Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Review Article - (2015) Volume 4, Issue 5
Noroviruses are non-enveloped, single-stranded viruses that are part of the family of viruses called Caliciviridae. Emergence of different geno-groups was as a result of genetic combinations. Expression of histo-blood group antigen (HBGA) corresponds with an organism’s response to infection. Less than 10% of plant species on the earth have been utilized by human being, which indicates that more bioactive compounds are yet to be discovered. The outbreak of norovirus infections is made possible by their infectivity at freezing temperatures, accidental swallowing of virus-containing droplets, ability to resist of common disinfectants, contact with individuals lacking obvious symptoms, and meeting with recuperating persons - uncontrollable risk factors. Conversely, manageable risk factors include contact with diarrhea patients, frequency of cutting fingernails, ingestion of uncooked food, and households with looseness of bowels. Various methods are used to extract the phytochemical used in preventing the norovirus infection. These phytochemicals are effective at various concentrations in modulating bacteria, fungi and viral infections including those of mice and chimpanzee that mimic human infections. Inherent bioactive compounds in these phytochemicals are credited with affecting the microorganisms. The HBGA attachment of P-particles is effective in a dose-regulated fashion with various extracts. The Cranberries reduce the impact of the HBGA-interaction, which is achieved by permanent mutilation or temporary inhibition of some areas of the capsid protein. Virus-like particles (VLPs) such as Baculovirus and Venezuelan equine encephalitis replicon systems developed from genetic manipulation of the capsid protein of the norovirus have been instrumental to the study of norovirus pathogenicity. The noroviruses, example GII.4 viruses, continue to stay in human mainly by herd immunity and adaptation to the found environment.
Time of exposure and, the amount of phytochemicals play important roles in modulating norovirus infections.
Keywords: Norovirus; Phytochemicals; Prevention; Therapeutics
The norovirus (NoV) belongs to the family of viruses called the Caliciviridae. Caliciviruses are non-enveloped viruses, having 27 to 35 nm in diameter, single-stranded RNA positive strand genomes of 7 to 8 kb size [1].The genome encodes three open reading frames (ORFs) [2]. Each of the three ORFs encodes different proteins. The three ORFs are differentiated into 1-3. ORF1 encodes polypeptides, which have regions resembling the helicase, cysteine proteinase and RNA-dependent RNA polymerase. ORF2 encodes a viral capsid protein while ORF3 encodes a little protein whose function is yet to be determined [1,3].
The Norovirus is assumed to be the cause of almost half of all gastroenteritis outbreaks world-wide and 75%-90% [4], of gastroenteritis not caused by bacteria in humans of all ages [2,3]. About 23 million cases of gastroenteritis caused by norovirus are projected to occur in the United States each year and an estimated of nearly 60-85% of all gastroenteritis outbreaks within the United States [5], Europe, and Japan are linked with NoV (Truong et al.). Moreover, genome differentiation studies have placed NoV into five separate geno groups (G1-GV), a sixth genogroup was newly discovered [6]. The five genogroup was further subdivided into 44 genotypes made of 16 G1, 23 G11, 2G111, 2G1V, and 1GV. The emergence of these genotypes was purportedly as a result of genetic recombination of these genotype [3]. Human noroviruses (HuNoVs) occurrence usually originate from healthcare facilities (e.g., nursing homes, residential care centers, hospitals), schools [7], day-care-centers, food service establishments, recreational settings (such as cruise ships [7] and hotels [2]. They also occur in military barracks and in assisted living communities giving rise to about two-hundred and sixty-seven million yearly outbreaks worldwide [8].
