Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 1
The southern- central Tyrrhenian Sea is a poorly studied area of the Mediterranean Sea, although its importance for the biological fluxes among the western and eastern basins is recognised. This note aimed at evaluating in the waters of the Southern Tyrrhenian Sea the efficiency of some steps of the organic matter processing (enzymatic hydrolysis, prokaryotic production and oxidation) and the role played by the prokaryotic community in this context. During the MEDBIO multidisciplinary cruise performed in July 2005, the distribution of prokaryotic abundance and activities (exo enzymatic activity, heterotrophic production and respiratory activity) was studied. The results showed the presence of high exo enzymatic activity rates (mostly leucine amino peptidase) in the deep sea, while heterotrophic production and respiratory activity rates were high at surface. Some speculations on the different behaviors of the Carbon fluxes occurring among the different epi, meso and bathypelagic layers are reported. At the epipelagic layer both the decomposition and biomass production processes were high. At the mesopelagic layer the organic matter was quickly mobilised but not efficiently incorporated into biomass, therefore the dissolved monomers remained available in the water column for export to other trophic levels or utilization in other chemical processes (lateral advection, oxidation). The bathypelagic layer was characterised by high levels of hydrolytic activity coupled with relatively high prokaryotic production; in this layer, exo enzymatic activity levels fall in an order of magnitude similar to that observed at the epipelagic one. This finding highlights the importance of microbially mediated processes in sustaining life in the marine depths.
<Keywords: Prokaryotic community; Exoenzymatic activities; Heterotrophic prokaryotic production; Respiration; Prokaryotic abundance; Carbon cycle; Tyrrhenian Sea
Prokaryotes (including Bacteria and Archaea) are an important component of the planktonic community of aquatic ecosystems and a critical link in the microbial loop, playing a vital role in nutrient cycling and food-web structure within the marine environment [1-3]. Particularly, heterotrophic microorganisms (bacteria, fungi) are the main consumers of Dissolved Organic Matter (DOM) and therefore they are the major players in organic matter turnover within the “microbial loop” [4]. Thanks to their wide biodiversity and their large variety of metabolic activities, microorganisms contribute significantly to the regulation of the flux of organic matter and energy from the biotic to abiotic compartments, acting in both decomposition (by DOM hydrolysis and nutrient release) and production processes (new living biomass production). Enzymatic hydrolysis of organic matter, which is mostly composed of both dissolved and particulate polymers, is recognized as a critical process and a “bottleneck” for organic matter utilization. In fact, heterotrophic microorganisms may uptake directly only low molecular weight substrates, such as aminoacids, simple sugars, fatty acids [5]. Many microorganisms, such as heterotrophic bacteria, microalgae, cyanobacteria, fungi and yeasts, are able to synthesize enzymes. Therefore, the determination of enzymatic hydrolysis rates allows to quantify the contribution of the microbial community to organic matter turnover [6-10].
Monomers released by enzymatic hydrolysis can be up taken and incorporated in new prokaryotic biomass (Heterotrophic Prokaryotic Production, HPP) with production of Particulate Organic Matter (POM). Oxidation of organic matter by Respiration (R) completes the mineralization process, releasing Carbon dioxide.
The importance of microbial food webs in marine environments and the crucial role of the interactions organic matter-heterotrophs in the biogeochemical cycles have been recognised by several studies [2,4,11]. Biogeochemical processes are mediated to a large extent by microorganisms [4,12], therefore the knowledge of their activity on organic polymers is needed to assess the role of microbes in organic matter transformation [13-15]. The study of microbial metabolism is one of the most fascinating topics in aquatic microbial ecology; the patterns of microbial metabolism and their regulation in response to biotic and abiotic factors have been described in ocean sites and different coastal areas of temperate, tropical and polar zones [5,13].
Nevertheless, in marine environments, knowledge of the microbial metabolism and of its modulation by environmental variables is still fragmentary [16]. Studies on prokaryotic activity and metabolism have firstly regarded the epipelagic waters [5,17-19]. Conversely, current research in microbial ecology has recognized that the variability of microbial processes across the epi-, meso- and bathypelagic zones represents an emerging topic [20-28].
Information on the deep sea is very scanty and predominantly collected in the open ocean [21-24,29,30]; comparatively few contributions are available for the Mediterranean Sea [6,9,26-28]. Focusing on the exoenzymatic activities, the occurrence of a wide temporal and spatial variability, with high activity rates also at bathypelagic depths, has been highlighted, and the significance of these measurements in the quantification of Carbon fluxes along the water column has been suggested [8,9,26,31-35]. The Mediterranean Sea is considered as an ecosystem particularly suitable for biogeochemical studies due to its particular characteristics of a semi-closed basin, with short residence times of water masses and the occurrence of deep water formation phenomena. Moreover, particular attention towards the biogeochemistry of this basin has been addressed in recent years [36-37], because of its vulnerability to climate changes [38].
The Tyrrhenian Sea is also characterized by the occurrence of anticyclonic gyres, which act as productive hot spots, showing nutrient rich waters at their margins. Gasparini, et al. [41] have also documented the spreading in the Tyrrhenian basin of the Eastern Mediterranean climate Transient (EMT), following the entrance of Aegean dense waters, which has resulted in temperature and salinity increases in the middle and deep Tyrrhenian waters.
An investigation was undertaken in July 2005 within the FIRBMIUR Project: “Biodiversity and organization of the communities in different environmental contexts,” to improve knowledge on the dynamics of the prokaryotic community inhabiting a Tyrrhenian area extending from the Aeolian Islands to the Gulf of Naples (centralsouthern Tyrrhenian Sea). The Tyrrhenian Sea has been considered as the most oligotrophic area of the western Mediterranean Sea in terms of nutrient concentrations and primary productivity [35].
