Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Review Article - (2013) Volume 1, Issue 1
Prostate cancer remains the leading cause of cancer-related morbidity and mortality for men in the Western world. Conventional anti-cancer therapies like chemotherapy, irradiation, and hormone ablation often slow tumor growth but do not engender long term benefits on patient survival. These therapies are limited by the fact that tumor re-growth and spread to distal sites usually occurs following the conclusion of treatment. Therefore, there is an increasing demand for the development of alternative therapeutic regiments. The use of oncolytic viruses for the treatment of prostate cancer is an attractive option due to the natural ability of viruses to target and kill cancer cells. Furthermore, oncolytic viruses may be genetically manipulated to transfer exogenous genes into cancer cells in order to provide new generations of biological controls. This brief review highlights the potential of select oncolytic viruses as promising modalities for prostate cancer treatments and presents the advantages and practicalities of such viruses as therapeutic agents.
Keywords: Oncolytic viruses, Prostate cancer, Cancer therapies, Adenovirus, Herpes simplex virus, Vaccinia virus, Newcastle disease virus, Vesicular stomatitis virus
Viruses are commonly thought of as opportunistic organisms that hijack cellular machinery with the ultimate goal of replicating and causing destruction in the host. However, viruses are increasingly being explored as therapeutic agents for a variety of disorders and diseases, including cancers, due to their natural ability to spread to various cell types. Oncolytic viruses are replication-competent viruses that have the ability to selectively target cancerous growths, either naturally through adaptation or through genetic manipulation. Th e fi eld of oncolytic virus development has spanned approximately twenty years, with numerous viruses currently in clinical trials and in various stages of development as anti-cancer agents. Although oncolytic viruses exhibit diversity in their biologics and host range, oncolytic viruses share common features of selective replication in tumors, eff ective lysis of tumor cells, either directly or through activation of anti-tumor immunity, and dispersion within tumor masses. Strategies employed by viruses to target cancer cells include exploiting the defective antiviral responses in cancer cells, the use of cancer-specifi c surface markers as viral entry receptors, as well as the use of cancer-specifi c promoters to activate viral gene products. Many viruses that are currently being developed as agents against prostate cancers have taken advantage of these strategies.
Depending on the disease state, the current treatments for prostate cancer include radiotherapy, surgery, and hormone-deprivation therapy [1]. Prostate cancer is particularly well-suited for oncolytic therapies due to the fact that prostate removal or ablation is not life- threatening. Furthermore, the major cause of death from prostate cancers results from metastatic spread. Th erefore, the natural ability of viruses to spread to distal sites and seek out susceptible cancerous tissues makes them attractive therapeutic agents for this disease. Th e response to oncolytic therapies may also be monitored by serum prostate-specifi c antigen (PSA) levels. Th is review will focus on select oncolytic viruses and the advances that have been made in developing them as therapeutic agents for the treatment of prostate cancer.
One of the most widely studied oncolytic viruses is adenovirus. Th is virus was initially isolated in the early 1950s from adenoid-infected cell cultures, thus leading to the name adenovirus. It is recognized as the etiological agent for diverse syndromes due to the presence of approximately 50 serotypes. Not only does adenovirus possess inherent oncolytic activity, it is widely being developed as a vaccine and gene therapy agent.
Adenoviruses are attractive therapeutic vectors due to their wide host range and the ease by which they may be manipulated genetically [2]. Furthermore, a decade worth of clinical trials have tested the safety and effi cacy of various adenoviral vectors, thus providing a framework from which to engineer novel agents. As an oncolytic virus, much research has revealed that unmodifi ed adenovirus is insuffi cient to eff ectively treat neoplastic tissue due to a variety of factors, including clearance from the immune system, hepatic virus sequestration and down regulation of the coxsackie-adenovirus receptor (CAR) in tumor cells [3-5], thus leading to the engineering of second- and third- generation viruses with greater therapeutic effi cacy.