Norovirus epidemics occur most readily in partially closed communities, such as cruise ships, disaster relief/evacuation site, nursing homes, hospitals, schools and military settings [9]. The sporadic nature of norovirus infection are attributed to increased infectivity of norovirus particles, continuity of norovirus surroundings, long-term shedding of virus from both asymptomatic and symptomatic persons, as well as absence of lasting immunity [9]. Symptoms of the norovirus infection include diarrhea, vomiting, and sometimes fever [5], although asymptomatic infection may occur occasionally [10]. It should be noted that GII.4 strains are responsible for the majority of outbreaks globally. The spread of norovirus is basically through the fecal-oral path [7] which entails direct or indirect contact with dirty environmental surfaces, ingestion of dirty water and food [11] and also direct contact with infected persons [5]. Certain inherent characteristics are responsible for the incessant cases of norovirus infections in the environment. These include its infectivity at freezing temperatures, accidental swallowing of droplets that contain these viruses, defiance to effects of common disinfectants, to mention a few. Also, making contact with individuals lacking obvious symptoms and meeting with diseased, but recuperating persons, make norovirus infections tough to manage [5].
Some manageable risk factors are associated with the norovirus infection. The factors had been tested with single factor logistic regression analysis and proven to be risk factors for this dreadful infection, and include contact with diarrhea patients, frequency of cutting fingernails, ingestion of uncooked food, and households with looseness of bowels. In addition, the rate of consuming seafood, and eating commercially prepared meals significantly add to the risk of getting the norovirus infection. Also, frequency of hand washing, proper washing of vegetables and fruits before consumption, and observing proper hand washing as at when due will limit the spread of this disease [7].
Certain groups of individuals including the immune-compromised, the elderly and young children are at increased risk of infection with norovirus and its later complications. Others like food handlers and health care workers are at greater chance of norovirus transmission [12]. Noroviruses have been implicated in the cases of diarrhea in pediatric transplant patients and are with known clinical complexities. Equally, norovirus diarrhea patients are known to experience more common ICU admission than non-norovirus diarrhea patients [13]. Noroviruses can render a long-lasting infection in immune-compromised hosts, giving rise to gastrointestinal disease, which may lead to wasting and debilitating diseases, as well as persistent virus shedding. Inherited traits like immune disorders, small-bowel transplantation, cancer or cancer therapy, kidney transplantation and associations that could lead to immunosuppression in children have been known for norovirus infection. In adults, deteriorating health conditions have established a link to heightened norovirus infections, as reduced immune functions are a major contributory factor [14].
It is certain that proper hand hygiene would drastically reduce the spread of the norovirus infections. Strict adherence to proper hand washing and prevention of norovirus contamination of foods could be more efficacious than treatment. Hence, adequate diligence to hand hygiene will obviously improve the quality of fresh produces. Other hygienic processes like water quality regulations need to be strictly adhered to [15]. In this article, we use two words “Norovirus” and “Phytochemicals” to screen the research as well as review articles. We do not follow only one database, we used various database randomly using two words only.
The use of herbal plants to treat human diseases has opened an alternative door for treating some of the diseases that are eluding orthodox medicine. The surge in traditional plant use to find permanent solutions to ever emerging human’s disease may provide the longawaited answer to handling human noroviruses. These plants have proven to possess anti-inflammatory, antiviral, antitumor, antimalarial and analgesic properties [16]. Non-bacterial gastroenteritis is an emerging threat that is currently ravaging the world, and greater than 75% gastroenteritis not caused by bacteria are coming from noroviruses. This cuts across humans of all ages [3].
It has been proven that there is a link between human genetic factors and norovirus susceptibility. The availability of a secretor status (presence of a functional fucosyltransferase2 gene) and the manifestation of histo-blood group antigen (HBGA), have proven to change an individual’s susceptibility to norovirus infection [17]. Fucosyltranferase2 (FUT2) controls the secretion of ABO HBGAs at the gut surface. HGBAs are inert carbohydrates found on lipids or proteins which are joined by different HuNoV strains and their expression corresponds with an organisms response to infection [18]. These carbohydrates bring together ligands and supposed receptors for Caliciviruses, of which the norovirus is a member [19]. However, a set of individuals is considered to be non-secretors of this gene and they constitute of about 20% of the white population. The non-secretors are so termed because the FUT2-dependent HBGAs are not expressed in them. Contrarily, the remaining set of the individuals (about 80%) carries at least one functional FUT2 allele and are known as ‘secretors [19].
