Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 5
Aim: The objective of this study was to evaluate the prophylactic and therapeutic effects of Krill Oil on alcohol induced fatty liver in rats on high-fat diet. Methods: Male Sprague-Dawley rats (7-8 weeks old) were gavaged with alcohol and fed with high-fat diet for six weeks to induce alcoholic fatty liver. The concentration of the alcohol formulation had a step increase to have the animal acclimated, with 8% in week 1, 12% in week 2, 20% in week 3, 30% in week 4, and 40% alcohol in the following weeks, and the dose volume was 10 ml/kg. Krill oil at daily oral dose of 10, 100 or 1000 mg/kg was given to animals along with alcohol gavage in a prophylactic experiment, or at 100 and 1000 mg/kg after six weeks of alcohol gavage in a therapeutic experiment, with or without continuing oral gavage of alcohol for four weeks. Blood samples and liver tissues were taken for triglyceride and total cholesterol assays, liver tissues were taken for microarray analysis and histopathological evaluation of hepatosteatosis at the end of the treatment. Results: Krill Oil dose-dependently inhibited the elevation of liver triglyceride level from 17. 8% to 66.3% compared with control, and the incidence and severity of liver cytoplasmic vacuolization. In the therapeutic experiment, Krill Oil group showed an accelerated reduction of liver tissue triglyceride level and incidence and severity of liver cytoplasmic vacuolization. Krill Oil also reversed the elevated liver tissue triglyceride and the elevated incidence and severity of liver cytoplasmic vacuolization in rats which had been given alcohol for ten weeks with Krill Oil treatment initiated at the beginning of week 7. Conclusion: Our results suggest that Krill Oil supplementation not only has prophylactic effects on alcoholic fatty liver formation, but also has potential therapeutic effects on the alcoholic fatty liver.
<Keywords: Alcoholic fatty liver; Krill Oil (KO); Rat; Polyunsaturated fatty acids (PUFA); Histopathology
Chronic and excessive alcohol consumption is a major cause of fatty liver, which is a problem in health care [1]. It is also classified as alcoholic fatty liver, an early stage of alcoholic liver disease (ALD) [2]. Over 60% heavy drinkers will develop fatty liver [3] and, if left untreated, the fatty liver will gradually aggravate and progress with high likelihood to more serious diseases, i.e. steatohepatitis, fibrosis and cirrhosis [1]. According to a WHO report [4], the deaths of about 50% of cirrhosis of the liver and about 30% of liver cancer are attributable to alcohol consumption. Fatty liver is a reversible condition where large vacuoles of triglyceride fat accumulate in liver cells via the process of steatosis at levels higher than normal, i.e. >5% of hepatocytes are steatotic. Pathogenesis of fatty liver also involves excessive calorie intake, dietary imbalance, inadequate exercise, and is associated with obesity, type 2 diabetes mellitus, coronary heart disease, hypertension, and dyslipidemia, which are all classified as non-alcoholic fatty liver [5-7].
As ambulatory patients with alcoholic fatty liver are often asymptomatic, and for most of the heavy alcohol consumption is not evitable due to physical dependency or the need of social drinking, thus the dietary supplementation becomes one of the best choices to prevent or control alcoholic fatty liver. Among which the most mentioned and reported are polyunsaturated fatty acids (PUFA). Many studies indicated that ω-3 fatty acids, the most important families of PUFA, have protective effects on fatty liver diseases [8-12]. PUFA are constituents of the animal and vegetable fats found in food. They are considered essential fatty acids, meaning that they cannot be synthesized by the human body but are vital for normal metabolism. They have to be provided from food or with an appropriate supplement [13,14]. Common sources of ω-3 PUFA include fish oils, algal oil, squid oil and some plant oils such as echium oil and flaxseed oil.
As another important source of PUFA, Krill Oil has received increasing attention in recent years for its beneficial effects on human health. Krill Oil is a mixture extracted from Antarctic krill (Euphausia superb Dana). Krill Oil contains a high proportion of ω-3 fatty acids, mainly Eicosapentaenoic Acid (EPA, C20:5) and Docosahexaenoic Acid (DHA, C22:6) which are bound to phospholipids [15,16], in contrast to traditional ω-3 supplements such as cod liver oil and fish oil which are based on ω-3 fatty acids that bound to triglycerides. By binding with phospholipids, long-chain ω-3 fatty acids have a higher level of passage through the intestinal wall [17]. Besides ω-3 fatty acids, Krill Oil also contains many kinds of antioxidants including Vitamin E, all-trans-retinol Vitamin A, beta carotene and the carotenoid relatives, canthaxanthin and astaxanthin [17].
While the prophylactic effects of the Krill Oil on the non-alcoholic fatty liver had been proved in high-fat-fed mice and rats [18,19], the beneficial effects of Krill Oil on the alcoholic fatty liver are speculated but not yet demonstrated in pre-clinical studies. In this study, both prophylactic and therapeutic effects of Krill Oil on the pathogenesis of alcoholic fatty liver in rat model were investigated. The alcoholic fatty liver rat model was induced with daily alcohol gavage along with high fat diet feeding.
Chemicals and reagents
Krill Oil and Medium Chain Triglyceride (MCT) were obtained from Nippon Suisan Kaisha, Ltd., Tokyo, Japan. Alcohol (95%) was purchased from Taicang Xintai Alcohol Co. Ltd. Normal diet (ND) was purchased from Beijing Keao Xieli Diet Co. Ltd. High fat diet (HFD, fat content: 23%) was purchased from Beijing HFK Bioscience Co. Ltd. All other chemicals and solvents were of the highest grade commercially available.
Animals
Male Sprague-Dawley rats (SPF grade) were used in the whole study (Permit Numbers: ACU-11-491 and ACU12-142). Rats (160-220 g) were purchased from Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). Rats were housed in a controlled environment (temperature 22-26°C, humidity 40-70%, and 12 hour light/12 hour dark cycle) in the animal facility of JOINN Laboratories with access to food and water ad libitum unless otherwise specified. Rats were deprived of food (overnight) prior to blood sampling, and prior to terminal euthanasia. Rats were acclimated before use for the experiments.