Th e fi rst oncolytic adenovirus used to treat human cancers is ONYX-15, containing a deletion of the viral EIB 55 KD gene. Th e lack of EIB 55 KD expression restricts ONYX-15 replication and killing in cells infected in the G1 phase of the cell cycle [6-8]. Th is poses a limitation to cancer therapies because a signifi cant number of cells within tumors exist in the G1 phase. Because of this, ONYX-15 therapies have been more successful when combined with chemotherapy or radiation therapies [7,9]. Additional adenoviruses with mutations in the E1A gene product have been generated. E1A stimulates S phase entry and serves to transactivate both cellular and viral genes essential for a productive viral infection [10]. For targeting prostate cancers, investigators have taken advantage of prostate-specifi c promoters and inserted them upstream of the EIA gene of adenovirus type 5 (Ad5), thus restricting viral replication to prostate cancer cells. An example of such a virus is CN706 which was created by inserting the prostate specific enhancer (PSE) derived from the 5’ end of the PSA gene into Ad5 [11]. In treatments with CN706, virus replication correlates with the level of PSA expression within given prostate cancer cells. Studies show that this virus is effective at restricting growth of PSA-producing LNCaP prostate tumors in cell culture and animal model studies. Numerous studies have also shown efficacy by targeting prostate cell surface markers that are shown to be up-regulated during tumorigenesis. Prime candidates include prostate-specific membrane antigen (PMSA), whose increased expression correlates with aggressive tumors, prostate stem cell antigen (PSCA), the urokinase-type plasminogen activator receptor (uPAR), which is involved in tumor angiogenesis [12-15], and differential display code 3 (DD3 or DD3(PCA)) [16]. ONYX15 and the prostate-specific adenoviruses illustrate the two main strategies employed to restrict oncolytic adenovirus replication to tumor tissues: 1) By inactivating viral genes whose functions can be compensated in cancer cells, and 2) By placing essential viral genes under control of cancer or tissue-specific promoters [17].
In addition to the success of engineered adenoviruses utilizing targetable prostate cancer-specific receptors, prostate cancers have been targeted with viruses containing the promoter for human telomerase reverse transcriptase (hTERT), the catalytic componenent of the telomerase ribonucleoprotein complex found in cancer cells. An example of such a virus is OBP-301, which shows strong anticancer effects by inducing the lysis of human prostate cancer cells and also demonstrates antimetastatic effects by eradicating detectable contralateral LNCaP tumors in vivo [18]. More recently, Hu et al. [19,20] developed an hTERT promoter-containing adenovirus engineered to express sTGFβRIIFc, a protein which directly targets and inhibits the TGF-β pathway. TGF-β has been shown to play an important role in the control of bone metastases [21,22] and high levels of this factor in the blood circulation are poor prognostic markers of prostate cancer [23,24]. This virus induces significant reduction of tumor burden, osteoclast number and bone destruction in a bone metastasis mouse model [19], thus displaying its potential as a therapeutic for prostate cancer metastases.
Further therapeutic regiments employed for prostate cancers include immune and suicide gene therapies. Adenoviruses have been developed to express cytokines, chemokines, tumor-associated antigens or other immunomodulatory factors. For example, adenoviruses armed with immune-therapeutic genes such as IL-12 and IL-24 have shown some efficacy in preclinical studies for the treatment of prostate cancers [16,33]. Immune cells including macrophages have also been utilized to deliver adenoviruses to hypoxic areas of prostate tumors [31]. In terms of suicide gene therapy, the two most widely used prodrug therapies for prostate cancers include HSV thymidine kinase (HSV-tk) together with ganciclovirir (GCV) or acyclovir, cytosine deaminase (CD) and 5-fluorocytosine (5-FC) [34,35]. Each of these therapeutic regiments represents a targeted approach for prostate cancers that have acquired resistance to conventional treatments.
Herpes simplex virus type I (HSV-1) is a natural human pathogen which has been studied as an oncolytic agent for over two decades. During this time, increased strides have been made in developing HSV for the treatment of a variety of different cancers. This progress is highlighted by the translation of at least six oncolytic HSV vectors to the clinic, some having progressed to Phase II/III clinical trials [36]. A benefit to oncolytic HSV-1 therapies is the availability of anti-HSV specific drugs (acyclovir) that may be administered upon detection of a life threatening infection. Early studies of oncolytic HSV focused on developing safe anti-cancer agents by deleting the γ34.5 gene, which governs neuropathogenicity [37,38]. Further vectors were developed by introducing mutations or deletions in specific genes to prevent reversions to wild-type strains. However, these changes led to limited success due to attenuation of replication in susceptible tissues including prostate carcinoma cells [39], indicating that greater potency was necessary to promote oncolytic efficacy. Current studies are focusing on synergizing the effects of oncolytic HSV with a variety of agents.