Studies have shown that out of about five hundred thousand species of plants on earth, only 1-10% are utilized by man [20]. There are lots of bioactive compounds that are yet to be identified both in those plants that are familiar to humans and those yet to be investigated. One of these eminent bioactive compounds is the phenolic compound. This group of compounds encompasses greater than eight thousands compounds known to possess a single aromatic ring and a single or more hydroxyl groups attached [21]. Phenolic compounds are divided into flavonoids and non-flavonoids based on the arrangement as well as the number of their carbon atoms. These carbon atoms are often in conjugated forms with sugars as well as in biological acids [21]. The availability of these natural compounds in plants calls for more intensive scientific research towards discovering the therapeutic potentials and be able to unveil them in the near future. The components of phytochemical that could be examined include, but not limited to flavonoids, phenols, terpenoids, anthraquinonoid, glycosides, and steroids in different plant extracts [22]. The primary objectives of this review article are to find out if phytochemicals would proffer a permanent solution to the dreaded norovirus infection/gastroenteritis, to elucidate on the pathogenesis and mechanism of action of norovirus infection, to review the impacts of these natural chemicals on other microorganisms, and to compare the effects of different plant extracts on the norovirus surrogates. The above-mentioned objectives are based on the hypothesis that phytochemicals modulate the effects of noroviruses, and that natural extracts of plants are potentially the solution to the nonbacterial gastroenteritis.
Novel scientific studies are paving way for the use of phytochemicals in abating viral infections. A number of scientists have channeled their research efforts in this direction. For instance, in developing countries, human papilloma virus infection occurs concomitantly with many down-regulated signaling pathways, which are prone to cervical carcinogenesis. Transforming growth factor beta (TGF-β) signaling has been shown to promote tissue alterations encouraging cancerous build-up [23]. TGF-β is involved in numerous cellular processes both in adult organism and in developing fetus. Amongst their functions are cellular responsibilities, homeostasis, apoptosis, cell growth and cell differentiation as well as other cellular processes. The procedure of TGF-β pathway involves binding of a type II receptor which employs and phosphorylates a type I receptor. SMADZound that curcumin and emodin efficiently down-regulated TGF-expression and also, according to their result, TGF-β was found to be a critical player in Wnt/β-catenin signaling pathway by lowering the manifestation of TGF-β receptor II, P-Smad3 and Smad4. Curcumin and emodin collaboratively decrease cell population and migration in SiHa and HeLa cells. Equally, cyclin D1, p21 and Pin1 signaling pathways wereshut down upon the use of curcumin and emodin treatment. Emodin is a potent selective inactivator of the enzyme 11β-HSD1-a pleasing therapeutic target of type two diabetes and metabolic syndrome. It is a natural product and possesses bioactive effects in Chinese herbs [25]. Curcumin is a constituent of spice turmeric. It has proven to offer elaborate solutions to a vast number of human problems ranging from antioxidant, antiviral, anti-inflammatory diseases, diabetes, arthritis, to anticancer, Alzheimer’s disease and various long-lasting illnesses [26]. These are modulated via the regulation of numerous transcription factors, protein kinases, growth factors, inflammatory cytokines and other enzymes [26].
Another study x-rayed how dengue viruses could be targeted using the principle of molecular docking. It was found that flavonoids, a potent constituent of phytochemicals, can inhibit Asn-130 glycosylation site where Dengue virus nonstructural 1 (NS1) is attracted for its onward propagation [27]. It has already been documented that patients with Dengue hemorrhagic fever are more prone to dengue virus infection than those with only Dengue fever. This was attributed to increase in quantity of NS1 in their blood. NS1 encompasses triple N-linked glycosylation points at Asn-130, Asn-207 [28], and Asn-175 (27). N-linked glycosylation is the most prevalent type of glucosidic bond, and very critical for the folding of eukaryotic proteins as well as cellextracellular matrix and cell-cell attachment. It can equally control the function of proteins, acting as a control switch [29]. Qamar et al, found out that barricading the Asn-130 from NS1 will obviously stop replication of Dengue virus (DENV), even as flavonoids offer strong evidence of their ability to prevent proliferation of dengue virus. There is a strong bond-effect between NS1 and flavonoids and this positive interaction offers great hope for treating patients with dengue virus infection in the near future.