The rationale for the present study was to assess whether there were different patterns of prokaryotic activity and abundance along the water column. Since surface and deeper zones are characterised by the relative predominance of different ecological processes - namely production in the first and decomposition in the second ones as well as in microbial loops between surface and deep oceans [25], we expected that the differences between the two zones were evident also at the level of the prokaryotic community, which is the major driver of aquatic ecosystems functioning.
Hydrological properties of the basin and sampling
Three main depth intervals have been considered in this study, in order to search for physical, chemical and microbial variations occurring along the water column: epipelagic zone (0
This separation roughly reflects the localization of the main water masses in the Tyrrhenian basin, which is a three-layer system. Its top layer (0-200 m) is characterized by the presence of the Modified Atlantic Water (MAW); potential temperature (θ) is greater than 14°C (with a maximum of 28°C) and Salinity (S) spans between 37.4 and 38.4. This layer, called summer-Atlantic Water (s-AW), occupies the upper 20 m and is the result of the thermal modification of the AW during the summer heating. The AW extends its influence below this warm and relatively salty upper layer down to 100-150 m of the water column [45]. The intermediate layer (200-700 m) is occupied by intermediated waters, among which the Levantine Intermediate Water (LIW, LIW (θ ≈ 13.5 - 13.9°C, S ≈ 38.45 – 38.75, σ ≈ 29.00 – 29.10 kg m-3) entering the Tyrrhenian Sea along the Sicily slope and leaving it mainly along the Sardinia side of the Sardinia-Sicily section. The deep layer is occupied by the Tyrrhenian Deep Water (TDW), a product of the mixing between LIW and Western Mediterranean Deep Water (WMDW), positioned below 1000 m depth which leaves the basin through the southern opening [46].
During the MEDBIO oceanographic cruise performed by R/V Universitatis (CONISMA) in the Southern-Central Tyrrhenian Sea in July 2005, water samples were collected from five stations located along a West-East transect (Figure 1), at different depths (5, 25, 50, 75, 100, 200, 500, 1500, 2500, 3000 m and near the bottom (10 m above the seafloor) using a rosette sampler equipped with 10-litres Niskin bottles. The seafloor depth varied in relation with the maximum depths of the different stations: stations D and 1 had maximum depths of 1059 and 2412 m, respectively; stations 2 and Vector were deeper (3364 and 3437 m, respectively). The deepest station was station 5 (3483 m).
Figure 1: Map of the sampling sites in the Tyrrhenian Sea. The arrows in the small picture indicate the direction of the circulation within the basin.
Physico-chemical measurements
Temperature (T), Salinity (S) and Dissolved Oxygen (O2) measurements were performed using a CTDO probe 911plus SeaBird profiler. Chlorophyll-a measurements were performed on discrete samples (1.5 – 2.0 litres) filtered on Whatman GF/F glassfibre filter. Filters were stored at -20°C, extracted in a 90% acetone solution, and measured with a spectrofluorometer (Varian Cary Eclipse) at 429 nm and 669 nm excitation and emission wavelengths before and after acidification [47]; calibration was performed with a standard of Chlorophyll-a from Anacystis nidulans (Sigma Co). Chl-a concentration was calculated according to Lorenzen [48].
For the determination of nutrient concentrations - ammonia (NH4), nitrate (NO3) and orthophosphate (PO4) contents - water samples were filtered through Whatman GF/F glass-fibre filters and kept frozen (-20°C). NH4 concentration was estimated by indophenol method [49]; NO3 was estimated by reduction in Cd-Cu column followed by diazotation, PO4 concentration was determined by reaction with ascorbic acid (Strickland and Parsons [50]).
Total Suspended Matter (TSM) was evaluated by a gravimetric method using a Mettler AT261 electronic microbalance (accuracy ± 1 microgram). Particle material was collected by filtering variable volumes of water on precombusted (480°C for 4 h) pre-weighted glass fibre filters (Whatman GF/F), which was then oven-dried at 60°C for 24 h [51].
Particulate Organic Carbon (POC) was determined on seawater samples ranging from 1.5 to 2 litres, filtered through pre-combusted 25 mm Whatman GF/F glass fibre filters (0.75 µm nominal porosity). After filtration, the filters (stored at-20°C until analysis) were exposed to HCl fumes to eliminate the inorganic carbon, dried at 60°C for 24 hours and then analyzed with a Perkin Elmer CHN Elemental Analyzer, utilizing Acetanilide as standard [52].
Prokaryotic metabolism and abundance measurements
The potential rates of exoenzymatic activities (leucineaminopeptidase, LAP; beta-glucosidase, b-GLU; alkaline phosphatase, AP) involved in the degradation of proteins, polysaccharides and organic phosphates, respectively, were measured according to Hoppe′s [53] method using the fluorogenic compounds L-leucine- 4-methylcoumarinyl-7-amide, 4-methylumbelliferyl-b-Dglucopyranoside and 4-methylumbelliferyl phosphate (Sigma-Aldrich, Co) as substrates for LAP, b-GLU and AP, respectively. Substrates were added at increasing concentrations (ranging from 0.2 to 4 mM final concentrations) to triplicate sub-samples and blanks, incubated, in the dark, for 2-3 hours at “in situ” ±1°C temperature The increase in fluorescence intensity, measured by a spectrofluorometer, was converted into the amount of substrate potentially hydrolysed. The maximum velocity of hydrolysis (Vmax) was calculated for each sample after linear (Lineweaver- Burk) transformation of the Michaelis- Menten equation. The Michaelis-Menten constant (Km= 1/2 Vmax) as an index of the affinity or strength of binding between the enzyme and its substrate, was also calculated using the same linear transformation.
Heterotrophic Prokaryotic Production (HPP) [54] was estimated from the rate of [3H] leucine incorporation using the micro centrifugation method according to Smith & Azam [55], using a conversion factor of 3.1 Kg C mol-1 [56]. Triplicate 1.0-ml samples and two blanks were incubated in the dark, at in situ ±1°C temperatures with l-[4,53H] leucine (Amersham Biosciences UK Limited) (final concentration: 25 nM).