Alcoholic Fatty Liver Model Preparation and Study Design
Fatty liver in rats was induced by alcohol gavage plus HFD feeding based on Li’s method with some modifications [20]. Briefly, rats were fed with HFD plus orally administered alcohol (10 mL/kg, twice daily, at least 4 hours apart) for 6 or 10 weeks. The concentration of alcohol in the formulation increased with time along with glucose adding in the formulation to have the animal acclimated to the alcohol administration, with 8% alcohol plus 30% glucose in week 1, 12% alcohol plus 20% glucose in week 2, 20% alcohol plus 15% glucose in week 3, 30% alcohol plus 10% glucose in week 4, and 40% alcohol plus 10% glucose in following weeks.
For the prophylactic experiment, rats were randomized into six treatment groups at the initiation of alcohol gavage, and MCT or Krill Oil (diluted in MCT) was given at dose volume of 1 mL/kg via oral gavage once daily for six weeks (Figure 1). MCT, which does not contain ω-3 PUFA, was used as the vehicle control article and the diluent for Krill Oil formulation. Body weights and food consumption data were collected 3 times weekly. At the end of treatment, the rats were euthanized and necropsied. Blood samples were collected for serum triglyceride (TG) and total cholesterol (TC) analyses. Liver tissues were collected for TG and TC measurements, and for histopathological evaluation of hepatosteatosis.
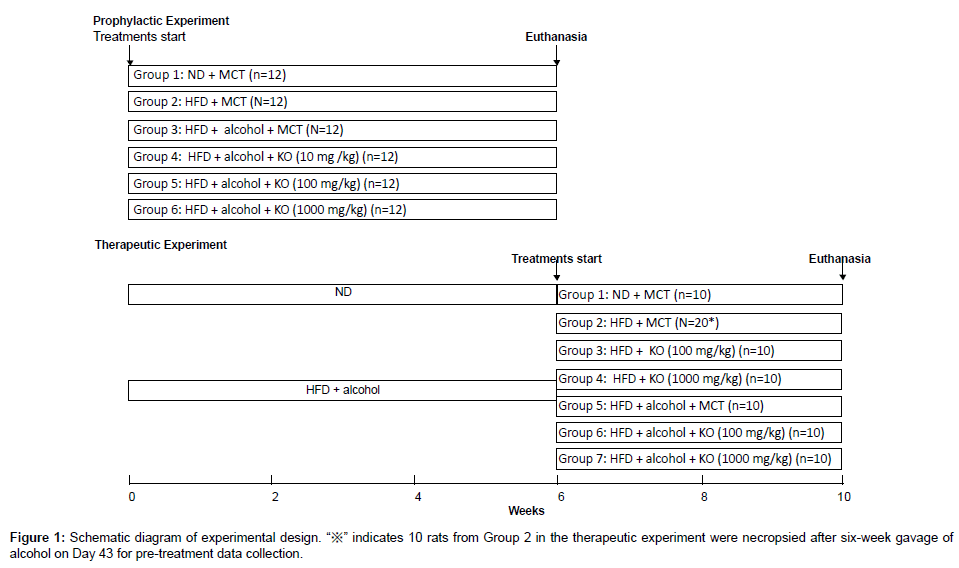
Figure 1: Schematic diagram of experimental design. “※” indicates 10 rats from Group 2 in the therapeutic experiment were necropsied after six-week gavage of alcohol on Day 43 for pre-treatment data collection.
For the therapeutic experiment, rats were randomized into seven groups after six-week gavage of alcohol and 10 rats from Group 2 were euthanized and necropsied on Day43 for pre-treatment data collection, then MCT or Krill Oil (diluted in MCT) was given to the remaining rats at dose volume of 1 mL/kg via oral gavage once daily through week 7 to week 10, with or without continuing oral gavage of alcohol (Figure 1). Body weights and food consumption data were collected twice weekly. At the end of treatment, the rats were euthanized and necropsied. Blood samples were collected for serum TG and TC analyses. Liver tissues were collected for TG, TC measurements, and for histopathological evaluation of hepatosteatosis.
Biochemical analyses
The serum TG, TC concentrations were measured by enzymatic methods, using GPO-PAP and COD-PAP kits (Shanghai Huashi Asia- Pacific Bio-pharmaceutical Co., Ltd). The liver TG and cholesterol were also determined using the supernatant after homogenated with PBS, individual hepatic TG and TC were quantitated enzymatically as described above.
Liver histopathology
Liver specimens from the rats were fixed in 10% neutral buffer formalin and embedded in paraffin. The specimens were sliced at 5 μm and mounted on glass slides. After staining with hematoxylin and eosin, the slides were evaluated microscopically by a veterinary pathologist. Fatty liver (cytoplasmic vacuolization) was graded according to the following grading scale [21]: slight severity (+) was recorded when cytoplasmic vacuolization was observed but was present in ≤1/3 of the hepatocytes in the section; moderate severity (++) was recorded when >1/3 and ≤2/3 of the hepatocytes in the section were affected; marked severity (+++) was recorded when >2/3 of the hepatocytes in the section were affected.
Microarray analysis
In the therapeutic experiment, liver samples from the rats were collected at necropsy and placed into RNAlater (Ambion). The samples from 3 control rats (Group 5) and 3 Krill Oil-treated rats (Group 7) were transferred to CapitalBio for Microarray analysis. Briefly, RNA was isolated from liver samples using the Ambion’s kit following the manufacturer’s instructions. The RNA samples were analyzed for quantity and purity by UV analysis and were evaluated for RNA integrity by gel electrophoresis. The RNA samples (≥1 μg) were used to synthesize double-stranded cDNA using the Affymetrix One-Cycle Target Labeling and Control Reagents kit. Then the cDNA was used as a template to synthesize biotin-labeled antisense cRNA using an in vitro transcription labeling kit. The cRNA was purified and fragmented according to the manual provided with the GeneChip Sample Cleanup module (Affymetrix). All GeneChip arrays (Rat Genome 230 Version 2.0 arrays) were hybridized, washed, and stained according to the Affymetrix Technical Manual. The chips were scanned using an Affymetrix GeneChip Scanner 3000 with Affymetrix® GeneChip® Command Console® Software. CEL files were imported in the R statistical package and were RMA normalized. SAM analysis was performed to identify differentially expressed genes between vehicle and Krill Oil-treated samples. The genes with q-value ≤ 5% and Fold Change ≥2 or ≤ 0.5 were identified as differentially expressed genes.