For many years, plant extracts have proved to have therapeutic remedies on a wide range of human illnesses. Human models, example, mice noroviruses have been grown in the laboratory, and at the same time they share the same genus as the human noroviruses [30]; hence are excellent models for investigating human noroviruses. However, clinical results, host receptors, and the types of cells that are infected [30], are significantly different. Ueda k. et al. [31] opined that tannins are therapeutically effective against a wide range of viruses: both enveloped and non-enveloped ones. It was found that mouse norovirus responded positively to tannin. Their experiment, comprised using extracts from persimmon (Diospyros kaki), comprises 22% of persimmon tannin, which drastically diminished viral infectivity above 4-log scale compared to other tannin extracts examined.
In another study, the outcomes of 13 food phytochemicals using protruding (P) particles of human NoV GII.4 as a research alternative in the study of attachment tendency of human norovirus were investigated [30]. The P-particle was initially observed while observing the reaction between the host HBGAs, the binding factors of noroviruses and the human NoV capsid [32]. It was further found that there were both S and P- domains in the NoV capsid, and these domains are organizationally and operationally separate. The main difference between the two particles is that S-particle does not attach to HBGAs, even though it has close skeletal resemblance to the inner shell of the norovirus capsid.
The P-domain compounds showed attachment function to HBGAs, portraying that P-domain is the HBGA domicile [33]. P-particles are known to consist of only the P-domains of the NoV capsid and devoid of the preserved shell domain [34]. The study yielded improved and diminished binding capability of phytochemicals to the NoV viral receptor using enzyme immune-sorbent assay as well as saliva-binding methods in triplicate form [35]. The outcome of the study indicates that the oyster extract possibly restrained the HBGA attachment of NoV P particles in a dose-regulated fashion from the lowest to the highest concentration. However, in the mussel extract, the lower the concentration, the better the suppression. Further, in hard clams and razor clams, the degrees to which binding was thwarted were rarely noticed even in high amounts. Similarly, studies on phytochemical effects of different kinds of berries on the HGBA-interaction tendency of human norovirus were carried out. It was found that Cranberry and Cranberry-pomegranate lowered the impact at various concentrations; with Cranberry juice exhibiting better anti-viral effect [35]. On the contrary, extracts of blueberry, blackberry, raspberry, and strawberry were not as efficient as the ‘Cranberries’ in lowering the attachment of NoV P-particles. Li et al, [35] opined that pomegranate and cranberry juices both are effective at impeding the colonization of human NoV surrogates, efficiently. They achieve this by combination of viral capsid protein with inherent polyphenolic chemicals in many soft fruits, which could lead to permanent mutilation or temporary inhibition on some areas on the capsid protein. This further elucidates that cranberry and pomegranate juices clearly disrupt NoV attachment [35]. However, owing to the softness of some fruits, especially the raspberry, they are more prone to human NoV retention due to the difficulty associated with proper washing of the berries, manual picking, and their inability to adequately restrict capsid attachment [35].