Potential Respiratory Activity (R) rates of the entire microbial community (phytoplankton, microzooplankton and prokaryotes) were derived from the Electron Transport System (ETS) assay based on the technique of the tetrazolium reduction into INT-formazan according to Packard & Williams [57], as modified by Packard, et al. [58] and La Ferla, et al. [59]. Water samples were prefiltered on a 200 µm pore size mesh, in order to remove large particles and study the microplankton assemblage (0.5-200 µm sized). ETS assay values measured the maximum velocity (Vmax) of the dehydrogenases transferring electrons from their substrates (NADH, NADPH and succinate) to a terminal acceptor (O2). ETS values were converted into metabolic production of CO2 (Carbon Dioxide Production Rate, CDPR, in Carbon units) applying the Takahashi oxygen/carbon molar ratio [60] and conversion factors as usually adopted in oceanographic studies [9,61]. The results are reported as Vmax; a respiratory quotient of 1 was applied.
Total Prokaryotic Abundance (PA) was determined by epifluorescence microscopy on formalin fixed samples (2% final concentration) stained with 2, 3- diamidino phenylindole (DAPI) [62]. An epifluorescence AXIOPLAN2 Imaging microscope (Zeiss) equipped with the Axiocam digital camera and the AXIOVISION 3.1 software were used. To obtain adequate estimates, at least two hundred cells per each sample were counted.
Conversion factors adopted for the prokaryotic activities and abundances
LAP and b-GLU activity rates (expressed in nanomoles of leucine and b-glucoside released per hour) were converted into the amounts of Carbon potentially released from protein and carbohydrate hydrolysis, respectively, considering a number of 72 C atoms of leucine and b-glucoside. AP was expressed in terms of nanomoles of P potentially released from organic phosphates per hour and converted into nanograms of P by multiplying by a factor of 31.
Prokaryotic Carbon Demand (PCD) and Bacterial Growth Efficiency (BGE) calculations
The Prokaryotic Carbon Demand (PCD), namely the amount of Carbon required by the community to sustain its metabolism, was calculated, per each layer, as the sum HPP+R in terms of Carbon from the CO2 produced by R.
The Bacterial Growth Efficiency (BGE), indicating the fraction of Carbon utilized by bacteria for their growth, was also calculated as the percentage ratio between HPP and PCD [63]. This ratio was calculated assuming that in the euphotic zone (0-100 m) both the autotrophic and heterotrophic components contribute equally to the community respiration (R=50%), while in the aphotic zone R is completely heterotrophic (R=100%).
Data elaboration and statistical analysis
To allow the spatial comparison among the stations and epi-, meso and bathypelagic layers, the prokaryotic activity and abundances values were integrated using a trapezoidal method and normalised to the difference between the deepest and the shallowest depth of each layer. To determine the relationships among exoenzymatic values and prokaryotic and environmental parameters, the Pearson correlation coefficients were calculated. Analysis of Variance (ANOVA) was performed on data after their test for homogeneity of variances (Cochran’s test); a log transformation was performed when data did not satisfy the assumption of normality. Post-hoc comparisons among layers were performed by Student Newman Keuls test. All these analyses were performed using the SigmaStat software version 3.0.
Analysis of Similarities (ANOSIM) was carried out in order to assess statistical differences between clusters into the multivariate data sets. ANOSIM is a non-parametric permutation procedure that tests whether differences in dissimilarity between groups exceed differences within groups [64]. ANOSIM results produce a sample statistic, R, which represents the degree of separation between test groups: a value close to 1 indicates that all replicates of a sampling site or a treatment are each other more similar than any replicates of another site or treatment, whereas a value of 0 indicates no difference. The contribution of each parameter to the dissimilarity among clusters, as well as to the similarity within each cluster, was identified using SIMPER analysis [65].
The Multi Dimensional Scaling (MDS) analysis performed on normalized exoenzymatic activity rates evaluated the degree of similarity among the samples. The strength of the relationships among the three zones was calculated from similarity coefficients yielded by Cluster analysis.
The ANOSIM, SIMPER and MDS analyses were performed on normalized data using the PRIMER 6 software, version 6 B Plymouth Marine Laboratory, United Kingdom).
Physical and chemical measurements
The patterns of physical, chemical and trophic characteristics measured along the examined transect are reported in the contour plots of Figure 2. Temperature, O2 and TSM decreased with depth; Salinity reached its maximum at the mesopelagic layer, while decreased in the layer below. All the nutrient concentrations increased towards the bottom. Chl-a and POC, determined within the surface-200 m depth layer, showed quite low values, with a leap in correspondence with the pycnocline.
