Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2015) Volume 4, Issue 1
High-Cell-Density Cultivations show a huge potential to produce recombinant proteins to amounts greatly exceeding the availability in natural resources. An interesting example of a recombinant protein is an L-arabinose isomerase, which is able to convert D-galactose to the low-caloric and low-glycaemic bulk sweetener D-tagatose. Within this study, the L-arabinose isomerase from Geobacillus stearothermophilus was expressed intracellularly in Escherichia coli. The cultivation medium contained glucose, yeast extract and various macro- and micronutrients. The effect of air flow rate on E. coli growth and expression of L-arabinose isomerase was studied. After 52 hours, an Optical Density and Dry Cell Weight of 154 ± 4 and 54.8 ± 1.3 g L-1 were reached, respectively by regulating the air flow rate between 0.2 and 30 L min-1. A corresponding L-arabinose isomerase activity of 6.99 ± 0.46 U mL-1 was reached. A drawback of High-Cell-Density Cultivation is the production of the by-product acetic acid which may inhibit growth. However, the acetic acid concentration was maintained as low as possible during fermentation to avoid inhibitory effects inherent to this compound. With the L-arabinose isomerase produced, a conversion percentage of 37.1 ± 1.5% was achieved, corresponding to 94.9 ± 3.7 g L-1 D-tagatose. Thus, the implementation of a High-Cell- Density Cultivation led to an efficient expression of the L-arabinose isomerase enzyme and D-tagatose production. Also the storage stability of the cells was investigated during several months at 4°C. A stable L-arabinose isomerase enzyme was noticed during at least 8 months storage at 4°C.
Keywords: Escherichia coli, High-Cell-Density cultivation, L-Arabinose isomerase, D-tagatose, Alternative sweetener
L-AI: L-Arabinose Isomerase; HCDC: High- Cell-Density Cultivation; WTi: Wild Type enzyme produced with an inducible expression system; CIP: Center for Protein Engineering; IPTG: IsoPropyl β-D-1-ThioGalactopyranoside; Tris: Tris (hydroxymethyl) Aminomethane; OD: Optical Density; DCW: Dry Cell Weight; HPLC: High-Performance Liquid Chromatography; DAD: Diode Array Detector; RI: Refractive Index
Nowadays, the efficient production of recombinant proteins has become more and more important in biotechnology. The combination of recombinant proteins and large-scale processes make it possible to produce these proteins in amounts exceeding greatly the availability in natural resources [1-4]. The most commonly used host organism for recombinant protein production is still Escherichia coli because its genetic properties and physiological behaviour are well-known [1-3,5-9]. Within this study, High-Cell-Density Cultivation (HCDC) using E. coli as host organism was evaluated in view of a good expression and production of the L-arabinose isomerase (L-AI) from Geobacillus stearothermophilus. The L-AI enzyme converts D-galactose into D-tagatose. D-tagatose is a natural occurring bulk sweetener showing a wide range of healthy properties. The sweetness and aroma profile of D-tagatose are similar to sucrose. Furthermore, D-tagatose possesses a low caloric value (1.5 kcal g-1), a low glycaemic index, prebiotic properties and shows non-cariogenic properties [10-17]. D-tagatose is present in apples, pineapples, oranges, sterilized cow milk etc. but not in sufficient amounts for commercialization [12,13]. For industrial implementation, a biochemical production of D-tagatose by means of an L-AI enzyme is preferred. However, a cost-efficient production of the L-AI enzyme is therefore required.
An interesting approach for a cost-efficient production of recombinant proteins is High-Cell-Density Cultivation (HCDC) of the host organism. This method is especially interesting for E. coli, where recombinant proteins often accumulate intracellular [6]. In this case, the productivity is directly correlated with the final cell density [1]. The advantages of HCDC are its reduced reactor volume, enhanced downstream processing, higher volumetric productivities, reduced waste water, less production costs and reduced investment in equipment [1,2]. However, several disadvantages are also encountered during HCDC such as the high initial concentration of medium components and low oxygen solubility. Furthermore, under oxygen limiting conditions and a high carbon concentration, by-products like acetate can be produced which inhibit growth [2-3,6].