Furthermore, Dan Li et al., [30] reported that anti-NoV effect of GSE (grape seed extract) was investigated using RT-PCR, Caco-2 cells as well as pig gastric mucin as sensory nerve endings that receive and transmit stimuli. The study was done taking note of the attachment pattern of human noroviruses, considering the great advantage of Caco-2 cells, which includes manifestation of H types 1 and 2 of human HBGAs that are able to attach to a vast range of human NoVs from either G1 or G11. It was discovered that on addition of phosphate buffered saline (PBS) -and dried milk diluted human NoV G11.4 P particle, human NoV G11.4 fecal suspension, and MNV-1 lysate prior GSE treatment of the samples (Figure 1). It was shown that dried milk interrupts the effect of GSE on human NoV, and as the amount of dried milk increases, so does the level of inhibition to human NoV infection Li et al. [35]. To confirm that the protein disruption alters the viral capsid, microscopic examination was carried out after GSE treatment. The result revealed lump formation of virus-like particles (VLPs), clear deformation and bloating. At greater dose of 2 mg/ml, a huge protein concentration was seen, and VLPs vanished [35].
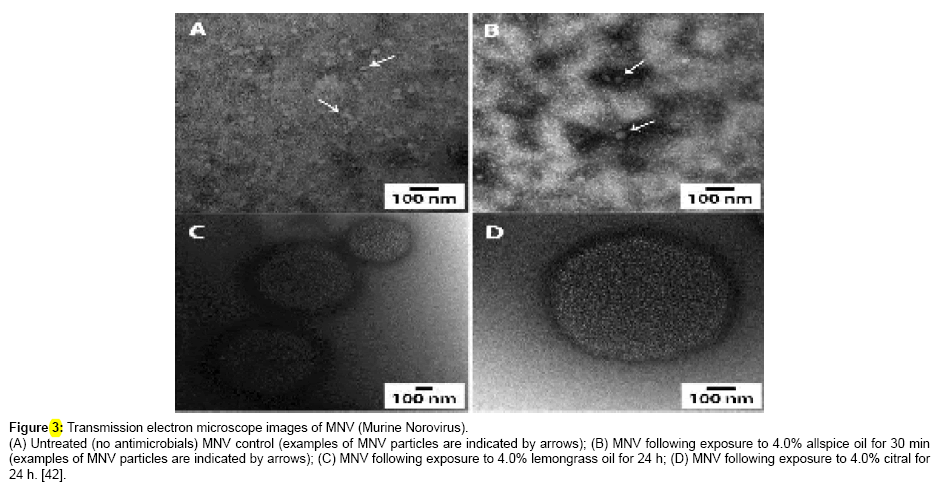
Figure 1: Transmission electron microscope images of MNV (Murine Norovirus). (A) Untreated (no antimicrobials) MNV control (examples of MNV particles are indicated by arrows); (B) MNV following exposure to 4.0% allspice oil for 30 min (examples of MNV particles are indicated by arrows); (C) MNV following exposure to 4.0% lemongrass oil for 24 h; (D) MNV following exposure to 4.0% citral for 24 h. [42].
The effect of antimicrobials (phytochemicals) to the structural changes innorovirus, typically the murine norovirus (MNV) particles were carried out to find if there were obvious structural changes to the virus particles. Transmission electron microscopy (TEM) was directly used to monitor the process [36-42].
The experiment recorded that very few particles of MNV could be found. The author confirmed that bigger amount of debris may be masking the appearance of MNV on the grids, hence the need for the 30-minutes exposure, which tentatively has less effect to the virus particles, confirmed from the outcomes of phytochemicals. This study has shown that lemongrass oil, allspice oil and citral demonstrated a significant viral reductions within 6 h contact with MNV [42].
The antiviral properties of phytochemicals are tested through time-of-addition experiments to viruses, wherein viruses are pretreated with test compounds before infection was initiated. Pretreatment with the tested compounds are performed to evaluate the ability of the phytochemicals to inactivate receptor(s) hence preventing viral attachment and internalization. In addition, pretreatment test methods produce anti-noroviral activity resulting from immune enhancement of the host cells [2]. Phytochemicals known to possess anti-noroviral activity of natural compounds include the following: Flavonoids are found in most biomaterials with anti-NoV activity, including grape seeds [43], pomegranates [44], mulberries [45], black raspberries [46], cranberries, and green tea extracts [47] and persimmons [48]. Other compounds, such as ginseng oregano essential oils, chitosan, and citric acid, as well as common proanthocyanidins and flavonoids from plant foods also effectively reduce NoV replication. Studies have also shown that myricetin, L-epicatechin, tangeretin, and naringenin produced significant anti-norovirus activity in a plaque reduction assay [49].