Statistical analysis
Results are expressed as means ± SD (standard deviation). Comparisons between means were performed using One-way analysis of variance (ANOVA), followed by a post-hoc LSD test. χ2-test was used to evaluate the significances of histopathological evaluation. Differences were considered to reach statistical significance when P≤0.05. Statistical analyses were carried out with SPSS software (version 13.0 for Windows).
Body weight
In the prophylactic experiment, the mean body weight of ND control (ND+MCT) and HFD control (HFD+MCT) groups increased with time and there was no statistically significant difference between these two groups. The body weight of MCT control (HFD+Alcohol+MCT) group also increased with time in the initial 3 weeks. There was no apparent body weight gain in the following 2 weeks followed by body weight gain until week 6. The changes in body weight in MCT control group were considered to be related to the decrease in food consumption. After the animals acclimated to the high concentration of alcohol, the body weight gains recovered partially until week 6. By comparison with MCT control group, no apparent changes in body weight were observed in the Krill Oil treated groups (Figure 2A).
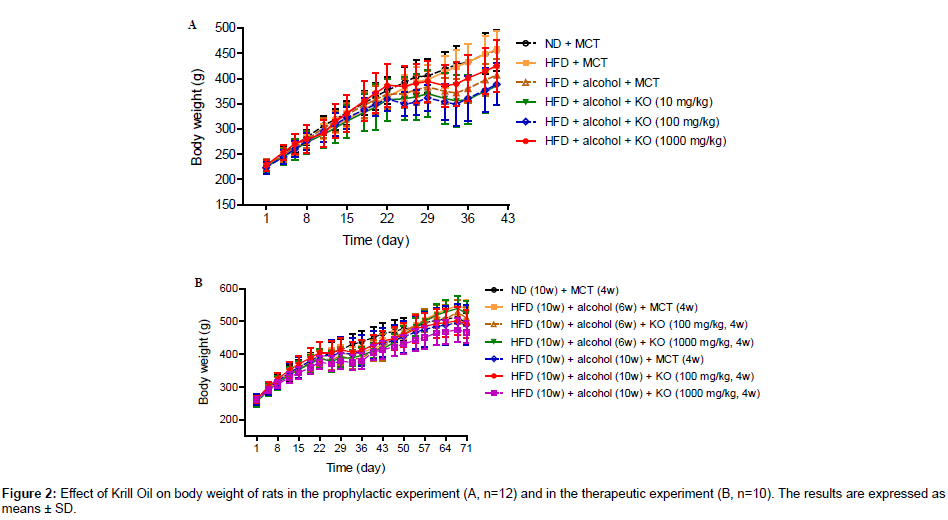
Figure 2: Effect of Krill Oil on body weight of rats in the prophylactic experiment (A, n=12) and in the prophylactic experiment (B, n=10). The results are expressed as means ± SD.
In the therapeutic experiment, similar body weight changes were observed in the initial 6 weeks for the ND control (ND+MCT) group and other groups, respectively. After alcohol treatment ended, the body weight gain in the alcohol withdrawal MCT control (HFD+Alcohol+MCT, alcohol was given for 6 weeks) group recovered to a higher level than the ND control group, and no significant differences were observed when comparing with Krill Oil treated groups. The body weight of the alcohol non-withdrawal MCT control (HFD+Alcohol+MCT, alcohol was given for 10 weeks) group increased with time during weeks 7-10 and Krill Oil treatments had no effect on body weight (Figure 2B).
Food consumption
In the prophylactic experiment, the ND control group had a consistent food consumption ranging from 22.2 to 27.7 g/rat/day, while the HFD control group had a decreased food consumption ranging from 17.8 to 21.2 g/rat/day. There was no difference between MCT control group and HFD control group in the initial 3 weeks while a marked decrease in HFD group was noted in the following 2 weeks with the lowest food consumption of 8.5 g/rat/day, and partially recovered in week 6. By comparison with MCT control group, no apparent changes in food consumption were noted in the Krill Oil treated groups except that relatively higher values were noted for 1000 mg•kg-1•d-1 Krill Oil group during D13 to D22 (Figure 3A).
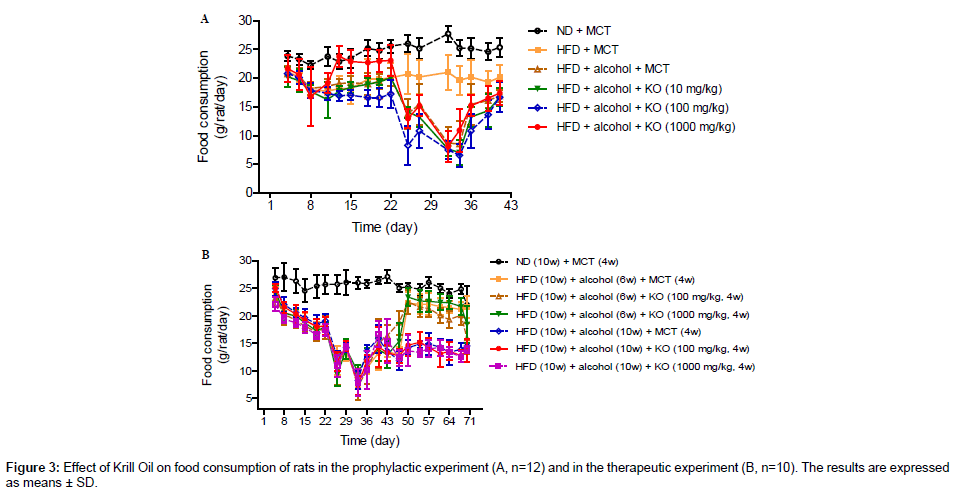
Figure 3: Effect of Krill Oil on food consumption of rats in the food-rats experiment (A, n=12) and in the therapeutic experiment (B, n=10). The results are expressed as means ± SD.