In literature, the cultivation of E. coli for the production of the L-AI with a complex medium [18]. The medium contained 3.0 g L-1 yeast extract, 3.0 g L-1 malt extract, 5.0 g L-1 peptone, 10.0 g L-1 glucose and 50 μgmL-1 ampicillin. The temperature, air flow rate, and agitation were maintained at 37°C, 1.5 L min-1 and 600 rpm, respectively. The cultivation resulted in an optical density (OD) and Dry Cell Weight (DCW) of 12.4 and 3.83 g L-1, respectively [18]. Fermentations with complex media are less reproducible because nutrients like malt extract and peptone may vary in composition and quality. In HCDC, mostly defined media are used so that nutrient concentrations are known and can be controlled during fermentation. The medium for a HCDC contained 25.0 g L-1 glucose, 13.3 g L-1 KH2PO4, 4.0 g L-1 (NH4)2HPO4, 1.2 g L-1 MgSO4·7H2O, 1.7 g L-1 citric acid, 8.4 mg L-1 EDTA and several trace elements. As trace elements, 2.5 mg L-1 CoCl2·6H2O, 15.0 mg L-1 MnCl2·4H2O, 1.5 mg L-1 CuCl2·2H2O, 3.0 mg L-1 H3BO3, 2.5 mg L-1 Na2MoO4·2H2O, 13.0 mg L-1 Zn(CH3COO)2·2H2O, 100.0 mg L-1 Fe(III) citrate, 4.5 mg L-1 thiamine HCl and 0.1 mL L-1 antifoam were added [6]. Fed-batch cultivations were carried out in a 5 L bioreactor at 28°C. Air flow rate, oxygen flow rate, and agitation were maintained at 2.5 L min-1, 0.1 L min-1 and 500 rpm, respectively. The feeding solution contained 795.0 g L-1 glucose, 20.0 g L-1 MgSO4·7H2O, 13.0 mg L-1 EDTA and several trace elements. As trace elements, 4.0 mg L-1 CoCl2·6H2O, 23.5 mg L-1 MnCl2·4H2O, 2.5 mg L-1 CuCl2·2H2O, 5.0 mg L-1 H3BO3, 4.0 mg L-1 Na2MoO4·2H2O, 16.0 mg L-1 Zn(CH3COO)2·2H2O and 40.0 mg L-1 Fe(III) were added. After 25 hours of fermentation, a DCW of 128 g L-1 was achieved [6]. Nevertheless, the application of semi-defined media can sometimes be necessary to boost product formation [1].
In this study, the HCDC of E. coli using a semi-defined media was investigated for the overexpression of the L-arabinose isomerase enzyme from Geobacillus stearothermophilus [6]. In literature, few studies deal with an efficient production of the L-AI enzyme for implementation in the biochemical production process of D-tagatose. A Wild Type enzyme produced with an inducible expression system (WTi) was used within this research. The WTi enzyme needs to be induced with IsoPropyl β-D-1-ThioGalactopyranoside (IPTG) to initiate L-AI production. Inducers are often applied in fermentation of genetically modified microorganisms. An inducible system for cloned genes is incorporated since maximum growth must be achieved during the first part of the fermentation, in order to reach a high cell density [7]. In this study, the effect of air flow rate on growth of E. coli and expression of L-AI enzyme was studied.
Strain
The L-AI enzyme from Geobacillus stearothermophilus was expressed intracellularly in E. coli BL21. The WTi clone was supplied by the Center for Protein Engineering (CIP, University of Liège, Belgium). The E. coli BL21 production strain was used as expression system together with the pCm470 plasmid where an inducible pTac promoter was incorporated. Induction was realized with IPTG.