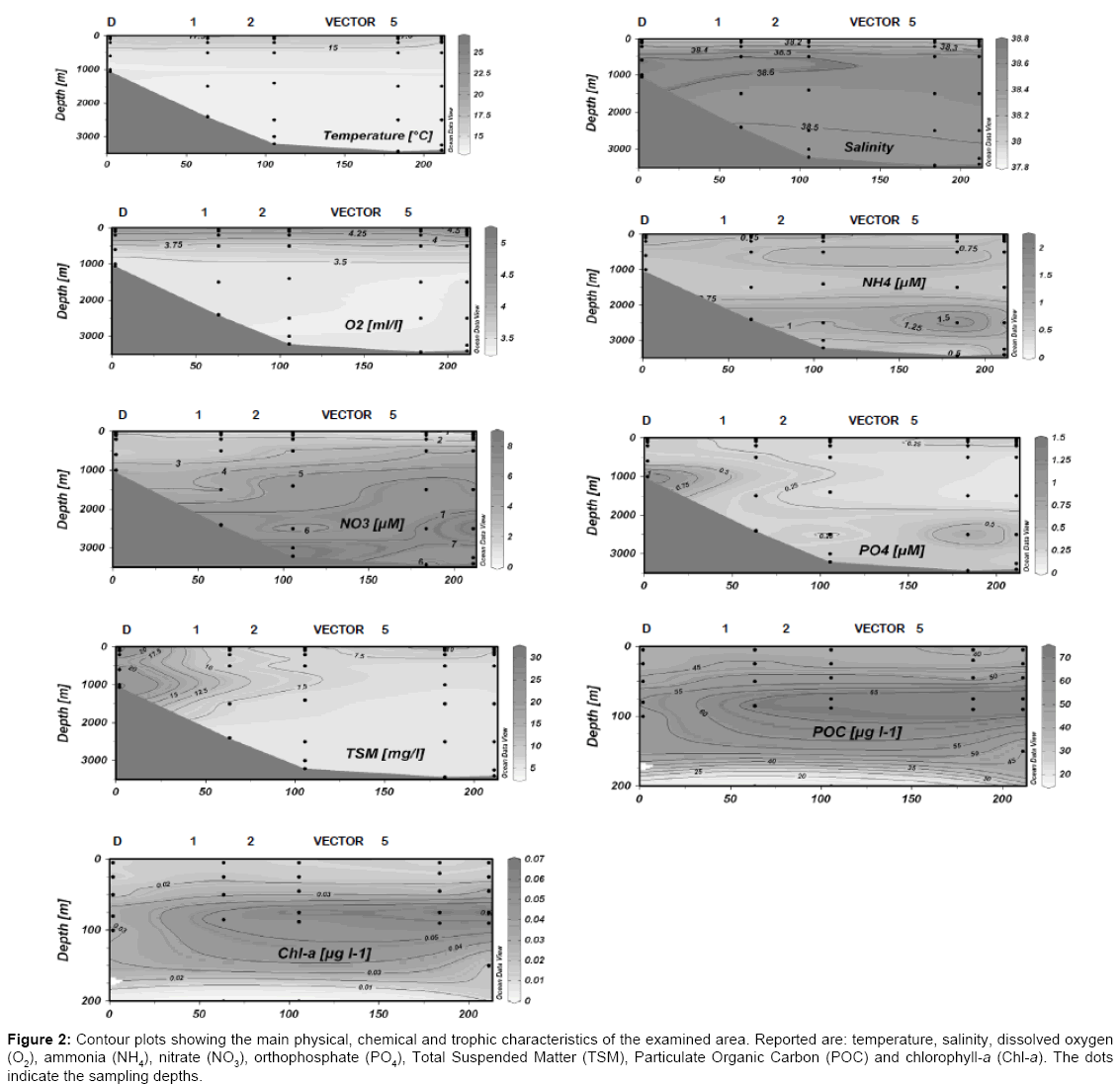
Figure 2: Contour plots showing the main physical, chemical and trophic characteristics of the examined area. Reported are: temperature, salinity, dissolved oxygen (O2), ammonia (NH4), nitrate (NO3), orthophosphate (PO4), Total Suspended Matter (TSM), Particulate Organic Carbon (POC) and chlorophyll-a (Chl-a). The dots indicate the sampling depths.
Prokaryotic parameters (activities and abundances)
The rates of prokaryotic activities and the values of prokaryotic abundances are reported as the mean value and the range of variation (minimum-maximum) in Table 1.
| Pelagic layer | LAP (nmol l1h-1) |
B-GLU (nmol l1h-1) |
AP (nmol l1h-1) |
HPP (nmol l1h-1) |
PA X 105 cell ml-1 |
R (µCl-1h-1) |
|
|---|---|---|---|---|---|---|---|
| EPI | Mean Min-Max vs MESO |
17.55 0.94-8189 ns |
0.59 0.01-2.39 |
3.60 0.04-10.34 ** |
0.0167 0.0047-0.0381 ** |
3.1 1.2-5.1 ** |
0.225 0.023-0.918 * |
| MESO | Mean Min-Max vs BATHY |
23.71 0.13-79.85 ns |
0.80 0.10-2.38 ns |
0.78 0.24-1.95 ns |
0.0015 0.0003-0.0035 * |
1.3 1.0-1.9 ** |
0.007 0.003-0.012 ns |
| BATHY | Mean Min-Max vs EPI |
37.72 1.47-112.67* |
0.40 0.01-2.35 ns |
0.93 0.09-3.21 ** |
0.0043 0.0005-0.0121 ** |
0.7 0.48-0.99 ** |
0.004 0.001-0.011 ** |
Table 1: Mean, minimum and maximum values of leucine aminopeptidase (LAP), b-glucosidase (B-GLU), alkaline phosphatase (AP), heterotrophic prokaryotic production (HPP), prokaryotic abundance (PA) and respiratory rates (R) measured in the different epipelagic (EPI), mesopelagic (MESO) and bathypelagic (BATHY) layers.
The spatial distribution of the depth-integrated and normalized values is shown per each station and per layer in Figure 3. LAP showed the highest activities in the water column followed by AP and b-GLU.