In the therapeutic experiment, similar food consumption were noted in the initial 6 weeks for the ND control group and other groups, respectively. The food consumption in the alcohol withdrawal MCT control group recovered in weeks 7 to 10 while no apparent changes were noted for the alcohol non-withdrawal MCT control group, and Krill Oil treatment had no effects on food consumption compared with the model control group (Figure 3B).
Liver and serum TG and TC
In the prophylactic experiment, in comparison with ND control group, the liver TG level elevated significantly in the HFD control group, and which elevated further after alcohol exposure for six weeks, indicating an accumulation of TG in liver tissue, a characteristic sign of fatty liver. By comparison with ND control group, an elevation of liver TC was also observed in HFD control group, while no apparent further elevation was noted after alcohol exposure. Krill Oil treatment inhibited HFD and alcohol induced elevations in liver TG and TC in a dose dependent manner (Figure 4A-a; Figure 4A-b).
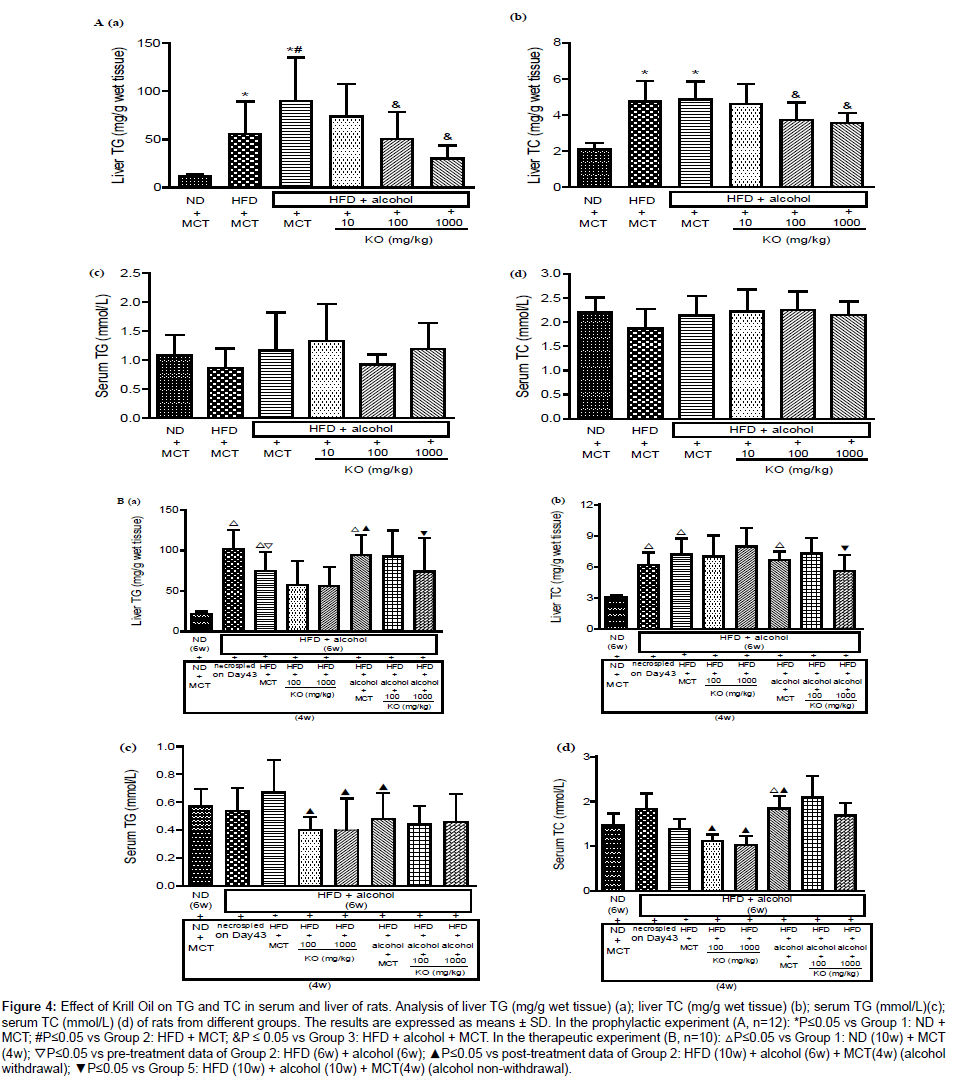
Figure 4: Effect of Krill Oil on TG and TC in serum and liver of rats. Analysis of liver TG (mg/g wet tissue) (a); liver TC (mg/g wet tissue) (b); serum TG (mmol/L)(c); serum TC (mmol/L) (d) of rats from different groups. The results are expressed as means ± SD. In the prophylactic experiment (A, n=12): *P≤0.05 vs Group 1: ND + MCT; #P≤0.05 vs Group 2: HFD + MCT; &P ≤ 0.05 vs Group 3: HFD + alcohol + MCT. In the therapeutic experiment (B, n=10): △P≤0.05 vs Group 1: ND (10w) + MCT (4w); ▽P≤0.05 vs pre-treatment data of Group 2: HFD (6w) + alcohol (6w); ▲P≤0.05 vs post-treatment data of Group 2: HFD (10w) + alcohol (6w) + MCT(4w) (alcohol withdrawal); ▼P≤0.05 vs Group 5: HFD (10w) + alcohol (10w) + MCT(4w) (alcohol non-withdrawal).
Unlike the changes of TG and TC noted in liver tissue, the serum TG and TC had no change either after exposure to alcohol or after treated with Krill Oil (Figure 4A-c; Figure 4A-d).
In the therapeutic experiment, after withdrawal from alcohol exposure, the liver TG showed a spontaneous recovery. Krill Oil treatment accelerated recovery independent of doses. For the groups that continued with alcohol gavage until the end of week 10, Krill Oil treatment decreased liver TG in a dose dependent manner (Figure 4Ba). After withdrawal from alcohol gavage, the liver TC showed a slight increase in comparison with pre-treatment data, while there was no changes in the Krill Oil treated groups. For the groups continuing with alcohol gavage until the end of week 10, no apparent change in the liver TC levels was observed in Krill Oil low-dose groups while a decrease was observed in the high-dose group (Figure 4B-b).
