applications have been hammered by the use of natural human fetal tissues, even though some patients, such as those with Parkinson’s disease (PD) who received cell transplantation therapy, have shown exciting improvements [1]. Thus, one of the key issues for translational research is to explore the resolutions by replaceable technical approaches such as applying engineered natural cell/tissue to substitute natural human fetal tissues. At this point, the greatest challenge is to optimize the isolation, proliferation and differentiation of specific functional types of cells from alternative sources. Transcription factors Oct4, Sox2, Klf4, and c-Myc have been identified as having the ability to reprogram embryonic stem cells [2] and adult Reprogramming genes and neurotrophic factors fuel stem cells somatic stem cells [3] into induced pluripotent stem cells (iPSCs), which can then be further differentiated into targeted functional types of cells, including neurons [4].
The lack of efficient methods to produce high quantities and increased generation of reprogrammed stem cells hinders the transition to clinical application. High-yields of stem cells, neuronal cells and even dopaminergic neuronal cells will undoubtedly expedite successful clinical applications. At present, this issue has greatly inspired researchers’ interests by exploring a variety of technical approaches. One methodology is to use chemical small molecules for modifying cell culture conditions [5] for accelerating high harvest generation of neuronal cells, such as retinoic acid for neuronal differentiation [6]. The second route is to apply multiple active growth factors and to identify signaling transductions for specific cell transformations and increased rates of products. Conditions, which transformed natural stem cells into neuron cells and further differentiated into dopaminergic cells, tyrosine hydroxylase (TH)-positive cells, have been determined. These active growth factors include interleukin-1beta, glial cell line-derived neurotrophic factor (GDNF), neurturin, transforming growth factorbeta3, dibutyryl-cyclic AMP [7,8], sonic hedgehog [9-11], fibroblast growth factor-8 (FGF8) [11], and brain-derived neurotrophic factor (BDNF) [12-16], and, the signaling transductions involving Bcl-XL [17], Lmx1b/Wnt [18], developmental transcription factors ASCL1, NURR1, and LMX1A [19]. The third approach is to supply spherical neural masses as a physical method for producing the high yield of dopamine neurons [20,21]. The fourth path is to use biomaterial or combining biomaterial with biological cell growth active factors coated cell culture surfaces [22-24]. All of the above are under Reprogramming genes and neurotrophic factors fuel stem cells the ideal natural source conditions, but not applying reprogrammed stem cells. However, the sources of the natural stem cells were restricted by ethics, and thereby applying reprogrammed stem cells become into important sources in the future for clinical cell translation applications. What are the impacts of reprogramming genes themselves on stem cells to produce neuronal cells in the amounts and rapidity? What are interactive effects of reprogramming genes themselves with the neurotrophic factors? They are quite unclear among these relationships.
Another important technical concern is that transplantation of stem cells may pose the potential risk for causing cancers [25], because the viral vectors carrying engineered reprogramming genes may integrate and modify host genomic DNA. Effective and safe of patient-specific stem cells for applications of personalized medicine are still deficient at present.
Therefore, our present report investigated the efficiency of speeding ratio and quantities of products for transforming mice embryonic brain stem cells into neuron cells by engineering non-viral vector with four reprogramming genes: Oct4, Sox2, Klf4, and c-Myc. Lastly, we studied the impact of BDNF, as the most important neurotrophic factor, on quantities of neurons at the protein level.
Plasmid constructions
Reprogramming genes and neurotrophic factors fuel stem cells. In order to re-program mice embryonic brain cells into induced pluripotent stem cells (iPSCs), a reprogramming cassette c-Myc-Klf4- Oct4-Sox2-(MKOS)-IRES-mOrange, named pCAG2LMKOSimO (vector 20866, Addgene), was engineered [26,27] into the primary culture cells. The most significant advantages for this plasmid are a non-viral vector backbone and removable reprogramming four genes c-Myc-Klf4-Oct4-Sox2. These reprogramming genes are transcribed by synthetic CAG enhancer/promoter driving for the universally expressed [28,29]. In addition, this plasmid contains a mOrange for it to be delivered into cells as a gene expression marker.