Several oncolytic HSV-1 strains have shown promise at treatment of prostate cancers. G207 is one of the first onoclytic HSV-1 strains taken into clinical trials. This virus, derived from strain F, contains deletions in both copies of the γ34.5 gene and has an inactivated ICP6 gene, which encodes a viral ribonucleotide reductase function [40]. The double mutations permit viral replication within quiescent tumor cells carrying specific oncogene deletions but not in normal cells [41]. G207 has been shown to be effective at killing human prostate cancer cells in vitro, as well as in vivo in both subcutaneous xenograft and transgenic mouse models [42-44]. Additionally, it displayed no evidence of clinical disease and virus spread into other organs when injected into the prostates of HSV-1 susceptible mice and non-human primates [45]. NV1020 is a multimutant HSV-1 strain that contains several genetic modifications including deletion of the UL24 gene and one copy of the γ34.5 gene [46]. This virus has also shown efficacy in reducing prostate tumor growth in vivo and significantly decreasing serum PSA levels [47]. Additional attenuated, replication competent viruses derived from first generation oncolytic HSV-1, such as NV1023 and G47Δ, are being evaluated for their ability to promote greater antitumor activity against prostate cancers [43,48].
In addition to testing the ability of oncolytic HSV-1 strains to induce tumor cell killing, they have also been used as a platform to deliver transgenes of interest. The integration of membrane-fusion activity into these viruses has been shown to promote anti-tumor effects in prostate cancer cells [29,49]. Furthermore, viruses have been armed with agents commonly used for prostate cancer vaccinations such as prostatic acid phosphatase (PAP) [50], immune modulators such as IL-12 [51], as well as factors that enhance virus replication such as Ing4 (inhibitor of growth 4) [52,53].
Studies have focused on enhancing virus replication at prostate cancer tissues as well as exploring combination approaches. Lee et al. [54] have developed recombinant viruses whose expression is regulated by the presence of the prostate specific promoter (AAR(2)PB) and the 5’UTR of rFGF-2, thus promoting tumor specificity. G47Δ, a multimutated, replication competent HSV-1 vector derived from G207, was engineered by creating an additional deletion within the non-essential α47gene [54]. The combination of androgen ablation with G47D therapy resulted in greater tumor growth suppression than either therapy alone in the TRAMP-C2 subcutaneous model. These are a few of the many examples of approaches to enhance the oncolytic potential of HSV-1 vectors, similar to those outlined for adenoviruses.
Vaccinia virus is a large, enveloped virus belonging to the poxvirus family. The study of vaccinia virus began with its popularity as the choice for smallpox vaccination and in its role in the successful global eradication of smallpox by 1979. Since then, there has been great interest in developing vaccinia virus a vector for the expression of foreign genes. This virus is attractive as a delivery vehicle because of its ability to stably accept as much as 25 kb of foreign DNA, thus enabling it to express large genes. Furthermore, studies have shown that it is able to enter and replicate efficiently within numerous cell types without causing natural disease in humans [55]. However, to promote safety, attenuated, avirulent versions of vaccinia viruses, including those lacking replication capacity, have been utilized as delivery vectors for gene therapies or as vaccine vectors for the expression of immunizing antigens. More recently, vaccinia virus has also gained popularity as an anti-cancer agent. Beginning in 2007, Zhang et al. [56] described the oncolytic potential of the attenuated recombinant vaccinia virus GLV-1h68 in breast tumors. Since then, the oncolytic effect of this virus has been demonstrated in numerous cancer models, including in the treatment of lymph node metastases originating from prostate carcinoma cells [57].
Current studies are interested in determining the mechanisms by which GLV-1h68 promotes anti-cancer activity. GLV-1h68 was engineered by inserting three expression cassettes into different loci of the viral genome. Further genomic analysis confirmed that these insertions reduced the virulence of this virus and promoted cancer cell tropism [58]. Recent studies have attributed the ability of GLV-1h68 to effectively treat lymph node metastases of prostate carcinoma cells to the elevated vascular permeability in metastases leading to greater release of virus particles and spread to susceptible tissues [59]. Furthermore, the presence of increased number of immune cells and the proliferation of cancer cells at metastatic areas are thought to provide favourable conditions for virus infection and replication. Taken together, these data indicate that vaccinia virus GLV-1h68 may be used for the preferential destruction of metastatic prostate carcinoma cells, which represent a major cause of cancer-related deaths.