Inoculum
20.0 μl frozen stock culture was grown in 3.00 mL medium containing 20.0 g L-1 glucose (Tereos Syral), 16.0 g L-1 tryptone (Oxoid), 10.0 g L-1 yeast extract (Organo Technie), 5.00 g L-1 NaCl (VWR), 0.197 g L-1 MnCl2·4H2O (Caldic) and 0.0125 g L-1 chloramphenicol (Sigma- Aldrich) for 6 hours at 30°C. The mixture was stirred at 200 rpm (IKAWerke). Afterwards, 3.00 mL was transferred into 250 mL cultivation medium and stirred at 200 rpm for 16 hours at 30°C. The cultivation medium consisted of 20.0 g L-1 glucose (Tereos Syral), 10.0 g L-1 yeast extract (Organo Technie), 15.0 g L-1 K2HPO4 (Caldic), 7.50 g L-1 KH2PO4 (Caldic), 3.00 g L-1 (NH4)2SO4 (Caldic), 2.20 g L-1 Na-citrate (Caldic), 1.50 g L-1 MgSO4·7H2O (Caldic), 0.197 g L-1 MnCl2·4H2O (Chem-Lab), 0.0125 g L-1 chloramphenicol (Sigma-Aldrich) and 2.00 mLL-1 trace solution. The trace solution consisted of 33.8 g L-1 Zn(CH3COO)2·2H2O (VWR), 14.1 g L-1 Na-EDTA·2H2O (VWR), 3.00 g L-1 H3BO3 (Alfa Aesar), 2.50 g L-1 CoCl2·6H2O (Alfa Aesar), 2.10 g L-1 N MoO4·2H2O (VWR), 1.50 g L-1 CuCl2·2H2O (VWR) and 0.197 g L-1 MnCl2·4H2O (Chem-Lab). Two precultures were grown in parallel and 320 mL was used to inoculate 7.00 L cultivation medium in the bioreactor.
High-cell density cultivation
The cultivation was performed in a 15 L bioreactor containing an initial fermentation volume of 7.00 L cultivation medium (BioFlo IV, New Brunswick Scientific co., Enfield, U.S.A.). 18 mL Antifoam (Brenntag) was added at the start of the fermentation. The pH was controlled at 7.0 with 24.5% NH3 (v/v) (Brenntag). When glucose was depleted, a concentrated feed solution was added to the bioreactor at a flow rate of 1.0 mL min-1. The feed consisted of 354 g L-1 glucose (Tereos Syral), 178 g L-1 yeast extract (Organo Technie), 1.00 g L-1 MgSO4·7H2O (Caldic), 0.394 g L-1 MnCl2·4H2O (Chem-Lab), 0.0250 g L-1 chloramphenicol (Sigma-Aldrich) and 1.00 mL L-1 trace solution. During cultivation in the bioreactor, the dissolved oxygen concentration was kept above 15% saturation through regulation of stirring speed between 300 and 700 rpm. The air flow rate was studied at 5 and 10 L min-1, respectively. In addition, a variable air flow rate between 0.2 and 30 L min-1 was studied. If the dissolved oxygen concentration dropped below 15% saturation at maximum stirring speed, the air flow rate was enhanced. The temperature at the start of the fermentation was set at 30°C. After induction with 0.5 mM IPTG, the temperature was lowered to 25°C. After 31 and 33 hours of fermentation, 0.5 mM IPTG was added during the fermentations with an air flow rate of 5 and 10 L min-1, respectively. During fermentation with a variable air flow rate between 0.2 and 30 L min-1, cells were induced with 0.5 mM IPTG at 28 hours. A second induction with 0.5 mM IPTG took place after 32 hours of fermentation. Throughout the cultivations, pH, temperature, dissolved oxygen concentration DO, stirring speed and air flow rate were continuously monitored and recorded. Fermentations were carried out in triplicates in order to assess the reproducibility and corresponding standard deviations were calculated.