With the intention of examining impact of BDNF on formation of reprogrammed neuron cells from mice embryonic brain cells, the complete length sequences of 750bp cDNA for BDNF from Mus musculus (GenBank BC034862), 723bp cDNA for GDNF from Mus musculus (GenBank NM_010275), 465bp cDNA for FGF2 from Mus musculus (GenBank NM_008006), and 480bp cDNA for IGF1 from Mus musculus (GenBank NM_010512) were respectively constructed. Briefly, the cDNA BDNF, cDNA GDNF, cDNA FGF2, and cDNA IGF1 were separately constructed by using PCR and cloned into the pZsYellow1-N1 Vector (Clontech 632445) at the ligation sites of XhoI and SalI. These cDNA constructs were transcribing for gene expression by synthetic human cytomegalovirus (CMV) immediate early enhancer/promoter driving. The cDNA constructs were verified for Sanger Sequencing by using Applied Biosystem’s 3730xl DNA Analyzer technology; robust gene expressions by transfecting them into adult mouse primary hippocampal and pituitary cells, adult rat primary hippocampal cells in Reprogramming genes and neurotrophic factors fuel stem cells vitro system; and by transplanting them into the cortical region of mouse brain in vivo system.
Primary tissue cell culture
Primary embryonic brain tissues were isolated 14 days pregnant mice (Figure 1). All actions were permitted by the Carleton University Committee for Animal Care and were directed in adherence to guidelines set out by the Canadian Council for the Use and Care of Animals in Research. A matrix of 6-well plates were set by coating culture containers with Poly-D-Lysine (Millipore A-003-E, final covering concentrations from 1 μg/cm2 of surface area) and laminin (Invitrogen 23017-015, final coating concentrations from 1 μg/cm2 of surface area,). Brain tissues were positioned on a plate with 4 mL of the Hibernate®-E Medium without Ca2+ (BrainBits LLC, HE-Ca). Minor fragments of tissue were consumed using 2 mg/mL of filter-sterilized enzyme papain (Worthington, LS003119) at 37°C for 45 minutes. Continually, tissues were removed and spun down and the supernatant re-suspended in 5 mL of complete Hibernate®-E medium (BrainBits LLC). The tubes were additionally centrifuged for 4 minutes at 200×g, after which supernatants were removed and cell pellets re-suspended in 41.5 mL Neurobasal® medium (Invitrogen, 21103-049). A total of 50 mL of the medium was prepared by accumulating a extra 7.5 mL knockout™ serum replacement (Invitrogen, 10828-028), 0.5 mL L-glutamine (Invitrogen, 25030-081), 0.5 mL MEM Non-Essential Amino Acids Solution (NEAA) (Invitrogen, 11140-050), 50 μl of 10 μg/mL (final 10 ng/mL) bFGF (Fibroblast growth factors, Invitrogen, 13256-029) and 91 μl (final 0.1 mM) β- Reprogramming genes and neurotrophic factors fuel stem cells mercaptoethanol (Invitrogen, 21985-023). Cells were raised at 37°C in a moistened air of 5% CO2 in the ambient environment and were newly nourished every third day with the suitable medium.
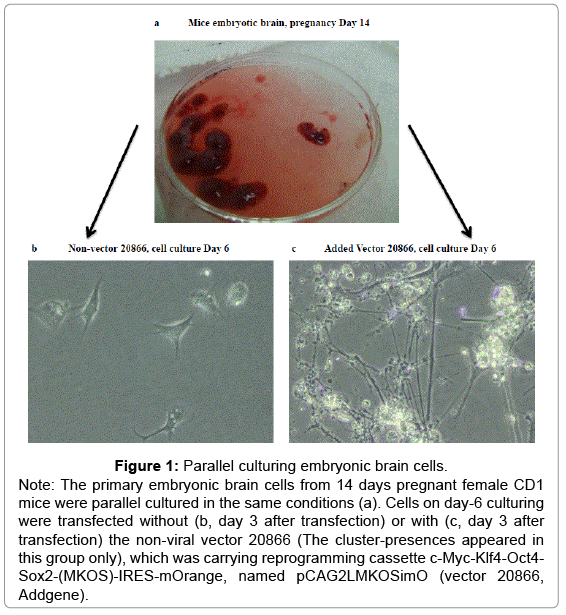
Figure 1: Parallel culturing embryonic brain cells.