No change in serum TG was observed after six weeks of alcohol gavage, Serum TG was slightly increased after alcohol gavage withdrawal. By comparison with the MCT control group, Krill Oil treatment (4 weeks) decreased the serum TG independent of dose. No change in serum TG level was observed after 10 weeks of alcohol gavage, and Krill Oil treatments had no effect on serum TG. The serum TC had a slight increase after six weeks of alcohol gavage and recovered to normal value after alcohol gavage ended. Krill Oil treatments decreased serum TC. There was no further change in serum TC after ten weeks of alcohol gavage and Krill Oil treatments showed no effects on serum TC either. (Figure 4B-c; Figure 4B-d).
Liver weight and liver-to-body weight ratio
In the prophylactic experiment, by comparison with the ND control group and HFD control group, the liver to body weight ratio showed a significant increase after oral gavage of alcohol. The increased ratio was reversed by the concurrent intake of krill in a dose dependent manner (Figure 5A).
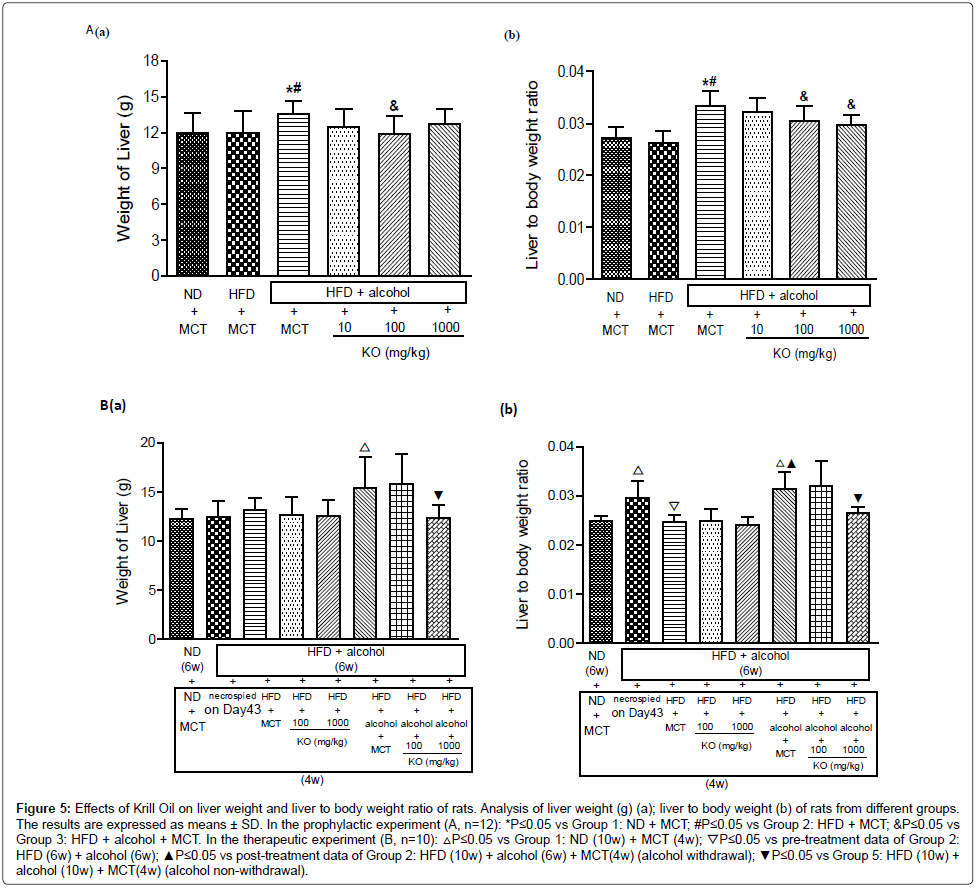
Figure 5: Effects of Krill Oil on liver weight and liver to body weight ratio of rats. Analysis of liver weight (g) (a); liver to body weight (b) of rats from different groups. The results are expressed as means ± SD. In the prophylactic experiment (A, n=12): *P≤0.05 vs Group 1: ND + MCT; #P≤0.05 vs Group 2: HFD + MCT; &P≤0.05 vs Group 3: HFD + alcohol + MCT. In the therapeutic experiment (B, n=10): △P≤0.05 vs Group 1: ND (10w) + MCT (4w); ▽P≤0.05 vs pre-treatment data of Group 2: HFD (6w) + alcohol (6w); ▲P≤0.05 vs post-treatment data of Group 2: HFD (10w) + alcohol (6w) + MCT(4w) (alcohol withdrawal); ▼P≤0.05 vs Group 5: HFD (10w) + alcohol (10w) + MCT(4w) (alcohol non-withdrawal).
In the therapeutic experiment, after withdrawal of alcohol gavage, the liver to body weight ratio returned to normal values and there was no difference between model control group and Krill Oil treatment groups, indicating a spontaneous recovery of fatty liver and no accelerating effect was noted in Krill Oil treated groups. However, the liver to body weight ratio remained at high level when the oral gavage of alcohol continued after initiation of Krill Oil treatment. The increased ratio was reversed by oral gavage of high dose of Krill Oil (Figure 5B).
Pathology
In the prophylactic experiment, after oral gavage of alcohol for six weeks, all of the vehicle control rats showed a yellowish discoloration of the liver at necropsy. The incidence of this discoloration decreased after Krill Oil treatment in a dose dependent manner.
Moderate (++) to marked (+++) cytoplasmic vacuolization in the MCT control group were noted, alcohol treatment led to higher severity of cytoplasmic vacuolization compared with normal diet and high-fat diet control groups, indicating a successful induction of alcoholic fatty liver. Concurrent treatments with Krill Oil decreased the frequency and severity of alcohol induced cytoplasmic vacuolization in a dosedependent manner (Figure 6A, 6C).