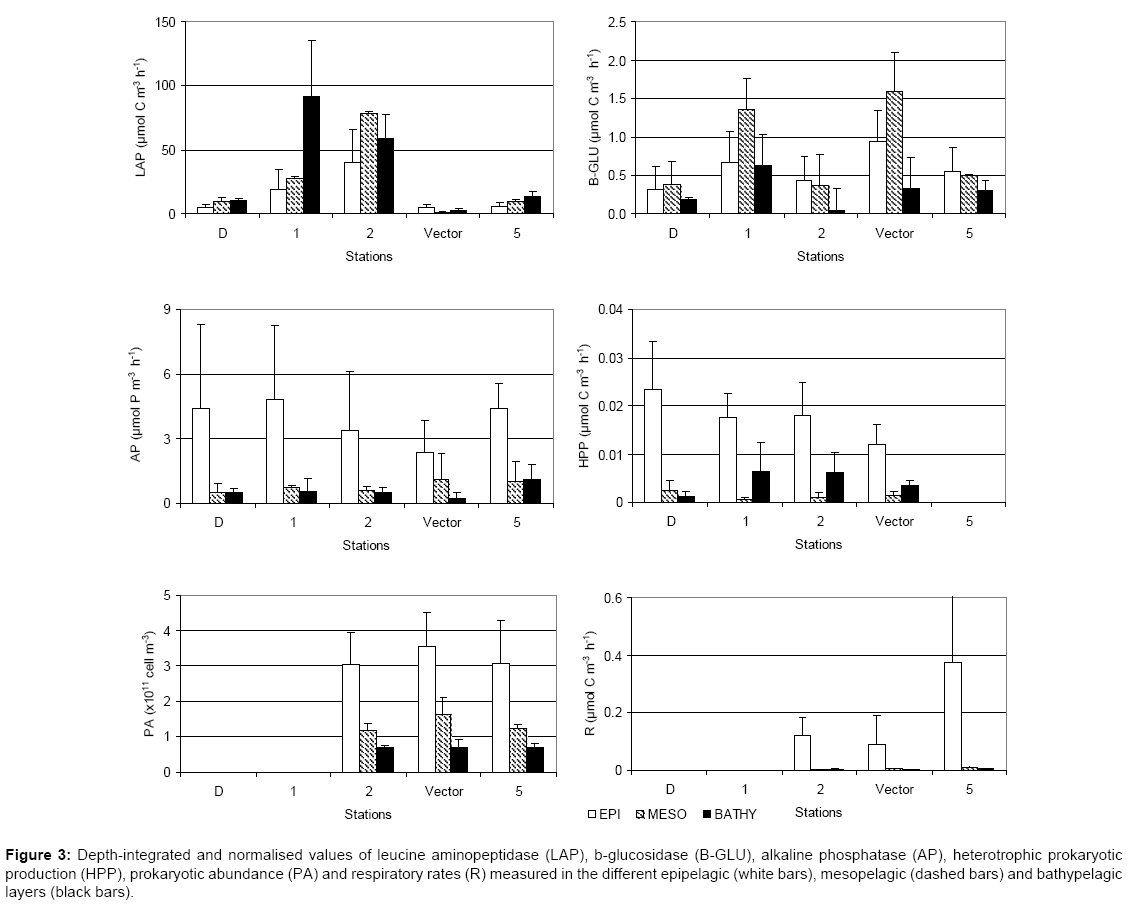
Figure 3: Depth-integrated and normalised values of leucine aminopeptidase (LAP), b-glucosidase (B-GLU), alkaline phosphatase (AP), heterotrophic prokaryotic production (HPP), prokaryotic abundance (PA) and respiratory rates (R) measured in the different epipelagic (white bars), mesopelagic (dashed bars) and bathypelagic layers (black bars).
LAP values gradually increased from surface towards the bathypelagic layer, with the highest values at the easternmost stations 1 and 2; activity rates were at their minimum at the station Vector. B-GLU values prevailed in the mesopelagic layers at the stations D, 1 and Vector. AP was predominant in the epipelagic layer; it decreased sharply at the meso- and bathypelagic layers; the highest values were recorded at the stations D, 1 and 5.
The values of the Michaelis-Menten constant (Km) (from epi- to bathypelagic layer, LAP: 197.16 ± 22.35, 266.01 ± 33.02, 153.71 ± 18.09; b-GLU: 16.67 ± 13.93, 19.70 ± 8.79, 4.71 ± 0.77; AP: 7.49 ± 0.02, 91.80 ± 12.06, 24.31 ± 3.24) were higher at the mesopelagic layer for all the three enzymes, particularly for AP. Conversely, the lowest values were obtained at the bathypelagic layer for LAP and b-GLU and at the epipelagic layer for AP.
HPP decreased from the epi- to the mesopelagic layer and from eastern to western stations; at bathypelagic depths it reached the maximum values at stations 1 and 2 (Figure 3). PA decreased from surface to bottom depths; the highest value was observed at Vector station at the epipelagic depth, while no variations in the cell numbers were found in the bathypelagic layer.
R values showed an abrupt decrease from the epipelagic to the meso- and bathypelagic layers; the maximum values were measured at the western station 5.
Statistical analysis
ANOVA results, shown in Table 1, highlighted significant differences occurring between epi- versus meso- and bathy-pelagic layers for AP, HPP, PA and R. HPP and PA values were also significantly different between meso- and bathypelagic layers. Significant differences were also found among the stations, in the patterns of LAP, b-GLU and TSM (ANOVA F= 28.95, P<0.001; 3.49, P<0.01; 47.93, P<0.001, respectively). Post-hoc comparisons showed that the station 1 accounted for the most significant differences in LAP and b-GLU distribution, while station D was responsible for differences detected in TSM patterns.
ANOSIM and pairwise comparisons confirmed significant differences between epi- and bathypelagic layers, with a R statistic of 0.635 (p=0.1%); significant differences occurred among the stations, with R statistics of 0.491 and 0.482 (p=0.1%) comparing station D versus stations 2 and 5. SIMPER analysis revealed that the most important parameters which separated the epi- and bathypelagic layers were nitrate and dissolved oxygen concentrations (with contribution percentages of 18.3 and 15.5%). The parameters which made station D different were TSM and PO4 concentrations (with contribution percentages of 26.7 and 19.6% for station 2; 32.3 and 21.1% for station 5).