Producing induced pluripotent stem cells
Cells (0.5 x 105 cells) were transfected with 4.0 μg DNA vector 20866 in each 6-well plate by using lipofectamine® 2000 Transfection Reagent (Invitrogen, 11668-027), when cells were 85% confluent. Cells with or without vector 20866 (parallel control group) were constantly cultured with fresh complete Neurobasal® medium as described above to induce pluripotency. The cluster-presences started to appear from Days 3-4 after transfection. Further, iPSCs were detected and verified by an expression of stem cell stage-specific antigens SSEA-1 and GF141 on Day 18. The vector 20866 mOrange-positive marker with iPSCs performed gene expression from Days 14-17 and peak period during Day 22-24. The cluster-appearance being confirmed by live staining with GF141 were further transferred to 100 x 60 mm plates, coated with 0.1% gelatin attachment factor solution (Invitrogen, S-006-100), and cultured with fresh a Neural Induction Medium. This was a complete N2B27 Medium (Invitrogen, 11330-057, plus N-2 Supplement 17502- 048, B-27® Supplement 17504-044, knockout™ serum replacement, L-glutamine, NEAA, β-Mercaptoethanol), supplemented with a CHALP cocktail, which has significantly improved the reprogramming efficiency [30]. The CHALP cocktail included: PD0325901 (MEK inhibitor, 0.5 μM, Stemgent 04-0006); CHIR99021 (GSK3β inhibitor, 3 μM, Stemgent 04-0004); A-83-01 (TGF-β/Activin/Nodal receptor inhibitor, Reprogramming genes and neurotrophic factors fuel stem cells 0.5 μM, Stemgent, 04-0014); hLIF (human leukemia inhibitory factor, 10 ng/mL, Millipore LIF1005); HA-100 (ROCk inhibitor, 10 μM, Santa Cruz, sc-203072); and bFGF (100 ng/mL). In order to avoid and reduce toxic effects of CHALP cocktail on cells and to increase vital ability of cells, the low dose 5% embryonic stem cell–qualified fetal bovine serum (Invitrogen, 10439) was added to the complete N2B27 Medium. The medium was altered to Essential 8™ Medium (Prototype), containing DMEM/F-12 (HAM) 1:1, Essential 8™ Supplement (50X) (Invitrogen, A14666SA), N-2 Supplement, B-27® Supplement, knockout™ serum, L-glutamine, NEAA, and β-Mercaptoethanol during Days 15-26. The medium was substituted every 3rd day until Day 26 post-transfection.
Induction and differentiation of neural cells
The neural progenitor cells were induced from the undifferentiated iPSCs by forcedly direction culturing under the Neural Induction Medium from Days 27 to 30. This medium was comprised of 50 mL Neurobasal® medium with bFGF, heparin solution (Sigma H3149, 50 μl of 2-mg/mL), N-2 supplement, glutaMAX™-I supplement, nonessential amino acids solution, knockout™ serum replacement, and β-Mercaptoethanol.
To differentiate to the neural progenitors of the midbrain fate from the neural progenitor cells, cell cultures were nourished with Dopamine Neuronal Progenitor Medium, incorporating 100 ng/mL FGF-8b (Invitrogen, PHG0271) and 200 ng/mL sonic hedgehog (SHH, R&D systems 1314-SH-025), as well as Neurobasal® medium, heparin solution, Reprogramming genes and neurotrophic factors fuel stem cells N-2 supplement without vitamin A (Invitrogen, 12587-010), and NEAA during Days 31–35.
Moreover, from Days 36 to 130 the more mature neurons were further derived from the dopamine neural progenitor cells by culturing in the Dopamine Differentiation Medium. This medium contains: recombinant human BDNF (Invitrogen, PHC7074, 50 μl of 25-μg/mL), recombinant human GDNF (Invitrogen, PHC7045, 50 μl of 20-μg/mL), ascorbic acid (Sigma A4403, 50 μl of 200 mM); dcAMP (Dibutyryl cyclic-AMP) (Sigma D0627, 50 μl of 1-mM) in 50 ml Neurobasal® medium containing heparin, N-2 supplement without vitamin A (Invitrogen, 12587-010); and NEAA. All different mediums above were freshly changed every other day.
Cell culturing for parallel control
In order to have a high quality control group for the neuron cells, the same source of cells from the primary embryonic brain tissues being isolated from 14 days pregnant female CD1 mice were prepared in the identical procedure and time course as the parallel culturing condition, as described before (Figure 1). The standard cell culture medium for the neuron cells were applied, containing total of 50 mL of the medium for each time freshly preparation every 3rd day before changing medium with 41.5 mL Neurobasal® medium, 3 mL embryonic stem cell– qualified fetal bovine serum, 4.5 mL knockout™ serum replacement, 0.5 mL L-glutamine, 0.5 mL MEM Non-Essential Amino Acids solution, 50 μl of 10 μg/mL (final 10 ng/mL) bFGF and 91 μl (final 0.1 mM) β- mercaptoethanol. Cells were set up in the same ambient atmosphere as the reprogrammed Reprogramming genes and neurotrophic factors fuel stem cells cells did.