Studies have shown that copper nickel (number C70600) of the Unified Numbering System for Metals and Alloys (UNS) was found to be effective at deactivating MNV-1 even when juxtaposed with a group of metals that has 6% extra copper. A recent study has documented that MNV-1 was drastically deactivated on alloys 79-89 Copper, however, efficiency (efficacy) was lost at 70%. This implies that a minute change in copper content 70-79% could have a tremendous effect on antiviral strength, with the optimal deactivation of MNV-1 being noticed during the first 30 minutes of exposure [36].
Understanding the pathogenesis of human noroviruses has not been easy because they cannot be cell cultured, but for the murine norovirus that has been cultured [37]. Most of what is known about its infection pattern is from observations made from human volunteers [8]. and the tissue and biochemical studies carried out on them. In addition, some elucidations have been made using cow, pig and mouse models [9,37]. The inability to culture human noroviruses in the laboratory has not offered the opportunity of studying this group of viruses just like the other human models, example the MNVs. Absence of this study may continue to keep us in the dark regarding subtle changes that may have been revealed. However, the study of human noroviruses has to move a step forward in the direction of using advance assays like the RT-PCR, which entails studying the virus at genetic and molecular levels. The various genera of norovirus permit the existence of the norovirus in the population [8]. The MNV model has been used to postulate many theories about the way the norovirus interacts with the host in order to cause infection. Studies have shown that mice that lack downstream signal transducer and activator of transcription 1 (STAT1) are ultimately vulnerable to lethal norovirus infection. STAT regulates various aspects of differentiation, survival, motility and growth in cells. Seven mammalian STAT families are known, and these are STAT1- STAT6, with STAT5 having A and B types. Each of the seven STAT family members is known for a designated biological role and is present in lots of processes ranging from regulation of immune responses, cell differentiation to development of embryo. More so, they are known to be intrinsic part of the amino-terminal portion (NH2) of the preserved areas, whereas the carboxyl-terminal adds to STAT specificity [38]. They remain unexpressed in the cytoplasm until triggered by receptorrelated (associated) tyrosine kinases.
Earlier experiments presented a lone amino acid to be responsible for the dangerous effects of the MNV in 129/STAT1−/−mice after analyzing two genotypes of the MNV1 varying only at two residues [39]. However, more recent efforts by the same authors limit the role of this amino acid at position 296 of the virus capsid within the body of B6/STAT1−/−mice. Instead, they postulated that something more significant is responsible for the virulence in viral protein 1’s (VP1’s) P-domain. Thorough study of human noroviruses point to the fact that subdomain P2 possesses dynamic antigen and receptor binding sites which facilitate the most harm caused by MNV [39].
In these studies, samples taken from the infected human participants revealed gastrointestinal disruptions following introduction of the norovirus GI (Norwalk; GI.1) or GII (Hawaii; GII.1). Certain humans, however, do not respond to infection with a particular NoV, which may be due to a previous infection or complexity of their immune responses. This may pave way in the future for personalized studies that will exploit the factors in these individuals, which make them resistant to infection to better understand treatment options as well as preventive measures in the management of norovirus infections. In the vulnerable human subjects, the villi, microvilli and cytoplasmic content were altered and accumulation of tissue fluid occurred between cells, yet electron microscopy showed the cells not to be damaged. Similarly, Cow GIII.2 norovirus (BoNoV) studied elsewhere caused different levels of continuous watery bowel flow and weakness in infected tender calves, but without causing much damage to the guts. There was also significant shedding of the virus in the feces, which may account for the continuous presence of this virus infection in the cattle population [40].