Significant Pearson correlation coefficients were obtained, separately per each layer, among exoenzymatic activity rates and environmental and other prokaryotic parameters (Table 2). Inverse significant relationships were found between TSM and LAP, b-GLU, AP. AP was also inversely related to PO4 concentration in all the three layers. Conversely, positive relationships were observed between HPP and LAP at the bathypelagic layer, as well as between HPP and AP at the epipelagic layer. Significant relationships between PA and b-GLU were detected both at the epipelagic and bathypelagic layers.
| LAPvs: | B-GLUvs: | APvs: | ||||
|---|---|---|---|---|---|---|
| EPI | S TSM | **0.63 **-0.54 | TSM T HPP Chl-a PA | *-0.55 **-0.76 **-0.77 *0.59 **0.94 | S PO4 HPP R | **-0.61 **-0.62 **0.82 **0.78 |
| MESO | Chl-a R | **-0.89 **-0.91 | LAP | *0.56 | PO4 R | **-0.63 **0.90 |
| BATHY | T B-GLU HPP | **-0.72 **0.62 **0.89 | T PA | *-0.68 *0.71 | S PO4 TSM | **-0.55 **-0.56 **-0.58 |
Table 2: Pearson correlation coefficients calculated among enzymatic activity rates (leucine aminopeptidase LAP; B-glucosidase, B-GLU; alkaline phosphatase, AP) and other environmental and prokaryotic parameters, separately per each layer (EPI, MESO, BATHY, epipelagic, mesopelagic and bathypelagic layer). For abbreviations, refer to caption to Table 1.
The outputs of MDS analysis performed on exoenzymatic activity rates, shown in Figure 4, pointed out that there was a clear difference among the stations along the study transect. The MDS analysis identified six main groups. Station 1-Bathypelagic was closely related to station 2-Meso and 2-Bathypelagic; similarities were also found between stations 5 and D in their Meso-and Bathy-pelagic layers. Station 1 Mesopelagic was related to these two groups and to the group composed by the Epipelagic layers of stations 1 and 2. The group including together the deepest layers of station Vector (V-Mesoand V-Bathypelagic) was characterised by the lowest exoenzymatic activity rates. Finally, close associations among the D-E, Vector-E and 5-E samples were detected. In general, the Epipelagic samples grouped separately from the Meso- and Bathy-pelagic ones, which grouped together; no relationships were found between the epi- and bathypelagic layers.

Figure 4: Output of Multi Dimensional Scaling analysis performed on exoenzymatic activity rates. The codes indicate the station number (1,2,5, D and V, Vector) followed by the indication of the depth layer (E, epipelagic layer; M, mesopelagic layer; B, bathypelagic layer). The circles and the arrows indicate the main groups of samples and their reciprocal relationships; on the right, the strength of the relationships [from None, -, to low, (+) , discrete, +, and high, +++] between Bathy- and Mesopelagic (B-M), Epi- and Mesopelagic (E-M) and Epi- and Bathypelagic (E-B) is shown.
This study represents a contribution to knowledge of prokaryotic community in the Tyrrhenian Sea in terms of both abundances and activities. The role played by the prokaryotic community at different depths of the Southern Tyrrhenian Sea is evaluated by estimating the efficiency of some steps in organic matter processing (i.e. enzymatic hydrolysis, HPP and Respiration), so allowing to draw some considerations on the implications of prokaryotic metabolism on organic matter transformation.
Values of both prokaryotic activities and abundance measured in this survey fall in the same range reported in other aquatic temperate ecosystems [9,26,28,32-35]. Enzymatic activity rates along the transect highlight significant variations in the metabolic levels among the examined stations, as shown by the ANOVA results. Along the transect, the patterns of Carbon mobilization by exoenzymes and Carbon oxidation by respiration, although examined for three stations only, show a synergic cooperation between the two processes toward the mineralization.
Vertical profiles examined by ANOVA point out major differences both in Prokaryotic Abundance (PA) and activities (AP, HPP and R) when comparing the epi- versus both the meso- and bathypelagic layers. Hot spots of prokaryotic metabolism are found at the meso- (b-GLU) and bathypelagic (LAP, HPP) layers; similar results have previously been reported in the Mediterranean basin [24,26,28,31-32]. MDS analysis performed on exoenzymatic activity rates allows to distinguish between one group which includes together the meso- and bathy-pelagic layers and another one collecting together the epipelagic samples from the stations D, Vector and 5. The close associations observed between the meso- and bathy-pelagic layers by MDS and the high values of metabolic activities measured even at bathypelagic depths suggest the export of biogenic carbon produced in the mesopelagic layer towards the bottom. High enzymatic rates at bathypelagic depths could also be related to lateral transport of organic carbon. Lateral transport from the continental margins may provide a significant component of the mesopelagic and bathypelagic carbon budget in certain regions of the ocean, like the central Arabian Sea and North Atlantic regions [66].
Besides the Vmax, the values of the Michaelis-Menten Km constant give additional information on enzyme kinetics. Km is characteristic for each enzyme; the lowest is its value, the highest is the affinity of the enzyme for its substrate. This is obtained at the bathypelagic layer for LAP; high substrate affinity is considered a strategy to support bacterial growth under low nutrient conditions [67]. For AP, the highest affinity of this enzyme for organic phosphoesters is detected at the epipelagic layer, probably related to the Phosphorus consumption by phytoplankton and bacterioplankton, which stimulates the enzymatic synthesis. Moreover, the values of Km recorded in the Tyrrhenian area are significantly lower than those found at similar depths in the Atlantic Ocean [23]. This finding suggests a higher affinity of the Mediterranean prokaryotic assemblage to low amounts of organic resources compared to the Atlantic one, confirming what observed by Luna et al. [26].
With respect to the relationships among exoenzymatic activity rates and environmental and other prokaryotic parameters, the significant Pearson correlations calculated between TSM and LAP in the epipelagic layer suggest that 54% of LAP variability is explained by the variability of TSM. Similarly, TSM variability affects the variability of b-GLU (55%) and AP (58%) in the epi- and bathypelagic layers, respectively.
The variability in HPP is related to the variability in LAP (89%) and AP (82%) activity rates at the bathypelagic and epipelagic layers, respectively; the monomers released by the exoenzymatic activities are uptaken and support the production of new prokaryotic biomass. On the other hand, the growth of heterotrophic microorganisms depends on the availability of inorganic P. Since P is considered as a limiting factor for productive processes in the Mediterranean Sea and is actively regenerated from organic phosphates through AP [15,29], the negative relationships found between AP and PO4 explain the stimulation of enzyme synthesis played by low P availability.