In addition to GLV-1h68, a recombinant vaccinia virus expressing PSA (rV-PSA) was constructed by inserting the PSA gene into the viral genome of the Wyeth strain of vaccinia. rV-PSA has shown some success in Phase I clinical trials as indicated by limited toxicity and evidence of immunological activity in patients with rising PSA levels after local therapy, and in patients with metastatic androgen-independent prostate cancer [60,61].
Newcastle disease virus (NDV) is a negative-sense single-stranded virus that causes deadly infection in various species of birds but is non-pathogenic to humans and domestic animals. NDV has been applied for the treatment of human cancers since the early 1960s with studies on uterine carcinoma [62]. Since then, it has been reported to possess oncolytic activity against a range of cancer types and various strains have been tested in clinical trials in different human cancers including glioblastoma multiforme and colorectal cancer [63-65]. Studies have shown that NDV exhibits inherent selectivity for a diverse group of tumors over normal cells due to defects in antiviral responses, such as the type I interferon (IFN) response, in certain cancer cells [66,67]. However, it has also been proposed that tumor specificity may be dependent upon tumor cell resistance to apoptosis [68]. These tumor-specific defects serve to enhance replication of NDV in cancer cells to promote virus-induced cytotoxicity.
The mechanisms underlying the antitumor activity of NDV have been investigated in numerous studies. Multiple studies have revealed the role of apoptosis in cell death by NDV, including both the intrinsic and extrinsic pathways. The exact mechanisms of apoptotic death are dependent on the strain of NDV, the cell lines and the detection assays [69-74]. In addition to direct killing induced by the virus, NDV also stimulates robust innate and adaptive immunity. Various strains of NDV are capable of stimulating macrophage activity as indicated by the detection of macrophage enzymes such as iNOS, lysozyme and acid phosphatase as well as the production of nitric oxide and TNF-α [75-77]. Natural killer (NK) cells have also been shown to mediate cytotoxicity against multiple tumor cell lines following infection of peripheral blood mononuclear cells (PBMCs) with NDV strain 73-T, one of the most well-characterized oncolytic strain of NDV [78].
NDV as an oncolytic agent for the treatment of prostate cancer is currently in the early stages of development. Studies with NDV 73-T have demonstrated antitumor effects in prostate carcinoma (PC3) xenografts upon systemic administration [79]. Furthermore, significant inhibition of tumor growth (77-96%) was also observed in epidermoid, colon, large cell lung, breast and low passage colon carcinoma xenograft models. Although phase I clinical trials using naturally attenuated NDV strains such as PV701 have been conducted, they have not included patients with prostate cancers. Nevertheless, clinical trials have revealed that PV701 was well tolerated by patients when administered intravenously [35]. Side effects, including flu-like symptoms, localized adverse effects at the tumor site and infusion reactions, were observed. However, there was no toxicity from the oncolytic virus treatment.
Infection with NDV is dependent on two viral glycoproteins; hemagglutininneuraminidase (HN) and fusion (F). A major determinant of virulence is the cleavage site in the F protein, which becomes fusogenic only upon proteolytic cleavage into two disulfide-linked polypeptides by host cellular proteases [80]. In an attempt to improve antitumor efficacy, Shobana et al. [81] have engineered the F protein cleavage site to target the serine protease, PSA, such that F protein is cleavable exclusively by PSA in prostate cancer cells. This strategy enhanced pathogenicity of oncolytic NDV in prostate cancer cells as a result of restricted viral replication and fusogenicity [82].
Vesicular stomatitis virus (VSV), a negative-strand RNA virus of the Rhabdovirus family, has been studied as an anti-cancer agent for several years. VSV exhibits numerous properties of an effective oncolytic agent including its well-defined biology, ability to induce apoptosis in a wide array of cancer cells and the lack of preexisting immunity in humans [67,82-85]. Similar to Newcastle disease virus, it has been proposed that the susceptibility of tumors to VSV is due to development of defects in antiviral responses during tumorigenesis [67,82,85-87]. While normal cells may be infected by VSV, they respond to the virus by enhancing the type I IFN response, leading to the attenuation of virus replication. However, wt strains of VSV have the ability to suppress the antiviral response, induce systemic immunity and replicate in the central nervous system [86,88,89], thus leading to safety concerns. Over the last decade, increasing strides have been made in the understanding of the interaction between the virus, cancers, and the immune response. This has led to the development of a number of recombinant attenuated VSVs with the goal of enhancing the oncolytic potential of the virus, either directly or indirectly through stimulation of the immune response, while maintaining safety.