Off-line data analysis
The OD was determined by measuring the absorbance of the cell solution at a wavelength of 600 nm with a Cary 100 Bio UV-Visible spectrophotometer from Varian after appropriate dilution.
The DCW was determined on 10.0 mL aliquots of the culture medium. Cell pellets were collected through centrifugation (Awel centrifugation) at 4000 rpm for 9 minutes and were afterwards resuspended in Reversed Osmosis water and centrifuged again at 4000 rpm for 9 minutes. Finally, cell pellets were dried overnight at 100°C.
An SDS-PAGE was performed on aliquots taken during the cultivation with a variable air flow rate with a mini-PROTEAN TGX Stain-free Precast Gel (Bio-Rad). Precision plus protein all bleu standards (Bio-Rad) were used with a molecular weight ranging from 10 to 250 kDa (10, 15, 20, 25, 37, 50, 75, 100, 150 and 250 kDa). The gel was read in a Gel Doc EZ imager (Bio-Rad). Aliquots taken during the cultivation with a variable air flow rate between 0.2 and 30 L min-1 were diluted 25 times and were subsequently loaded on the gel.
The acetic acid concentration during the course of fermentation was determined with High-Performance Liquid Chromatography (HPLC) with Diode Array Detector (DAD). The determination of the acetic acid concentration with HPLC was performed on an Aquasil 5 μm C18 column (250 × 4.6 mm). The column was eluted at 20°C with a mixture of acetonitrile and 0.05 M KH2PO4 at a flow of 1.25 mL min-1. The elution method was started with an eluent containing 99% KH2PO4 for 20 minutes. Afterwards, the KH2PO4 concentration was decreased towards 50% during 5 minutes. Next, isocratic elution was performed for 5 minutes followed by an increase towards 99% KH2PO4 during 5 minutes. Finally, isocratic elution was maintained with 99% KH2PO4 for 2 minutes.
Analysis of L-arabinose isomerase activity
The activity of the L-AI enzyme was determined by adding 0.500 ml cell solution to 1.500 ml D-galactose solution [350 g L-1 D-galactose (Inalco), 50.0 mM Tris(hydroxymethyl)aminomethane (Tris, Sigma- Aldrich), 1.00 mM MnCl2.4H2O (Chem-Lab)] of pH 7.0 [adjusted with 37% (w/w) HCl (VWR)]. Mixtures were incubated for 2 hours at 60°C in a water bath with shaking mechanism (Heto Birkerod Denmark). Then, the D-tagatose concentration was determined with HPLC with refractive index (RI) detection. The determination of the D-tagatose concentration with HPLC was performed on a Bio-Rad Aminex Carbohydrate HPX-87P column (300 x 7.8 mm). The -samples were pre-treated with 20 μl mL-1 Carrez I and II (VWR) to remove the proteins. The column was eluted at 80°C with demineralised water at a flow rate of 0.6 mL min-1.
Determination of conversion degree
In order to determine the conversion efficiency of D-galactose towards D-tagatose, also conversion experiments were performed within a timeframe of 24 hours. 10.0 mL cell solution (end of fermentation) was added to 30.0 mL D-galactose solution [350 g L-1 D-galactose, 50.0 mM Tris, 1.00 mM MnCl2.4H2O] of pH 7.0 [adjusted with 37% (w/w) HCl]. Mixtures were incubated for 24 hours at 60°C in a water bath with shaking mechanism (Heto Birkerod Denmark). Afterwards, D-galactose and D-tagatose concentrations were determined with HPLC with RI detection following the method which has been described for the determination of the L-AI activity. The conversion degree was defined as the D-tagatose concentration produced after a certain time interval divided by the initial D-galactose concentration.