During the present study, PA show significant differences across the water column. The positive correlations between PA and b-GLU at the epi- and bathypelagic layers suggest that the total prokaryotic community is actively involved in the synthesis of this enzyme. Moreover, the algal exudates released within the epipelagic layer contain high concentrations of bioavailable polysaccharides, therefore these compounds stimulate the activities of extracellular glucosidases [10].
Some speculations on the prokaryotic metabolism
Among the exoenzyme activity measured, the prevalence of LAP suggests that the C mobilized from proteinaceous material is quantitatively the most important source of Carbon and dissolved monomers for the prokaryotic community. Higher LAP than b-GLU activity rates are frequently recorded in other marine ecosystems [5,13]; on the other hand, protein-containing particles are very abundant in seawater and may act as an important C and N source for bacteria in the pelagic ecosystems [32,68]. High levels of proteolytic activity are expressed in response to the amount of protein matter released by the productive processes [68,69], although recent studies indicate that selective protein molecules may survive to microbial degradation and contribute to the pool of dissolved proteins in the sea [70]. Higher LAP and AP rates compared to b-GLU ones result in a preferential nitrogen and phosphorus cleavage with respect to carbon [20].
The reciprocal ratios between exoenzymatic activities (LAP/b-GLU and LAP/AP) are reported in Table 3. Variation in the relative activities of different enzymes is suggested as possible indicators of changes in the modalities of bacterioplankton nutrition [71]. Sinsabaugh and Follstad Shah [72] reviewed the reciprocal ratios among enzymes activities in plankton communities, suggesting connecting the “ecoenzymatic stoichiometry” approach as a tool to connect the elemental stoichiometry of microbial biomass and detrital organic matter to microbial nutrient assimilation and growth.
| LAP/b-GLU | LAP/AP | (LAP+b-GLU)/HPP | (LAP+b-GLU)/R | |
|---|---|---|---|---|
| EPI | 79.8 | 5.2 | 63.9 | 10.3 |
| MESO | 30.9 | 28 | 1523.1 | 46.6 |
| BATHY | 842.7 | 59.4 | 606.1 | 456.3 |
Table 3: Mean values, per each layer (Epipelagic, EPI; Mesopelagic, MESO; Bathypelagic, BATHY) of: enzymatic activities ratios (LAP/b-GLU and LAP/AP), Hydrolysis/Uptake ratio (LAP+b-LU)/HPP, Hydrolysis/Respiration ratio (LAP+b- GLU)/R. For abbreviations, refer to caption to Table 1.
Particularly, the LAP/b-GLU ratio provides information about the qualitative composition of degradable organic matter [15]; this ratio indicates the ratios between the two processes of proteolysis and glycolysis and depends on the functional features of the ecological system [71]. Its values may vary in a wide range, from an average of 0.28 in the Equator to 593 in the Southern Ocean waters [71]. Values of LAP/b-GLU ratio higher than 1 indicate a higher ability of prokaryotes to metabolize preferentially proteins rather than polysaccharides, and vice-versa. During this study the values of LAP/b-GLU ratio are always >1 and are higher in the bathypelagic layer than in the others, suggesting high metabolic potentialities of prokaryotes in the degradation of proteinaceous material in the deep realm. Along a Western-Eastern Mediterranean transect, Zaccone, et al. [33] found the prevalence of LAP with respect to GLU in the epi- and bathypelagic layers (LAP/b- GLU ratio ranged from 2.04 to 6.77 and from 1.07 to 7.46, in the two layers respectively), in the mesopelagic the ratio dropped to values between 0.29 and 0.95. A slight decrease of LAP/b-GLU ratio in the mesopelagic layer was supposed to be related to the availability of protein-derived materials for prokaryotes, which reduced the need for exopeptidases such as LAP [73]. In an oligotrophic area of the Southern Tyrrhenian Sea close to the area examined in this study, Misic, et al. [35] found LAP/b-GLU ratios ranging from 24 to 73.
Another ratio, the LAP/AP ratio has been considered to assess the nutrient limitation in the microbial population, as recognised in previous studies [15,28,74]. The LAP/AP ratios obtained in the Tyrrhenian waters are low at surface, indicating the quick remineralization of organic phosphates with prompt recycling of P in this layer. A low ratio between LAP and AP suggests that both enzymes equally satisfy the prokaryotic trophic needs, while the relative importance of AP decreases with increasing depth. However, different mechanisms are involved in the N and P regeneration: AP activity is responsible for the direct mineralization of organic phosphate esters into orthophosphate, while LAP acts in the first decomposition on proteins, with release of dipeptides which follow further decomposition pathways before complete mineralization.
To estimate the coupling between prokaryotic hydrolysis and uptake processes, the (LAP+b-GLU)/HPP ratio is computed. The high values of (LAP+b-GLU)/HPP obtained at the meso- and bathypelagic Tyrrhenian layers indicate that the amount of monomers released by the enzymatic hydrolysis largely exceeds that incorporated in HPP. Thus, despite the high hydrolysis rates, monomers are not efficiently consumed by microorganisms. In the northern Adriatic Sea low ratios of enzymatic activity (LAP) to HPP rates were found in June 1997 [8]; this suggested a good coupling between these two processes, with a high probability of incorporation into new biomass of the Carbon released by exoenzymatic hydrolysis. Indications on the hydrolysisuptake coupling in bacterial utilization of proteins were also provided in a previous study by Hoppe et al. [75], who proposed the use of a Leucine turnover: hydrolysis ratio as an indirect measurement of the pool size ratio of polymers to monomers. The coupling between hydrolysis and uptake processes greatly increases the efficiency of organic matter utilization by prokaryotes in aquatic environments. Moreover, variations in the hydrolysis to uptake ratios depend not only on the nature (polimeric or monomeric) of natural substrates, but also on the bacterial-organic matter interactions at single cellbasis. Differences in the substrate hydrolysis to uptake ratios were reported between free-living and particle-attached bacteria; in particle associated bacteria, substrate hydrolysis might exceed substrate uptake, while free-living bacteria were more efficient in the incorporation of the small molecules resulting from enzymatic hydrolysis [8,10]. Thus, free-living bacteria benefit from the monomeric compounds (free aminoacids and monosaccharides) released from attached bacteria, which may provide a suitable carbon and nitrogen source to fulfil their metabolic needs.