In wild-type mice given oral or nasal administration of MNV-1, there is evidence of ribonucleic acids of the virus in various organs, including the liver, spleen and early part of the gut, even though the length of incubation, age and type of mice determine whether the viral RNA can be found in various body organs and feces [37]. This consolidates the fact that virus infection can be severe, but certain attributes like hereditary factors, state of immunity, age, environmental exposure (could be due to epigenetics), can play a role in the individual’s disease manifestation. A human subject experienced delayed bowel movements, perhaps due to frequent emetic episodes associated with the infection, which in mice resulted in enlargement of the stomach. Sugar metabolism of D-xylose and lactose is altered as well as fat absorption when infection is acute [9].
Both MNV and human (HuNoV) have been demonstrated to move across the interior of a cell, however, evidence has not been documented that the cells of immune-competent recipients have been infected [18]. NoVs infect innate cells, for example, in vivo, MNV antigen is observable in cells responding to macrophage marker F4/80 [18]. It has been documented that noroviruses infect immune cells, typically, the macrophages and the antigen-presenting dendritic cells in-vitro. The viral particle upon entering the epithelial cells will meet with the immune cells in the peyer’s patches,lymphoid follicles and lamina propria. Also animal models which include immunodeficient mice and chimpanzees were exposed to HuNoV illness.They showed evidence of harboring viral foreign substances in known macrophages and established dendritic cells [18]. In adaptive immunity, NoV has shown to target the B cells, although its invasion is absolutely different from those of the macrophage and dendritic cell penetration (innate immunity). Equally, in vivo, NoV attacks B cells, and this has been shown in the diseased mice B cells that are close to peyer’s patch areas. It has been documented that virus attached to HGBAs shown on the host intestinal absorptive cells speed up the holding in the intestine and to disrupt peristaltic movement of particles [18].
The influence of three essential oils (EOs), zataria, clove, and oregano on murine norovirus (MNV) and feline calicivirus (FCV) (norovirus surrogates) was measured using varying dilutions [41]. In the study, it was unveiled that 2% of oregano EO reduced the FCV titers by 3.75 log TCID60/ml, and with the same concentration, lowered MNV titers by 1.04-1.62 log TCID60/ml at 37°C. Another study evaluated the effect of all spice oil, lemon grass oil and citral on MNV. It was deduced from the study that all the three produced significant reduction when compared with the unmodified control, with allspice oil exhibiting the highest effect in cell permeability, closely followed by citral and, then lemon grass oil [42]. Time of exposure of phytochemicals as well as amount affects the level of inhibition of the viruses, and so this was the case for the allspice oil. However, other phytochemicals, example lemon grass oiland citral seemed to be exceptionally influenced by length of contact. For detailed examination, the effect of RNase 1 digestion was used in treating the viral particles and analyzed using RT-qPCR. The outcome amazingly showed little or no reduction in the viral effect between the virus particles treated with or without RNase 1 digestion. The effect of this partial or absence of digestion could be traced back to the fact that the viral RNA are currently protected from the enzyme via other avenues which may include antimicrobials occluding the surface of the capsid or that the viral coatings have not been degraded properly by the enzyme, hence shielding the RNA from RNase 1 digestion [42]. To make the picture clearer, one could say that the phytochemicals may be obscuring the viral epitopes needed for effective adsorption of the virus to the host cells. For instance, after being introduced in a MNV cell culture and left for 24 hr, 4.0% citral and lemongrass oil produced a decrease in culture infectivity of > or = 2.74 log10, and this resulted in partial decrease in the number of viral genetic make-up in the presence or absence of RNase 1 digestion using RT-qPCR < or =0.43 log10. This indicates that whereas the nucleic acid is still unmodified, the viral particles may no more be pathogenic. The scientists concluded that a gradual accumulation of phytochemicals overtime on the outermost part of the capsid might lead to a substantial decrease in cell culture infectivity. They found out that as a result of such accumulation of phytochemicals at the surfaces, the MNV particles increased in size owing to lemongrass oil and citral treatment. They concluded that time, other than the amount, was crucial for determining the effect of plant extracts [42].