Storage stability
Also the storage stability of the L-AI enzyme was determined. 5.00 ml aliquots of the cell solution were stored at 4°C. The L-AI activity was determined at pre-defined time intervals by adding 0.500 ml cell solution stored at 4°C to 1.500 mL D-galactose solution [350 g L-1 D-galactose, 50.0 mM Tris, 1.00 mM MnCl2.4H2O] of pH 7.0 [adjusted with 37% (w/w) HCl]. Mixtures were incubated for 2 hours at 60°C in a water bath with shaking mechanism (Heto Birkerod Denmark) and corresponding D-tagatose concentrations were determined with HPLC. The relative L-AI activity was defined as the L-AI activity at the pre-defined time interval divided by the initial L-AI activity.
High-cell-density cultivation of E. Coli
High-Cell-Density Cultivation of E. coli was used to study if an efficient expression of L-AI enzyme can be reached. As mentioned already in the introduction part, these fermentations are characterized by several advantages. A well-known drawback is however associated with the limited oxygen solubility. Within this context, the effect of the air flow rate on cell growth and L-AI expression was evaluated. The air flow rate was respectively set at 5 L min-1 and 10 Lmin-1. In addition, the effect of a variable air flow rate between 0.2 and 30 L min-1 was studied. The mean values of the Optical Densities (OD) and the Dry Cell Weight (DCW) with the corresponding standard deviations are shown in Figure 1.
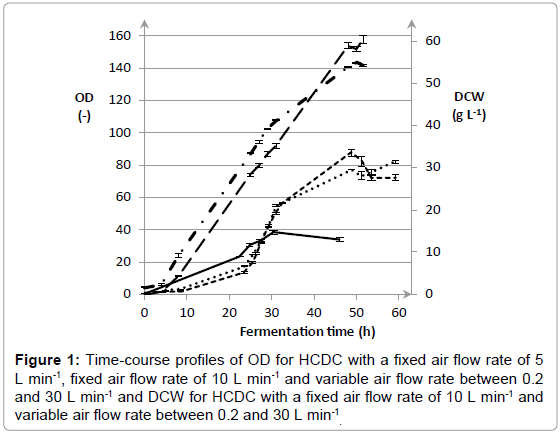
Figure 1: Time-course profiles of OD for HCDC with a fixed air flow rate of 5 L min-1, fixed air flow rate of 10 L min-1 and variable air flow rate between 0.2 and 30 L min-1 and DCW for HCDC with a fixed air flow rate of 10 L min-1 and variable air flow rate between 0.2 and 30 L min-1 .
The results in Figure 1 show that the air flow rate has a huge impact on growth of E coli. The highest OD and DCW was achieved with a variable air flow rate between 0.2 and 30 L min-1, namely 158 ± 2.3 and 54.1 ± 0.2 g L-1, respectively. An air flow rate of 10 L min-1 led to a decrease in OD and DCW towards 72.4 ± 2.2 and 31.4 ± 0.4 g L-1, respectively. A further decline of the air flow rate towards 5 L min-1 only resulted in an OD of 34.0 ± 1.0. Additionally, the effect of air flow rate on the L-AI activity was determined and corresponding results are included in Figure 2. For a fixed air flow rate of 5 L min-1, the L-AI activity was determined at the end of the fermentation. However, no L-AI activity was found. The highest expression of L-AI enzyme was achieved with a variable air flow rate between 0.2 and 30 L min-1, namely 7.37 ± 0.04 UmL-1. Decreasing the air flow rate to 10 L min-1 led to a decline in L-AI activity towards 5.04 ± 0.03 UmL-1. The results clearly illustrate that the air flow rate has a significant influence on the growth of E coli and on the expression of the L-AI enzyme. The best results were achieved with a variable air flow rate between 0.2 and 30 L min-1. In order to study the reproducibility of the fermentation procedure, the fermentation with a variable air flow rate was repeated three times. The time-course profiles of OD and DCW for cultivation of E. coli WTi with variable air flow rate between 0.2 and 30 L min-1 as well as the standard deviations calculated on three separate fermentations are shown in Figure 3. Only small variations in the time-course profiles were noticed for OD and DCW for the three cultivations which clearly demonstrate the reproducibility of the fermentation procedure. After 52 hours of fermentation, an OD and DCW were reached of 154 ± 4 and 54.8 ± 1.2 g L-1, respectively. Also, the mean L-AI activities and corresponding standard deviations were determined which are shown in Figure 4.