Also the LAP+b-GLU/R ratio in the epipelagic Tyrrhenian layer shows that the two processes of enzymatic hydrolysis and final oxidation by R are reciprocally balanced; thus, in this layer, the oxidative processes (R) are highly efficient in organic matter processing and both processes act synergically towards final organic matter mineralization. This suggests a quick turnover of organic matter in the epipelagic realm. At the bathypelagic depth, the uncoupling between the enzymatic hydrolysis and R is more evident, as shown by the high values of LAP+b-GLU/R ratio. The increase of the pool of dissolved matter released by hydrolysis, not associated by mineralization, could result in a reduced efficiency of Carbon turnover through the prokaryotic community and favour the sinking of matter not fully degraded at depths.
To quantify the Carbon flux within the prokaryotic community the Prokaryotic Carbon Demand (PCD, as the sum HPP+ R) and the Bacterial Growth Efficiency (BGE, as the percentage ratio between the processes of prokaryotic production-HPP and consumption by PCD) have been calculated (Table 4). BGE - This provides a descriptive index of the nutritional environment for marine bacteria - increases from surface to deep layers. In the aphotic zone, the BGE of the bathypelagic layer is about three times higher than the value recorded in the mesopelagic one. This trend suggests that in this last layer HPP alone is enough to satisfy PCD. Tanaka [76] claimed that in the mesopelagic layer the majority of the particulate organic carbon exported from the epipelagic layer is remineralized to CO2 and that data on prokaryotic growth efficiency on an annual scale suggest that mesopelagic prokaryotes are not simply remineralizers of organic carbon, but also play an important role in supporting the production of organisms belonging to higher trophic levels.
| HPP | R | PCD | BGE | T (Pearson r) | POC (Pearson r) | |
|---|---|---|---|---|---|---|
| mgCm-3d-1 | mgCm-3d-1 | mgCm-3d-1 | % | BGE-T | BGE-POC | |
| EPI | 0.401 | 5.52 | 5.921 | 6.8 | 17.21(-0.725**) | 49.21(+0.427*) |
| MESO | 0.036 | 0.168 | 0.204 | 17.6 | 14.02(-0.553*) | |
| BATHY | 0.103 | 0.096 | 0.199 | 51.8 | 15.50(+0.273) |
Table 4: Mean values, per each water column layer (Epipelagic, EPI; Mesopelagic, MESO; Bathypelagic, BATHY), of: Heterotrophic Prokaryotic Production (HPP), Respiration (R) as the amount of Carbon released from organic matter remineralisation, Prokaryotic Carbon Demand (PCD=HPP+R), Bacterial Growth Efficiency (BGE=HPP/PCD), mean Temperature (T,°C) and Particulate Organic Carbon (POC, µgC l-1).and Pearson correlations versus BGE (in brackets). The significance of the statistical correlations is shown (*, P<0.05, **, P<0.01).
BGE values found in the epipelagic Tyrrhenian layer confirm the wide variability of BGE reported by previous studies [9,63]. They are similar to other estimates (2-8%) reported from the Mediterranean Sea [51]. In most freshwater and marine systems BGE, ranges from less than 10 to 25%. In the present study, BGE is negatively related with T in the epi - and mesopelagic layers (Pearson r= -0.725, P<0.01, and -0.553, P<0.05, respectively), in agreement with Rivkin and Legendre [77]. In the Tyrrhenian waters, the increase of BGE with increasing depth is in contrast to lower values recorded in the bathypelagic layer of the dark ocean [18]. Since high BGE values mean high CO2 mobilization, microbial communities seem to act as sink of CO2 in the deep Tyrrhenian biota. BGE increases with depth could depend on higher temperature (never < 12°C) which characterizes the deep Mediterranean, compared with open ocean waters. Moreover, its vertical increase may be related to changes in the rate of supply and nutritional quality of organic substrates for prokaryotes; in the epipelagic layer BGE is positively related with POC and Chl-a concentrations (Pearson r=-0. 424 and 0.427, P<0.05, respectively) as well as in the whole column (r=0.409 and 0.417, P<0.05, respectively). The increase of BGE with the depth suggests that BGE does not depend exclusively on PP and its related export production. Moreover this finding lead us to hypothesize that the deep Tyrrhenian basin is more dynamic than other oceans or that organic matter has not been degraded yet; the results obtained in this study could also be linked to the supply of preformed C transported within the circulatory pattern of the Mediterranean water masses.
In conclusion, in the examined Tyrrhenian area the general decrease of activity rates and abundances with increasing depth suggests the predominant weight of export production in this basin. Both the decomposition and biomass production processes are high at the epipelagic layer. Conversely, at the mesopelagic one the organic matter is quickly mobilised but not efficiently incorporated into biomass; a loose of coupling between these two processes led us to hypothesize that the dissolved monomers remain available in the water column for export to other trophic levels or utilization in other processes (lateral advection, oxidation). The bathypelagic layer is characterized by high levels of hydrolytic activity coupled with relatively high prokaryotic production; in this layer, exoenzymatic activity levels fall in an order of magnitude similar to that observed at the epipelagic one. This finding highlights the importance of microbially mediated processes in sustaining life in the marine depths. Current knowledge on microbial dynamics and processes in this pelagic zone of the Tyrrhenian Sea is still far to be exhaustive, and further research in this field is needed to improve knowledge of the ecological role of microorganisms in the ecosystem processes.
The authors are grateful to the two anonymous referees who improved the first version of this manuscript with their valuable suggestions. This research was funded by the research project “Biodiversity and organization of the communities in different environmental contexts” funded within the FIRB-MIUR program.