Immunocytochemistry detection of staged reprogrammed stem cells
In order to determine iPSCs, the stem cell markers alkaline phosphatase live stain (Invitrogen, A14353), and the cell-surface Stage- Specific Embryonic Antigens (SSEA-1), as well as expression of GF141 antigens (Stem Cell Factor Protein; Millipore, GF141) were respectively utilized. The neuronal cells were, correspondingly, identified for immature neuronal cell by using marker anti-DCX antibody (Invitrogen, 48-1200), for mature neuronal cell by applying marker anti-MAP2 antibody (Abcam, ab32454), and for specific dopamine neuronal cell by employing marker TH antibody (ImmunoStar, 22941). Briefly, the primary antibodies at 1:25 ratio were diluted in blocking solution (1X PBS/ 4% normal goat serum/ 0.3% Triton™ X-100), and then incubated for 1 hour at room temperature following 3-time washes with 1 X Rinse Buffer. Furthermore, samples were incubated with secondary antibodies labeled with fluorescein isothiocyanate (FITC) for 1 hour at room temperature. Fluorescence images were envisaged with an Olympus 1x2-UCB series microscope and MediaCybernetics imaging software while living cell culture samples were pictured by applying a Zeiss Axiovert 40 CFL microscope and Lumenera Infinity software.
Impacts of reprogramming genes on BDNF, GDNF, FGF, and IGF
Once 85% confluence of primary embryonic brain tissue isolated from 14 days pregnant female CD1 mice, cells were transfected with 4.0 μg complete cDNA constructs BDNF, Reprogramming genes and neurotrophic factors fuel stem cells GDNF, FGF2, and IGF-1 into the appropriate experimental groups by using lipofectamine® 2000 Transfection Reagent (Invitrogen, 11668-027) in each 6-well plate. After 48 hours with fresh medium, half numbers of cell plates were co-transfected with 4.0 μg plasmid vector 20866 containing four reprogramming genes by applying lipofectamine® 2000 Transfection Reagent; another half numbers of cell plates were without transfection of vector 20866 as control groups. Then, all cells were continued for culturing with fresh Neural Induction Medium for extensive 48 hours, before harvesting by 0.5 ml of cold 4°C PBS after washing cells 3 times, and storing -80°C for late measurements of quantitative real-time PCR.
Quantitative real-time PCR for gene expression at RNA levels
Initially, total RNA was isolated from cells using a GeneJET RNA Purification kit (Thermo scientific, K0732). Thereafter, RNA was quantified using real-time PCR (qRT-PCR), applying the TaqMangene- specific primers and the LightCycler® 480 RNA Master Hydrolysis Probes kit (Roche, 04991885001). The primers were designed using the Integrated DNA Technologies website software program (Table 1). All results of RNA levels were normalized to total levels of a control housekeeping gene GAPDH, as 35 cycle phases were gene expression measured. The experiments repeated three times in the duplicate loading for each sample control.
| Genes | Primers | |
|---|---|---|
| BDNF | forward | GTGACAGTATTAGCGAGTGGG |
| Probe | CAGTTGGCCTTTGGATACCGGGA | |
| Reverse | GGGATTACACTTGGTCTCGTAG | |
| GDNF | forward | TGAAGACCACTCCCTCGG |
| Probe | CTGACCAGTGACTCCAATATGCCTGA | |
| Reverse | GCTTGTTTATCTGGTGACCTTTTC | |
| FGF2 | forward | TCTACTGCAAGAACGGCG |
| Probe | AGCGACCCACACGTCAAACTACAA | |
| Reverse | CTCCCTTGATAGACACAACTCC | |
| IGF1 | forward | TGGATGCTCTTCAGTTCGTG |
| Probe | AGGTGCCCTCCGAATGCTGG | |
| Reverse | TCATCCACAATGCCTGTCTG | |
| GAPDH | forward | CTTTGTCAAGCTCATTTCCTGG |
| Probe | CACCCTGTTGCTGTAGCCGTATTCA | |
| Reverse | TCTTGCTCAGTGTCCTTGC |
Table 1: Primers of TaqMan-specific probes for real time RT-qPCR.
Effect of cDNA BDNF construct on reprogramming genes
Once living cell culture samples were determined for 85% confluence of reprogrammed immature neuron cells from the undifferentiated iPSCs on Day 27 after ready using Reprogramming genes and neurotrophic factors fuel stem cells plasmid 20866, as described before, cells were transfected with/without 4.0 μg complete cDNA BDNF construct into the appropriate experimental groups by using lipofectamine® 2000 Transfection Reagent (Invitrogen, 11668- 027) in each 6-well plate. Cells were continued for culturing with fresh Neural Induction Medium from Days 27 to 30.