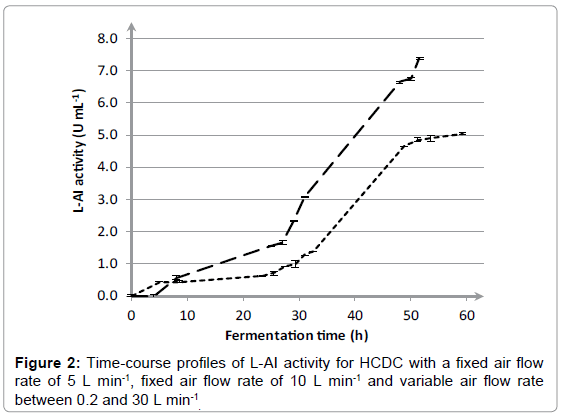
Figure 2: Time-course profiles of L-AI activity for HCDC with a fixed air flow rate of 5 L min-1, fixed air flow rate of 10 L min-1 and variable air flow rate between 0.2 and 30 L min-1 .
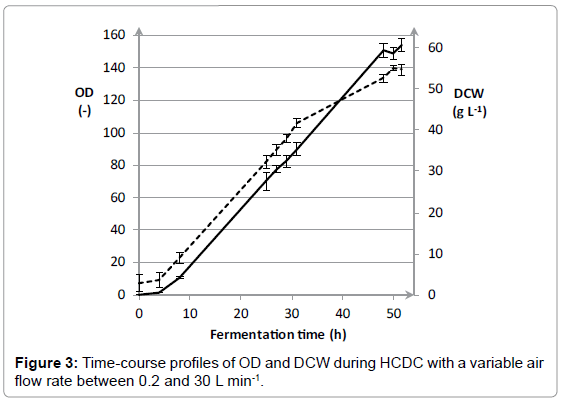
Figure 3: Time-course profiles of OD and DCW during HCDC with a variable air flow rate between 0.2 and 30 L min-1.
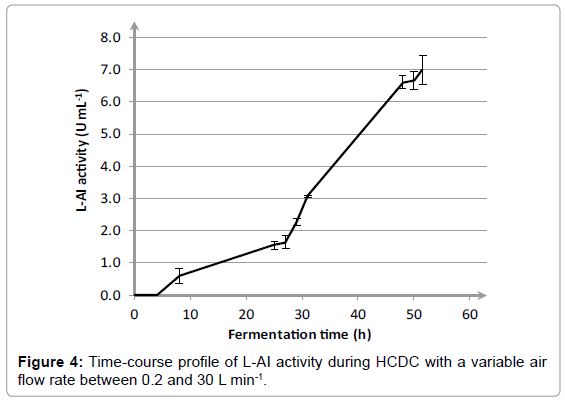
Figure 4: Time-course profile of L-AI activity during HCDC with a variable air flow rate between 0.2 and 30 L min-1.