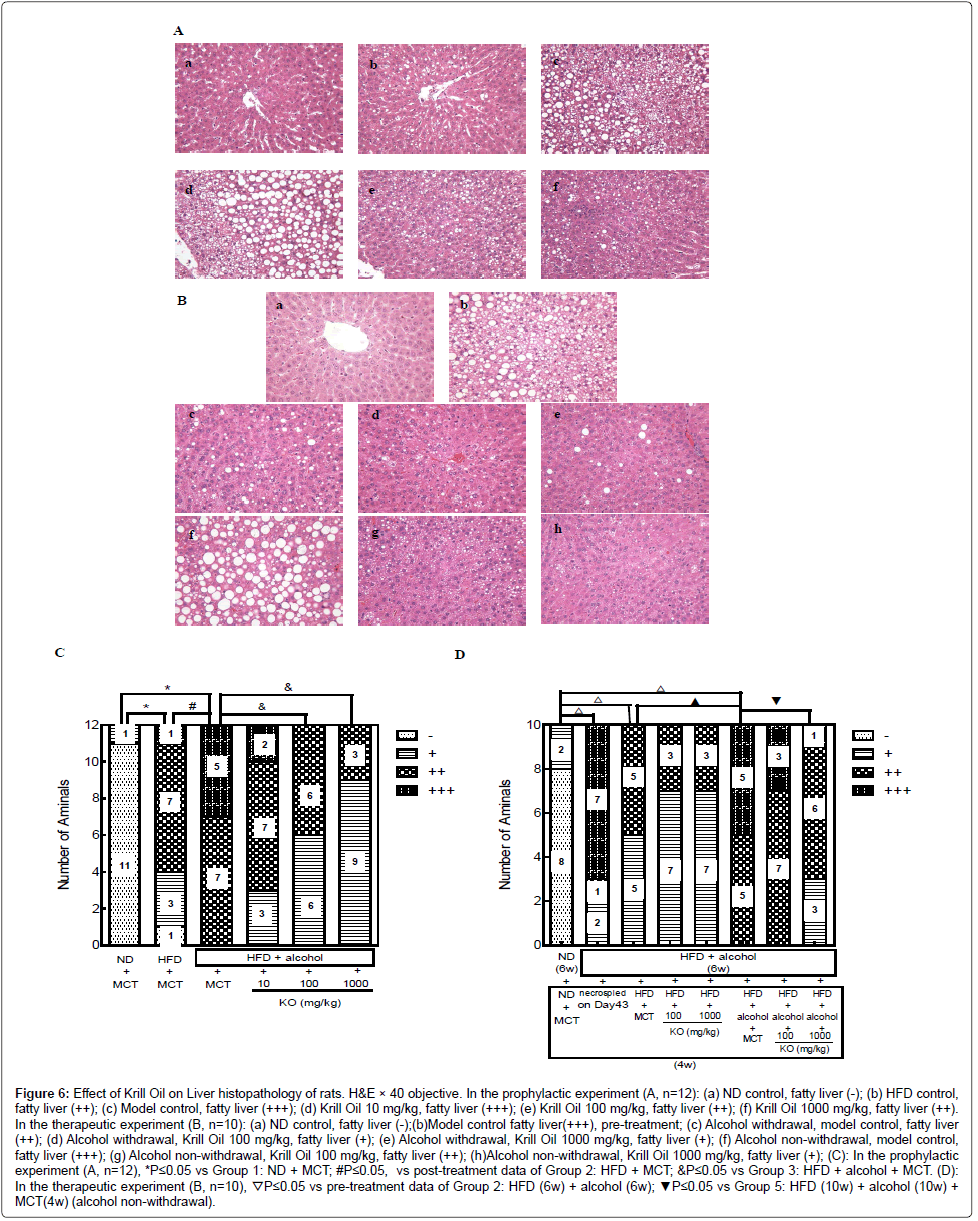
Figure 6: Effect of Krill Oil on Liver histopathology of rats. H&E × 40 objective. In the prophylactic experiment (A, n=12): (a) ND control, fatty liver (-); (b) HFD control, fatty liver (++); (c) Model control, fatty liver (+++); (d) Krill Oil 10 mg/kg, fatty liver (+++); (e) Krill Oil 100 mg/kg, fatty liver (++); (f) Krill Oil 1000 mg/kg, fatty liver (++). In the therapeutic experiment (B, n=10): (a) ND control, fatty liver (-);(b)Model control fatty liver(+++), pre-treatment; (c) Alcohol withdrawal, model control, fatty liver (++); (d) Alcohol withdrawal, Krill Oil 100 mg/kg, fatty liver (+); (e) Alcohol withdrawal, Krill Oil 1000 mg/kg, fatty liver (+); (f) Alcohol non-withdrawal, model control, fatty liver (+++); (g) Alcohol non-withdrawal, Krill Oil 100 mg/kg, fatty liver (++); (h)Alcohol non-withdrawal, Krill Oil 1000 mg/kg, fatty liver (+); (C): In the prophylactic experiment (A, n=12), *P≤0.05 vs Group 1: ND + MCT; #P≤0.05, vs post-treatment data of Group 2: HFD + MCT; &P≤0.05 vs Group 3: HFD + alcohol + MCT. (D): In the therapeutic experiment (B, n=10), ▽P≤0.05 vs pre-treatment data of Group 2: HFD (6w) + alcohol (6w); ▼P≤0.05 vs Group 5: HFD (10w) + alcohol (10w) + MCT(4w) (alcohol non-withdrawal).
In the therapeutic experiment, for the alcohol withdrawal model control group, 40% rats had a liver with yellowish or whitish discoloration. No apparent changes were noted in the Krill Oil treated groups. Eight of the 10 rats in alcohol non-withdrawal model control group had liver yellowish or whitish discoloration. After treated with high-dose Krill Oil the incidence decreased to 2/10. Low-dose Krill Oil had no effects on alcohol induced discoloration.