Western blotting for doublecortin gene expression at protein level
After washing 3 times by ice-cold PBS, cells were harvested by using ice-cold RIPA Buffer (Sigma, R0278), containing a ready-to-use 1X solution was formulated as follows: 150 mM NaCl, 1.0% IGEPAL® CA- 630, 0.5% sodium deoxycholate, 0.1% SDS, and 50 mM Tris, pH 8.0, in addition to freshly prepared Protease/Phosphatase Inhibitor Cocktail (Cell Signaling, 5872). Samples were frozen in the dry ice right away and put -80°C for detection of the protein by Western blotting. Cells were lysed and sonicated for 2 seconds and protein concentrations were measured by a standard BCA assay (Pierce) with instrument Spectra Max 190 (Molecular device company). All samples were heated in Laemmli buffer and 15 μg of total protein subsequently loaded on 12.5% SDS-PAGE gel. Blots were then probed with a rabbit polyclonal anti-DCX antibody (Invitrogen, 48-1200) against the immature neuronal cell marker, DCX (doublecortin, identifies the target band at ~40 kDa). Secondary antibodies which were HRP conjugated were then smeared and proteins visualized by ECL treatment and subsequent exposure to Kodak film. For measurement of loading controls, all lanes of membranes were further striped with Stripping buffer (25 mM glycine-HCl, pH 2, 1% SDS) for washing 3 times for 10 min with each in 1X PBS-0.1% Tween for washing out primer antibodies DCX and secondary Reprogramming genes and neurotrophic factors fuel stem cells antibodies. And then, the membranes were loaded with the betaactin antibody (Abcam, ab8227) at 1/5000 dilution for detection of each lane’s beta actin as the control loading.
Statistical analyses
These tests were performed using A StatView (SAS Institute, version 6.0) statistical software package. Power analyzes were completed by means of Power and Sample Size Calculations. Two-sided P-values below 0.05 were measured statistically significant.
Reprogramming genes transformed embryonic brain cells into iPSCs
The non-viral and excisional vector 20866 was constructed by Kaji et al (2009) for in vitro reprogramming to transform mouse and human fibroblasts into iPSCs. However, the effect of these reprogramming genes on the more natural states of embryonic brain cells is unclear, which encouraged us to inquire whether and what functional changes would occur after inducing reprogramming genes into nature brain cells themselves. This is a critical issue to be addressed in the application of reprogramming genes in the stem cell replacement.
We successfully established iPSCs from reprogrammed primary embryonic brain cells of 14-day aged pregnancy female CD1 mice after being transfected with the non-viral vector Reprogramming genes and neurotrophic factors fuel stem cells 20866 (Figure 1). Our results suggested that four reprogramming transcription factors Oct4, Sox2, Klf4, and c-Myc were able to transform embryonic brain cells into iPSCs with multiple induce pluripotency (Figure 2). The cluster-look iPSCs were confirmed by expressions of stem cell biomarkers alkaline phosphatase stain (AP, Figure 2b) on day 14, stage-specific antigens SSEA-1 and GF141 on day 18 (Figure 2c and 2d). The vector 20866 mOrange-positive marker with iPSCs performed peak gene expression on day 22 (Figure 2f). These cluster-appearance iPSCs present significantly difference with parallel original cell culturing embryonic brain cells as the control in the morphology (Figure 2a and 2e).

Figure 2: Producing iPSCs by reprogramming genes conversion of embryonic brain cells.
Reprogramming genes accelerated embryonic brain cells into neurons
Upon successfully validating the capability of the non-viral reprogramming system to transform embryonic brain cells into iPSCs, our next stage was to yield the neuronal phenotype cells. In particular, we were interested in increasing products of the neural cells derived from reprogrammed stem cells. Higher proliferative products would present more efficiency to get recovery of brain damages. To generate neuron cells and then dopamine neuron cells, the Neural Induction Medium, Dopamine Neuronal Progenitor Medium and Dopamine Differentiation Medium were applied throughout during Days 27–130 (see Methods section for details). In Figure 3A, transformation to produce the neuron cells from reprogrammed iPSCs via different staging - Progenitor neuron cells (Day 26; 3A-d), Mature neuron cells (Day 31; 3A-e), and Neuron network (Day 86; 3A-f) appeared significantly faster speeding than non-reprogrammed embryonic brain cells (original and parallel cell culturing) to form Progenitor neuron cells (Day 83; 3A-a), Reprogramming genes and neurotrophic factors fuel stem cells Immature neuron cells (Day 86; 3A-b) and Network forms of neuron (Day 127; 3A-c). Furthermore, these reprogrammed neuron cells were verified by the mature neuronal marker MAP2 (3B-a), and tyrosine hydroxylase (TH) as the rate-limiting enzyme in dopamine synthesis (3B-b) on Day 50. There are the accelerating ratios for Progenitor neuron cells for 3.2 times (Mean ± SEM for non-programmed cells 83 ± 2 days vs programmed cells 26 ± 2 days; P value<0.03); Neuron cells for 2.8 times (Mean ± SEM for non-programmed cells 86 ± 3 days vs programmed cells 31 ± 2 days; P value<0.03); and Neuron network for 1.5 times (Mean ± SEM for non-programmed cells 127 ± 4 days vs programmed cells 86 ± 3 days; P value<0.03), respectively (Figure 3a-i).