Results in Figure 4 show that limited enzyme expression was already present before induction. At 27 hours, an L-AI activity of 1.63 ± 0.20 U mL-1 was noticed. At a high cell density, the cloned gene can be activated by addition of IPTG. After 28 hours, enzyme induction was initiated by adding 0.5 mM IPTG and at 32 hours, an additional induction with IPTG took place. After induction, L-AI expression enhanced markedly. An L-AI activity after 52 hours of 6.99 ± 0.46 U mL-1 was achieved. Corresponding standard deviations also imply that a reproducible expression of L-AI enzyme is realized which is important in view of further valorization activities. Expression of the L-AI enzyme was verified with an SDS-Page of aliquots taken during cultivation of E coli, as shown in Figure 5. The Geobacillus stearothermophilus L-AI enzyme was found around 50 kDa. Lane 2, 3 and 4 illustrate that only minor enzyme expression was present before induction with IPTG. After the first induction at 28 hours, a small increase was noticed in L-AI expression. The samples after 48, 50 and 52 hours, show a substantial increase in formation of L-AI enzyme corresponding to the high L-AI activities which are represented in Figure 4.
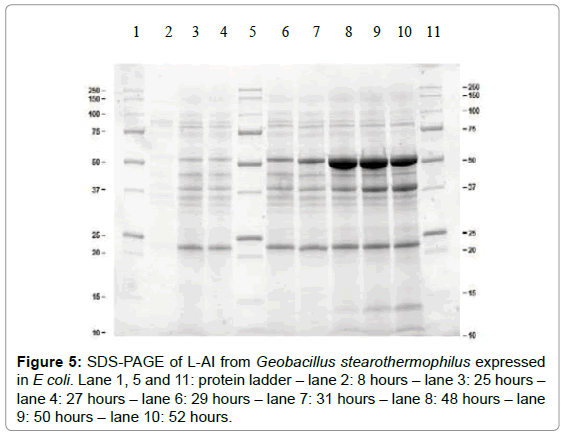
Figure 5: SDS-PAGE of L-AI from Geobacillus stearothermophilus expressed in E coli. Lane 1, 5 and 11: protein ladder – lane 2: 8 hours – lane 3: 25 hours – lane 4: 27 hours – lane 6: 29 hours – lane 7: 31 hours – lane 8: 48 hours – lane 9: 50 hours – lane 10: 52 hours.
A major drawback of HCDC of E coli is the production of acetate which can inhibit growth. When the carbon flux exceeds the capacity of the Kreb cycle (Crabtree effect), acetic acid is produced. Accumulation of acetic acid depends on the strain and on the medium composition. E. coli BL21 is a low acetate producer compared to E. coli K12 strain [19]. During fermentation, the accumulation of glucose was avoided by the choice of the feed flow rate in order to prevent the formation of acetic acid. Furthermore, from literature it is known that the presence of yeast extract in the feeding solution helps in achieving high cell density growth with little secretion of acetic acid. Moreover, yeast extract helps in decreasing the inhibitory effect of acetic acid [4,20]. During the HCDC of E. coli WTi with a variable air flow rate between 0.2 and 30 L min-1, the acetate concentration remained below 4 g L-1 taking in account the standard deviations.
Conversion degree
Next to the determination of the L-AI activity, also the conversion efficiency for the isomerization of D-galactose was investigated with cell solution obtained at the end of fermentation. The conversion degree was determined by determining the D-tagatose production during 24 hours. Figure 6 shows the conversion percentage as well as the corresponding D-tagatose concentration as a function of incubation time. During the first 2 hours, a strong increase was noticed in conversion percentage. Afterwards, conversion efficiency increased gradually, leading to a final conversion degree of 37.1 ± 1.5% after 24 hours. A conversion percentage of 37.1 ± 1.5% corresponds to a D-tagatose concentration of 94.9 ± 3.7 g L-1. A high conversion degree is required for implementation of a biochemical production process of D-tagatose on industrial scale. These results show however that the L-arabinose isomerase can be efficiently overexpressed in E. coli and that a high conversion degree of 37.1 ± 1.5% can be reached within a limited timeframe of 24 hours.