Alcohol withdrawal vehicle control animals had fatty liver (cytoplasmic vacuolization) with 50% slight and 50% moderate. Low dose Krill Oil and high dose Krill Oil treatment improved the ratio to 70% slight and 30% moderate. All the alcohol non-withdrawal model control animals had fatty liver (cytoplasmic vacuolization) with 50% moderate and 50% marked. Krill Oil treatment did not change the overall incidence of fatty liver with 10/10 rats in both groups. However the severity of fatty liver decreased in Krill Oil treated groups with 70% moderate and 30% marked found in the low-dose group, and 30% slight, 60% moderate and 10% marked in the high-dose group (Figure 6B, 6D).
Microarray analysis
A total of 19 genes were differentially expressed after Krill Oil treatment with q-value ≤5% and Fold Change ≥2 or ≤ 0.5 (Table 2). Of these 19 genes, 1 gene was up-regulated and the other 18 genes were down-regulated. However, None of the 19 genes were directly correlated with alcohol metabolism and lipid metabolism. There are another 105 genes whose Fold Changes were ≥2 or ≤0.5 while their q-values were >5%.
| K-Oil | F-oil | Soy-Lec | 1:1 Mix of F-O & S-L | ||
|---|---|---|---|---|---|
| Polar lipid (weight%) | 43.4 | — | 64.8 | 32.4 | |
| Nonpolar lipid (weight%) | 53.6 | 100 | 35.2 | 67.6 | |
| Fatty acid (%) | C14:0 | 11.9 | 5.2 | — | — |
| C16:0 | 21.6 | 7.3 | 29.1 | 17.4 | |
| C18:0 | 1.3 | 0.7 | 2.5 | 1.5 | |
| C18:1 | 17.7 | 8.6 | 12 | 10.3 | |
| C18:2 | 1.7 | 1.1 | 50.9 | 26 | |
| C18:3 | 1.2 | 0.8 | 5.5 | 3.2 | |
| C20:5 | 15.2 | 29.3 | — | 15 | |
| C22:6 | 7.7 | 13.1 | — | 6.7 | |
Table 1: Lipid and fatty acid composition.
| Gene ID | Gene Title | Gene Symbol | Fold Change | q-value (%) |
|---|---|---|---|---|
| 1384671_at | --- | --- | 2.23 | 0.00 |
| 1387242_at | eukaryotic translation initiation factor 2-alpha kinase 2 | Eif2ak2 | 0.49 | 0.00 |
| 1374337_at | --- | --- | 0.44 | 0.00 |
| 1391489_at | immunity-related GTPase family, M | Irgm | 0.43 | 0.00 |
| 1384180_at | interferon-induced protein with tetratricopeptide repeats 2 | Ifit2 | 0.43 | 0.00 |
| 1379568_at | interferon-induced protein with tetratricopeptide repeats 2 | Ifit2 | 0.41 | 0.00 |
| 1389034_at | ubiquitin specific peptidase 18 | Usp18 | 0.25 | 0.00 |
| 1382902_at | potential ubiquitin ligase | Herc6 | 0.21 | 0.00 |
| 1370913_at | radical S-adenosyl methionine domain containing 2 | Rsad2 | 0.21 | 0.00 |
| 1382314_at | interferon, alpha-inducible protein (clone IFI-15K) | G1p2 | 0.13 | 0.00 |
| 1387283_at | myxovirus (influenza virus) resistance 2 | Mx2 | 0.13 | 0.00 |
| 1371970_at | family with sequence similarity 111, member A | Fam111a | 0.03 | 0.00 |
| 1370314_at | solute carrier family 20 (phosphate transporter), member 1 | Slc20a1 | 0.50 | 2.69 |
| 1373197_at | similar to Protein KIAA1404 | LOC686701 | 0.47 | 3.66 |
| 1387354_at | signal transducer and activator of transcription 1 | Stat1 | 0.46 | 3.66 |
| 1393044_at | cytidine monophosphate (UMP-CMP) kinase 2, mitochondrial | Cmpk2 | 0.44 | 3.66 |
| 1383564_at | interferon regulatory factor 7 | Irf7 | 0.34 | 3.66 |
| 1371015_at | myxovirus (influenza virus) resistance 1 | Mx1 | 0.30 | 3.66 |
| 1383075_at | cyclin D1 | Ccnd1 | 0.34 | 4.65 |
Table 2: List of differentially expressed genes after Krill Oil treatment.
Many studies [22,23] indicate that high-fat diet is helpful to maintain relative high blood alcohol concentration and exacerbate to induce fatty liver disease in alcoholic rats. Thus in this study, we use alcohol gavage along with feeding a high-fat diet to induce alcoholic fatty liver in Sprague-Dawley rats. Our data showed that the liver tissue TG increased from 11.42 mg/g to 48.18 mg/g after feeding high fat diet for six weeks, and the value increased further to 89.65 mg/g after alcohol exposure for six weeks. The histopathology data also showed an increased incidence and severity of liver cytoplasmic vacuolization after alcohol exposure for six weeks, indicating that the alcoholic fatty liver model was induced successfully. Concurrent supplementation of Krill Oil for six weeks inhibited the increase in liver TG and the increases in incidence and severity of liver cytoplasmic vacuolization induced by alcohol exposure, indicating Krill Oil has a prophylactic effect on fatty liver formation in this rat model. When Krill Oil was supplemented, after alcohol exposure for six weeks, i.e. after alcoholic fatty liver was already induced, the increased fatty liver related parameters either had an accelerated recovery where alcohol exposure had ended at the end of week 6, or had an attenuation when alcohol exposure continued along with Krill Oil supplementation, indicating Krill Oil had a therapeutic effect on the alcoholic fatty liver in this rat model. To find out the potential mechanisms underlying Krill Oil’s effect on alcoholic fatty liver, we conducted a microarray analysis using Affymetrix GeneChip Rat Genome 230 2.0 Array. Unfortunately we did not find apparent changes at mRNA level that had a direct correlation with alcohol metabolism and lipid metabolism. Instead, the results showed most of the genes with marked changes were related to Interferon (IFN), a pluripotent cytokine in immune system modulation. The relationship between IFN and fatty liver formation has rarely been reported. Sbarbati’s study showed IFN treated suckling mice had liver lipid accumulation while there is no such lipid accumulation in the control mice [24]. Wallace’s data showed that PUFAs could inhibit production of IFN-γ in mice and Irons’ data showed that PUFAs could impair the IFN-γ responsiveness via diminished receptor signaling in mice [25,26]. As Krill Oil contains a high proportion of ω-3 PUFAs, its inhibitory effects on fatty liver may attribute to PUFAs’ inhibitory effect on IFN signaling and subsequently inhibited fatty liver formation. Our data indicated a probable involvement of IFN in the Krill Oil’s effects on fatty liver. More work needed to be done to verify this possible mechanism.