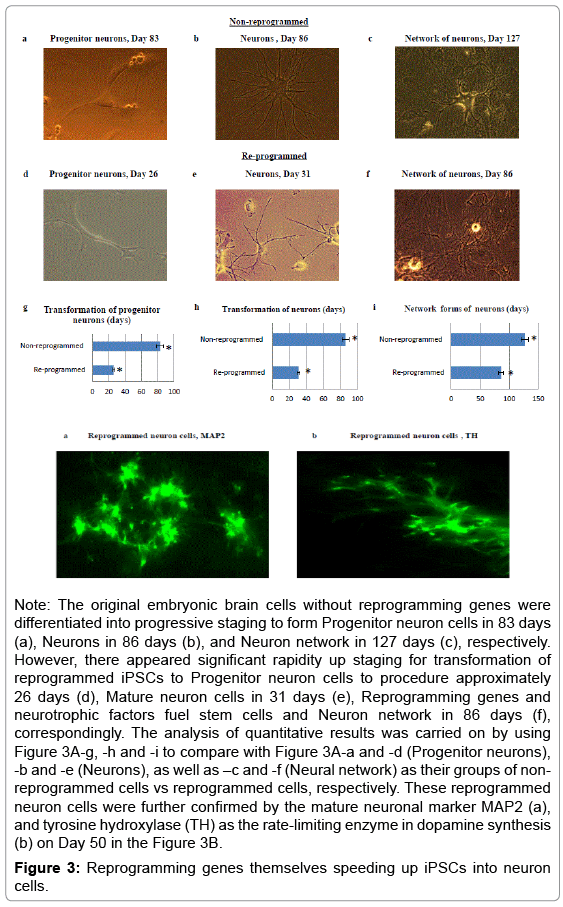
Figure 3: Reprogramming genes themselves speeding up iPSCs into neuron cells.
Reprogramming genes enhanced gene expressions of BDNF, GDNF, FGF and IGF
Since our data showed non-viral reprogramming genes speeding up transformation of mice embryonic brain stem cells into neuron cells, we further wonder whether or not influences of reprogramming genes themselves on neurotrophic factors BDNF, GDNF, FGF2, and IGF-1 at RNA levels. As shown in Figure 4, the mRNA expressions for the neurotrophic factors BDNF, GDNF, FGF2 and IGF1 by TaqMan-specific-probes real-time PCR were increased after 48-hour co-transfected with their respective complete length cDNAs plus the vector 20866 reprogramming genes (a, red lines; duplicates), when compared with their controls vector 20866 reprogramming genes alone (b, green lines; duplicates). The non-viral reprogramming genes themselves directly up-regulated gene expressions of BDNF (30%, Mean ± SEM for 30 ± 2.6, n=3), GDNF (25%, Mean ± SEM for 25 ± 2.3, n=3), FGF (17%, Mean ± SEM for 17 ± 2.1, n=3) and IGF (23%, Mean ± SEM for 23 ± 2.4, n=3), respectively. Figure 4 represented one of three experimental Reprogramming genes and neurotrophic factors fuel stem cells replications for Ct (threshold cycle) plots. All results of RNA levels were normalized to total levels of a control housekeeping gene GAPDH, whose primers are presented in Table 1.
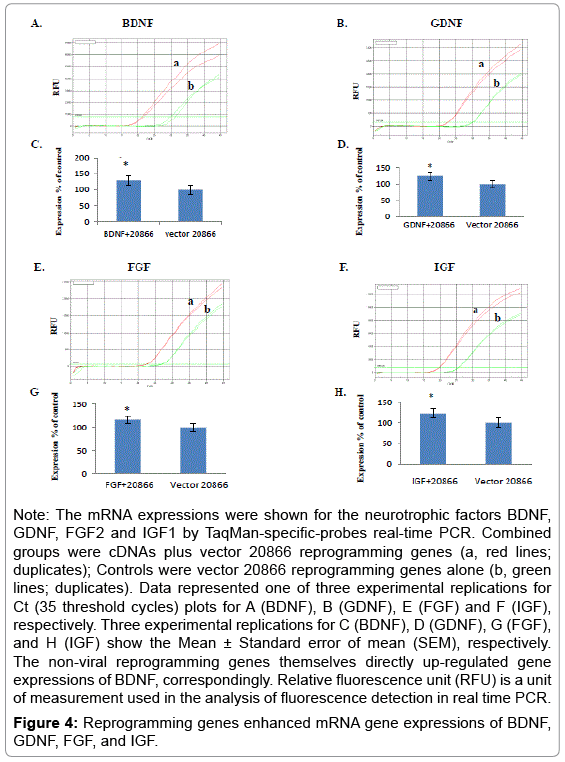
Figure 4: Reprogramming genes enhanced mRNA gene expressions of BDNF, GDNF, FGF, and IGF.