VSV has been tested as a candidate oncolytic virus for prostate cancer by several groups. Early studies with prostate cancers tested the ability of a matrix (M) protein mutant of VSV (rM51R-M virus) to kill LNCaP and PC3 prostate cancer cells in cell culture and xenograft model systems [86]. The M51R M protein mutation disrupts the ability of VSV to shut-off the host antiviral response in infected cells [85,86,90]. Xenograft studies showed that rM51R-M virus exhibits enhanced selectivity for tumor over normal cells as compared to wt VSV strains, as indicated by the ability of the virus to effectively kill tumor cells with limited signs of disease [86]. However, the efficacy of the virus depends on the cell type. LNCaP cells are extremely sensitive to the effects of the virus while PC3 cells remain resistant to infection and killing by rM51R-M virus perhaps due to the constitutive expression of numerous antiviral gene products in this cell line [91] (Figure 1).
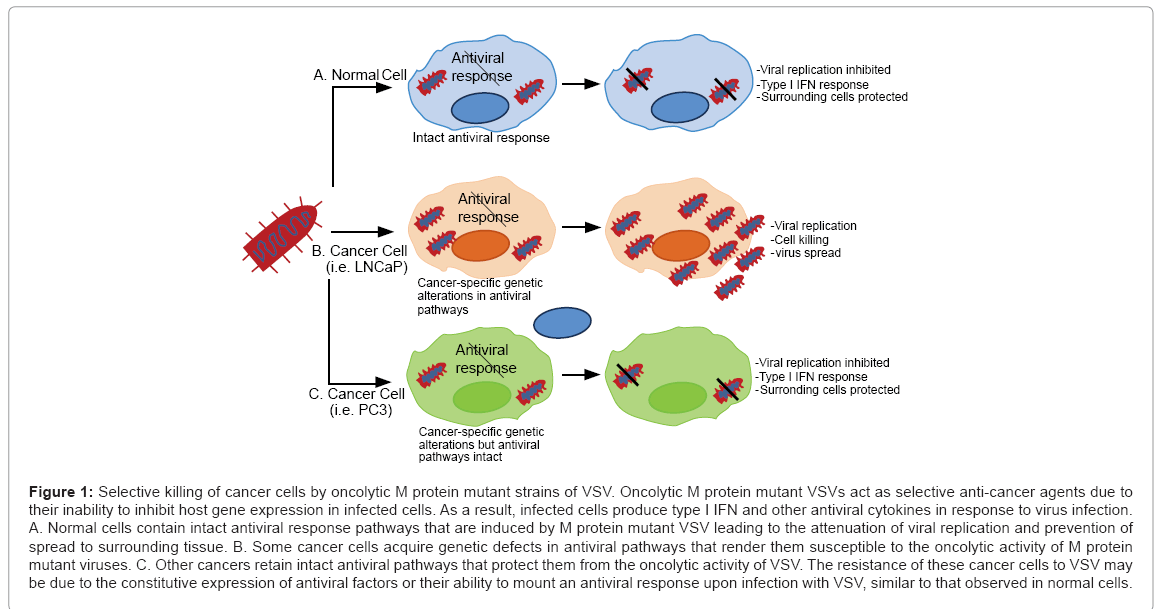
Figure 1: Selective killing of cancer cells by oncolytic M protein mutant strains of VSV. Oncolytic M protein mutant VSVs act as selective anti-cancer agents due to their inability to inhibit host gene expression in infected cells. As a result, infected cells produce type I IFN and other antiviral cytokines in response to virus infection. A. Normal cells contain intact antiviral response pathways that are induced by M protein mutant VSV leading to the attenuation of viral replication and prevention of spread to surrounding tissue. B. Some cancer cells acquire genetic defects in antiviral pathways that render them susceptible to the oncolytic activity of M protein mutant viruses. C. Other cancers retain intact antiviral pathways that protect them from the oncolytic activity of VSV. The resistance of these cancer cells to VSV may be due to the constitutive expression of antiviral factors or their ability to mount an antiviral response upon infection with VSV, similar to that observed in normal cells.