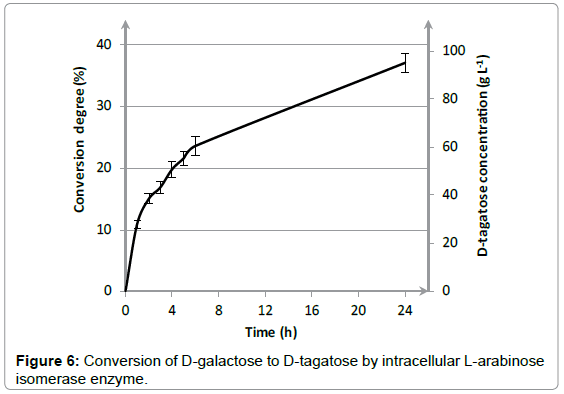
Figure 6: Conversion of D-galactose to D-tagatose by intracellular L-arabinose isomerase enzyme.
Storage stability
The storage stability of the cell solution containing the intracellular L-AI enzyme at 4°C was investigated during a period of 8 months. Figure 7 shows the relative L-AI activity as a function of storage time. During the first 2 months, a small increase in relative L-AI activity was noticed from 100 ± 8 to 110 ± 1%. Afterwards, the relative L-AI activity remained stable from 2 to 8 months. A high storage stability is an interesting feature for industrial application since storage of an enzyme batch is allowed during a longer period without compromising the activity of the L-AI enzyme.
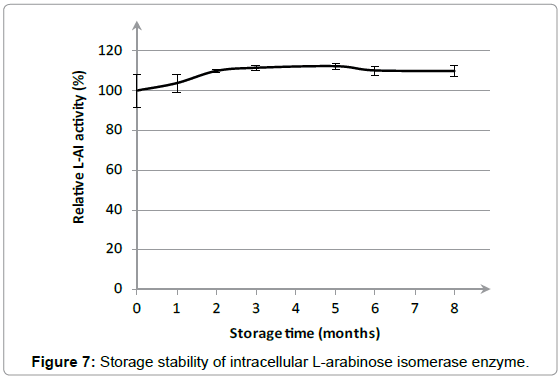
Figure 7: Storage stability of intracellular L-arabinose isomerase enzyme.
The L-arabinose isomerase enzyme can isomerise D-galactose to produce the high value-added sugar replacer, namely D-tagatose. As a low-caloric and low-glycaemic bulk sweetener, D-tagatose shows many applications in healthy food and nutrition for diabetics. However, a feasible enzymatic production process for D-tagatose is necessary to respond to the increasing demand for D-tagatose. Within this study, an efficient overexpression of the L-arabinose isomerase enzyme could be reached by means of HCDC of the host organism E. coli. The high DCW of 54.8 ± 1.2 achieved by HCDC by means of regulating the stirring speed and air flow rate resulted in a high productivity of the L-AI enzyme. During conversion, a conversion degree of 37.1 ± 1.5% was realized after 24 hours which corresponds to a D-tagatose concentration of 94.9 ± 3.7 g L-1. Future work will focus on the purification and immobilization of the L-AI enzyme in order to reach a stable and robust D-tagatose production process. Conversion efficiencies of immobilized cells will be compared and evaluated with the immobilized preparations of the crude extract and purified enzyme in order to determine the most efficient and economically attractive isomerization approach for biochemical production of the promising alternative sweetener, D-tagatose.
Author Monika Van Holsbeeck is supported by a PhD grant from KU Leuven (Belgium). Author Jan Van Impe holds the chair Safety Engineering sponsored by the Belgian Chemistry and Life Sciences Federation essenscia. The research was supported by project PFV/10/002 (Center-of-Excellence OPTEC - Optimization in Engineering) of the KU Leuven Research Council, Projects FWO-1518913N & FWO-G.0930.13 of the Research Foundation Flanders, and Interuniversity Attraction Pole IAP VII/19 (DYSCO - Dynamical Systems, Control and Optimization) of the Belgian Science Policy Office. Additional funding was realized through BAC resources from KU Leuven, Technology Campus Ghent. The authors would also like to thank CIP from the University of Liège (Belgium) for kindly supplying the strain.