BDNF upheld quantities of reprogrammed neuron cells
Since our data suggested that reprogramming genes themselves can speed up transformation of embryonic brain cells into neuron cells, and the trophic factors BDNF have demonstrated neuroprotective actions in a number of animal models [4,26,31-34], we wondered what effects on neuron cell formations, especially impacts on producing amounts of reprogrammed neuron cells, if combing reprogrammed stem cells as the cell replacement therapy with BDNF as the gene therapy. Two of the critical factors for successful stem cell therapy are to increase in speediness and quantity of producing neuron cells. The early stage reprogrammed neuron cells after transfection with vector 20866 on Day 27 were further co-transfected with/without complete length cDNA BDNF, and then cells were continued for culturing 72 hours with fresh Neural Induction Medium. The amount of producing neuron cells were measured by the western blotting. The data revealed that the cells that were treated with cDNA BDNF plus vector 20866 shown a robust induction of the immature neuronal marker doublecortin at protein level, suggesting significantly higher (3.5 folds higher than vehicle control group), while the group vector 20866 alone was also increased with 2.2 folds when compared with the control group (Figure 5). Reprogramming genes and neurotrophic factors fuel stem cells.

Figure 5: BDNF boosts doublecortin protein expression of reprogrammed neurons in the Western blotting.
The reprogrammed stem cells present greatly promises to downstream clinical applications for restoring, maintaining, or enhancing tissue and organ function for neurodegenerative disorders, such as Parkinson’s disease [1], Alzheimer’s disease [35], multiple sclerosis [36], spinal cord injury [37], stroke [38], and possibly neuropsychiatric illnesses [4,26]. However, for real-world clinical transition there are concerns about technologies in quantities and quickness of neuron products, including differentiated progressive staging neuronal cells generated by reprogrammed stem cells, and potential cancer risks for viral vector carrying reprogramming genes. Recently, the report of high-yield culturing of the human limbinnervating motor neuron has suggested making some progression in the aspect [39].
Numerous reports suggested that BDNF [31], GDNF [32], FGF [33] and IGF [34] have neuroprotective utilities and endorse neuroplasticity beneficial for neuropsychiatric diseases [4,26]. Growth factor-activated stem cells were accelerated by the stromal signals for iPSC reprogramming of human myeloid progenitors [40].
To end of these points above, we have studied: 1) Impacts of nonviral reprogramming genes themselves on speediness of reprogrammed embryonic brain cells being converted into staged neuron cells; 2) Influences of reprogramming genes themselves on gene expressions (amounts) of BDNF, GDNF, FGF and IGF, and evaluating the gene expressions of these neurotrophic factors, since they are key neurotrophic factors; And, 3) Reprogramming genes and neurotrophic factors fuel stem cells. Effect of BDNF, as most critical neurotrophic factors, on reprogramming genes for producing amounts of neuronal cells.
Our data firstly showed that the reprogramming genes with nonviral plasmid vector 20866 successfully converted mice embryonic brain cells into iPSCs, which have been proven by expressions of stem cell biomarkers: alkaline phosphatase stain (Figure 2b), stage-specific antigens SSEA-1 and GF141 (Figure 2c and 2d); as well as the vector 20866 mOrange-positive marker with iPSCs (Figure 2f). The nonviral four reprogramming genes were constructed by Kaji et al [27] for the initial purposes of transformation of fibroblasts into iPSCs. As a technological approach, this plasmid vector with reprogramming genes has extremely possibility to residually stay (cannot 100% clean out or loss of functions) on living brain tissues after transplantation of reprogrammed stem cells, including different staged neuronal cells when real-world clinical applications, even it may be stated that this vector is subsequent excision of reprogramming factors [27]. Thus, this attracted us to pay a great attention to the effectiveness of reprogramming cassette on more natural brain tissues/cells. Our evidence have proven this plasmid vector is no integration into host genomic DNA and no overexpression of Oct4, Sox2, Klf4, and c-Myc in the dopaminergic cells in a long-term (6 months) of cell culturing (unpublished data).