Several M51 protein mutants of VSV have been used to explore combination approaches for the treatment of a variety of cancers, including prostate cancers. In an attempt to augment the ability of VSV-Δ51-GFP to kill VSV-resistant PC3 cells, Nguyen et al. [92] pretreated prostate cancer cells with histone deacetylase inhibitors (HDIs) known to suppress the type I IFN response [93]. Using the HDIs, HDI-MS-275 and SAHA (Vorinostat), which have shown promising anti-cancer results in preclinical or clinical trials, they were able to augment the oncolytic activity of VSV-Δ51-GFP both in vitro and in vivo xenograft models. Another approach involved engineering the recombinant (VSV)-MΔ51 virus to express the cytosine deaminase/uracil phosphoribosyltransferase (CD::UPRT) suicide gene and 5-fluorocytosine (5FC) prodrug [93]. This virus had an enhanced ability to kill PC3 cells as compared to viruses lacking the suicide gene. Furthermore, it was effective at killing additional tumor cell lines derived from the breast. These, and similar studies with other oncolytic viruses demonstrate the concept that in order to promote oncolysis, synergistic combination approaches must be investigated.
Immunocompetent transgenic mice have served as useful model systems for measuring the safety and efficacy of VSV treatment of prostate tumors. Moussavi et al. [94] demonstrated that an IFN-sensitive VSV (AV3 strain) expressing luciferase effectively spreads in tumor-bearing prostate-specific PTEN(-/-) mice to selectively infect and kill prostate tumor cells while sparing normal cells in control mice [95]. In these studies the virus was injected at the prostate site, thus demonstrating the utility of this administration route. More recently, this same group showed that AV3 effectively targets metastatic lesions arising in the transgenic adenocarcinoma of the mouse prostate (TRAMP) model [95]. The TRAMP C2 cell line derived from TRAMP mice was also utilized to demonstrate the enhanced oncolytic properties of a recombinant VSV encoding SV5-F able to induce syncytial formation [96]. The SV5-F recombinant virus was constructed by replacing VSV glycoprotein (G) with that of the SV5-F to generate rVSV-DeltaG-SV5-F. rVSV-DeltaG-SV5-F virus replication was restricted to TRAMP-C2 tumors where it showed enhanced apoptotic and cytotoxic effects relative to a control virus lacking SV5-F.
In order to direct VSV to prostate cancer cells, investigators have pseudotyped replication defective VSV lacking its glycoprotein (VSVΔG) with MV-F and MV-H displaying single-chain antibodies (scFv) specific for or prostate membrane-specific antigen (PSMA) [97]. Results indicated that VSV replication was restricted to prostate cancer cells expressing the PSMA surface marker. In addition, upon engineering VSV to express antibodies for the epidermal growth factor receptor (EGFR) and folate receptor (FR), this group confirmed that retargeted VSV only replicated in cells expressing the target receptor. Therefore, taking advantage of the presence of cancer-specific surface markers represents and effective strategy to restrict the ability of the virus to kill cancer cells over normal cells.
Oncolytic viruses represent promising modalities for the treatment of prostate cancers due their ability to seek out and infect tumors, replicate in target cells and spread to surrounding cancerous or metastatic tissues. Numerous strategies have been employed to enable oncolytic viruses to selectively target, replicate in and kill cancer cells. Promising approaches include the targeting of viruses to prostate tumor-specific surface markers or the use of tumor-specific promoters to restrict viral gene expression to prostate tumors. In order to enhance killing of cancer cells, suicide gene therapies such as the use of HSV thymidine kinase and gancyclovir have been explored. In addition, the delivery of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) to tumors has been shown to enhance the release of progeny virions from infected cells in order to promote spread of virus to surrounding susceptible tissue. Because the efficacy of oncolytic therapies is greatly dependent on the enhancement of anti-tumor immunity, various methods to modify the immune response have been tested, including the viral delivery of immunostimulatory molecules and the co-administration of reagents to enhance immune function. Additionally, the multifaceted tumor environment has been shown to impact viral infection, replication and spread within the tumor. Therefore, studies are also exploring means to overcome environmental restrictions imposed on oncolytic therapies. Each of these approaches offers great promise, either alone, or in combination with established antitumor therapies such as radiation or chemotherapy. Therefore, together with results obtained from numerous clinical trials, the future of oncolytic therapies for prostate cancers remains promising.