As Krill Oil contains a high proportion of ω-3 PUFAs bound to phospholipids and many kinds of antioxidants including Vitamin E, alltrans- retinol Vitamin A, canthaxanthin and astaxanthin, its inhibitory effect on alcoholic fatty liver can be attributed to PUFAs, phospholipids and astaxanthin, or any combination thereof. Song et al. showed that alcohol-DHA/AA-supplemented diet ameliorates the fatty liver induced by chronic alcohol administration in rats alcohol induced elevation in CYP2E1, nitric oxide synthase, nitrite and mitochondrial hydrogen peroxide as well as reduction in mitochondrial aldehyde dehydrogenase, ATP synthase, and 3-ketoacyl-CoA thiolase all returned to normal levels in rats fed with the alcohol-DHA/AA-supplemented diet [27].
Unlike fish oil, which is based on ω-3 fatty acids bound to triglycerides, Krill Oil’s ω-3 fatty acids are bound to phospholipids [16,17]. Therefore Krill Oil contains a high proportion of phosphatidylcholine (PC). It has been reported that soybean polyenylphosphatidylcholine (PPC), a mixture of 94%-96% of polyunsaturated phosphatidylcholine, could affect the parameters in fatty liver induced by alcohol consumption. The alleviated fatty liver in the PPC-treated rats is associated with normalizing mitochondrial oxidation of palmitoyl-1-carnitine, and activities of cytochrome oxidase, serum glutamate dehydrogenase and aminotransferases which are inhibited or stimulated by alcohol exposure [28]. Aleynik’s study suggests alcohol exposure induced S-adenosylmethionine (SAMe) depletion can be reversed by PPC, along with the prevention of the alcohol-induced oxidative stress. As PCs are produced in the liver via methylation of phosphatidylethanolamine by SAMe, PPC might decrease the utilization of SAMe by providing PCs, and subsequently correct GSH depletion and prevent the alcohol induced oxidative stress [29].
As for astaxanthin, Ikeuchi et al showed a reduction of high-fat diet feeding induced increases in liver weight, liver triglyceride, plasma triglyceride, and total cholesterol [30]. Bhuvaneswaria’s study suggests that astaxanthin supplementation promotes insulin sensitivity and prevented liver injury by decreasing CYP2E1, myeloperoxidase, and nitro-oxidative stress and by improving the antioxidant status in them [31].
We had conducted a study to compare the prophylactic effect of Krill Oil, fish oil, soy lecithin and 1:1 mix of fish oil and soy lecithin on alcohol induced fatty liver in rats. The results indicate that Krill Oil has a prophylactic effect at dose level of 100 mg/kg, while fish oil, soy lecithin and 1:1 mix of fish oil and soy lecithin have no effects at this dose. As showed in Table 1, Krill Oil is lower in PUFA than all others. In respect of the highly unsaturated fatty acids (HUFA), a subset of PUFA which have 4 or more double bonds, Krill Oil is lower than fish oil. Krill Oil contains less polar lipid, i.e. the fatty acids which have at least one double bond, than soy lecithin. What makes Krill Oil more effective? We think that it is probably one of the characteristics of Krill Oil, i.e., the bond between PUFA with phospholipids. The supporting evidence can be found in several recently published clinical and preclinical studies. Three clinical studies showed that EPA+DHA in Krill Oil is more easily and effectively absorbed after ingestion and subsequently distributed in the blood of human subjects when compared with those in fish oil [17,32, 33]. In a study in obese Zucker rats, Krill Oil led to a significantly higher incorporation of EPA and DHA into tissue PLs than fish oil [34]. In another rat study, DHA in the form of PL had a 2-fold higher accumulation in the brain than that in the form of TG [35]. A later study in baboon neonates showed a 2.1-fold preferential incorporation and retention of dietary PL-derived arachidonic acid (AA) over TG-derived AA in the brain [36].
Based on the published data, we may attribute Krill Oil’s outstanding protective effect on fatty liver to: 1) PUFAs’ protective effects; 2) PC’s protective effects; 3) astaxanthin’s protective effects; 4) astaxanthin’s antioxidative activity which makes PUFAs more stable and thus improves PUFAs’ protective effects on fatty liver; 5) the bonds between PUFAs and PLs which make PUFAs more easily absorbed and thus improve PUFAs’ protective effects on fatty liver.
Our results demonstrate that Krill Oil supplement not only has a potential prophylactic effect on alcoholic fatty liver formation, but also has a potential therapeutic effect on the already formed alcoholic fatty liver. The protective effects of Krill Oil can be attributed to its main components, i.e. ω-3polyunsaturated fatty acids, phosphatidylcholine, astaxanthin, or any combination thereof, which have been reported to have protective effects on fatty liver.
This work was supported by Nippon Suisan Kaisha, Ltd., Tokyo, Japan. The authors are solely responsible for the design and conduct of the study; collection, management, analysis, and interpretation of the data; as well as preparation of the manuscript. We would like to thank the participants who contributed their time to this project.
HLW, LH, MWG, YXS, CLZ and TT were involved in the study design, data analysis, interpretation, and manuscript writing. Most of the experimental procedures were carried out by employees at JOINN Laboratories, Suzhou. QL was involved in the preparation of the study. The study was mainly performed by QL, WJZ and HW, who were also involved in data analysis and manuscript writing. MWG was involved in coordination of the study conduct. All authors have read and approved the final manuscript.
LH is employed in Nippon Suisan Kaisha, Ltd. HLW, QL, WJZ, HW and MWG were employed at JOINN Laboratories when the study was performed.