Further, our results suggested that the powers of non-viral reprogramming genes themselves speeded up the transformation of reprogrammed embryonic brain cells into progressive staged neuron cells. The ratio of speeding up for Progenitor neuron cells, Reprogramming genes and neurotrophic factors fuel stem cells. Neuron cells, and Neuron network were 3.2, 2.8, and 1.5 times by reprogrammed cell groups comparing with the control groups, respectively (Figure 3). This functional role would set up a solid base for high efficiency to acquire salvage for the precarious situation of brain neuronal cell damages and win time to repair. Although this exact mechanism is unclear and we needs to do exploratory works in the future, it is not difficulty to understand that adding reprogramming transcript factors into cells may induce more and higher activities in the differentiations and proliferations than relatively “quiet/silent” of natural gene actions. Of note, this rapidity is not equal scales: Progenitor neuron cells were faster than Neuron cells; Neuron cells stood quicker than Neuron network. The reprogramming transcript factors may play more functional roles on the primary initial steps of developing courses of living cell fates.
To the model of this study, we selected the primary mice embryonic brain tissues, but not used adult fibroblast cells or other somatic cells. This is because at this time we wondered whether or not the effectiveness of reprogramming genes themselves were associated with a few important neurotrophic factors such as BDNF, GDNF, FGF2, and IGF-1 in brain tissues. Indeed, the adult mammalian brains still deposit a numbers of the limited embryonic stem cells for replacements for aged or damaged neuronal cells [41]. Thus, this model may be much closer to real-world clinical stem cell transplantations into patients’ brain organ. We are eager to know what influences of reprogramming genes themselves on neuronal tissues and neurotrophic factors in brain tissues. By TaqManspecific- probes real-time PCR, our data displayed that this non-viral reprogramming genes themselves directly enhanced gene expressions of BDNF, GDNF, FGF and IGF as Reprogramming genes and neurotrophic factors fuel stem cells combining co-transfections, when compared with each control vector 20866 with reprogramming genes alone (Figure 4). Based on these data, reprogramming genes themselves would be powerfully sponsoring for producing neurotrophic factors and then promote neutralizing the disease-related cellular and plasticity deficits. Over-express trophic factors would be expected to facilitate the development of new brain circuits, and engineering them to express one of several trophic factors has been linked to neuron survival and regeneration [18].
On the other hand, as the most important neurotrophic factor BDNF sustains much higher amounts of reprogrammed neuron cells through an action on reprogramming genes, while reprogramming genes alone suggested some increased volumes when compared with the control group (Figure 5). The immature neuronal marker doublecortin at protein level were significantly up-regulated by higher 3.5 folds of group cDNA BDNF plus vector 20866, and 2.2 folds of group vector 20866 alone, respectively, than the control group (vehicle). These results suggested that BDNF fueled-up formation of reprogrammed neuron cells in high magnitude style. Obviously, there are great benefits for the rescue of damaged brain cells by reprogrammed stem cells for cell replacement therapy.
The transplantation of the fetal midbrain substantia nigra and dopaminergic neurons in patients with Parkinson’s disease is predominantly remarkable, since 1928 the human fetal tissues and stem cells have been globally used to treat various conditions [42]. However, the clinical reliance on fetal transplantation is gradually decreasing because of the ethical disputes. On other hand, the results of previous fetal neural transplantation Reprogramming genes and neurotrophic factors fuel stem cells therapy for PD have suggested more biologically defined and clinically reliable sources for the dopaminergic neurons. So, this may be required as the parallel control in the clinical trials and may be currently the sole benchmark for exploring applications of other new stem cell sources such as reprogrammed peripheral tissues [42,43]. Furthermore, the derived human iPS cells from fetal neural stem cells were continually applied as a tool for studying of reprogrammed stem cell approaches [43,44]. Of interest, a current good manufacturing practice (cGMP) cell expansion technique to amass large numbers of cells from the fetal brain tissue for research experiments and clinical trials has been developed for the greatly benefiting clinical-grade cell products [45]. The creation of conception after implanting uterine wall through the eighth week of development is devoted to as the embryo in humans, however from the ninth week to birth, the embryo is considered a fetus [42]. In our study, the primary embryonic brain tissues isolated 14 days pregnant mice were a standard application in the early stage of the embryonic tissues used as the description in the Material Section in order to avoid ethical controversies as possible as we can. The reprogrammed primary embryonic brain tissues to derive into iPSCs and progressive differentiated neural cells are an alternative and unique approach, which may be closer to the natural status of the brain and is diverse from other reprogrammed peripheral tissues.
In summary, our report present that the non-viral reprogramming genes themselves accelerated rapidness of converting reprogrammed embryonic brain cells into progressive staged neuron cells; greatly enhanced gene expressions of these important neurotrophic factors BDNF, GDNF, FGF and IGF; and furthermore BDNF, as most key neurotrophic Reprogramming genes and neurotrophic factors fuel stem cells factors, directly up-regulated producing neuronal cells through additional role with reprogramming genes. These data suggested a high efficient method in both quantities and speediness for generating reprogrammed stem cells and further differential neuronal cells for downstream clinical applications.
This work was supported by grants from the Canadian Institutes of Health Research.