The norovirus (NoV) belongs to the family of viruses called the Caliciviridae, which are non- enveloped, single-stranded, RNA positive sense viruses. The norovirus is purported to be the cause of about half of all gastroenteritis outbreaks world-wide and 75%-90% [4], of gastroenteritis not caused by bacteria in humans of all ages (3), (2). This includes about 23 million cases in the United States each year and an estimated 60-85% of all gastroenteritis outbreaks within the United States [5], Europe, and Japan.
Genome differentiation studies have placed the NoV into five separate genogroups (G1-GV), though a sixth genogroup was newly discovered [6]. The five genogroups were further subdivided into 44 genotypes made of 16 G1, 23 G11, 2 G111, 2 G1V, and 1 GV. The emergence of these genotypes was purportedly as a result of genetic recombination [3]. The human norovirus infection is most rampant in places where throngs of people gather, making easy spread likely. Symptoms include diarrhea, vomiting, and sometimes fever [5], although asymptomatic infection may occur [10]. It should be noted that GII.4 strains are responsible for the majority of outbreaks globally. Infection is through the fecal-oral path [7]. It’s been shown that there is a link between human genetic factors (FUT2 and HBGA) and norovirus susceptibility. In addition, the main reason for the emerging norovirus-gastroenteritis could be genetic alterations, which may include deletions, insertions, and multiplications of nucleotides, which may have contributed to the divergence from the virus evolutionary tree. Such manipulations will obviously result in single nucleotide polymorphisms (SNPs).
For countless years, man has explored the plant world in search of alternatives to orthodox medicine. Only about 1-10% of all plants have been used by human being in this regard [20], implying that lots of bioactive compounds (phytochemicals) are yet to be identified both in those plants that are familiar to humans and those yet to be investigated. It is on this premise that this review aims to discover or collate discoveries of plants and their phytochemicals capable of alleviating the human norovirus infection/gastroenteritis menace.
The phytochemicals of interest include flavonoids, phenols, terpenoids, anthraquinonoid, glycosides, and steroids in different plant extracts [22]. They have been used against bacteria, fungi, and viruses in various forms with promising results in many instances. Extraction methods included aqueous, petroleum ether, and ethanolic extractions. However, results are affected by the concentration of extract used and exposure time to the pathogen.
Understanding the pathogenesis of human noroviruses has not been easy because they cannot be cell cultured, but for the murine norovirus [37]. Most of what is known about its infection pattern is from observations made from human volunteers [8] and the tissue and biochemical studies carried out on them. In addition, some elucidations have been made using cow, pig and mouse models [9,37]. The use of advanced assays likes the RT-PCR, and methods which entail studying the virus at genetic and molecular levels have enhanced human understanding of the norovirus pathogenicity. The MNV model has been used to postulate many theories about the way the norovirus interacts with the host in order to cause infection. Both MNV and human (HuNoV) have been demonstrated to move across the interior of a cell. It has been documented that noroviruses infect immune cells, typically the macrophages and the antigen-presenting dendritic cells in-vitro (innate immunity), whereas they target the B cells in adaptive immunity. Also, the virus-like particles (VLPs) have been used to study the norovirus pathogenecity.
Comparisons made among different phytochemicals including zataria, clove, and oregano essential oils, as well as allspice oil, lemon grass oil and citral against the human norovirus surrogates (murine norovirus and feline calicivirus) were made. Significant results, which depended on the amount of extract and time of exposure, were obtained. These led to the conclusion that a gradual accumulation of phytochemicals overtime on the outermost part of the capsid may lead to a substantial decrease in cell culture infectivity. More studies are needed both on the noroviruses and the phytochemicals in an effort to find more endearing solutions to the human norovirus infection and gastroenteritis. Plant based chemicals are less toxic, have less or no side effects, and easy to formulate in food, the use of such phytochemicals should be expanded, which ultimately help to prevent the transmission of Norovirus. In the same time, it will improve the livelihood of the susceptible group of the people mainly infants and elderly.