Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2016) Volume 5, Issue 1
Keywords: Structure and function; Gene sequences; Tyrosinase; Browing inhibition; Antisense; GRAS inhibitor
Polyphenol oxidase (PPO) enzyme catalyzes the hydroxylation of monophenols to o-diphenols through monophenol oxidase activity and a subsequent oxidation of these o-diphenols to the corresponding o-quinones by a catecholase/diphenolase activity in the presence of its cosubstrate oxygen. Enzyme nomenclature differentiates the monophenol oxidase as laccase (EC 1.10.3.2) and the o-diphenol: oxygen oxidoreductase as catechol oxidase (EC 1.10.3.1). However, depending on the source of enzyme it may act on two general types of substrates, monophenols and diphenols, hence in general polyphenol oxidase (Tyrosinase; EC 1.14.18.1) name has still been in use in literature [1]. For example, tyrosinase from mushroom shows both monophenol oxidase and o-diphenol oxidase activity, whereas, the catechol oxidase from most plant sources shows only o-diphenol oxidase activity (Figure 1).
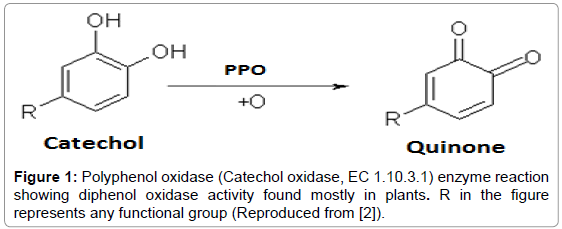
Figure 1: Polyphenol oxidase (Catechol oxidase, EC 1.10.3.1) enzyme reaction showing diphenol oxidase activity found mostly in plants. R in the figure represents any functional group (Reproduced from [2]).
The naming of this enzyme has not been consistent and many a times it has been parallely named on the basis of individual substrate upon which it has been found to work such as tyrosinase, phenolase, catechol oxidase, catecholase, o-diphenol oxidase, monophenol oxidase, and cresolase. The structure of the active site of the enzyme contains two copper ions, which are bound by six histidine residues and a single cysteine residue. The enzyme seems to be of almost universal distribution in animals, plants, fungi and bacteria. In plants this enzyme is required for defense against predators like herbivores and insects (Figure 2).
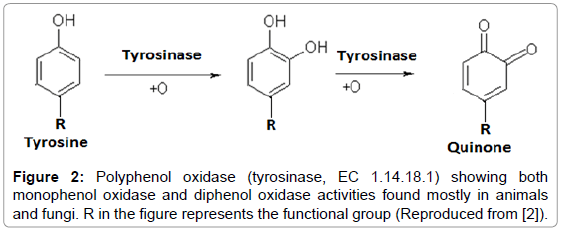
Figure 2: Polyphenol oxidase (tyrosinase, EC 1.14.18.1) showing both monophenol oxidase and diphenol oxidase activities found mostly in animals and fungi. R in the figure represents the functional group (Reproduced from [2]).
Although constitutively expressed in plants, PPO activity is often up-regulated in wounded tissues, and thus PPOs are often considered to be plant defense proteins [2,3]. Gene expression profiling in poplar (Liriodendron tulipifera) tissues and organs revealed that the PPO genes are all differentially expressed during normal development, but that only a small subset of PPO genes are significantly upregulated by wounding, methyl jasmonate or pathogen infection. However, its role in fungi and bacteria is not clear. This enzyme is responsible for undesirable enzymatic browning in plant products, seafoods, and melanin formation in human skin [4]. Therefore, it has received significant attention from researchers working in area of food science, plant physiology, and cosmetics development. Though in plants it is considered to be related for defense, not much is known about its exact biological functions. In plant cell, PPO and its principal substrate (phenolics) were reported to be present in chloroplast and vacuoles, respectively [5]. During minimal processing involving abrasion and cutting, cellular structures get disrupted leading to the contact of PPO with its substrate. In the presence of oxygen, the enzymatic reaction results in the formation of quinone which autopolymerises to produce melanin like brown pigment resulting in browning [6]. This postprocessing browning is a major problem for food industries dealing with cut fruits and vegetables, juices, shrimps and related products. The brown coloration basically causes loss in the eye appeal of these products. Therefore, the control of browning by inhibiting PPO activity has remained an exciting and challenging area.
In general PPO oxidizes phenolic substrates in the presence of oxygen. The active site of PPO undergo transitions among met-, oxy- , and deoxy-forms in a cyclic manner (Figure 3) and in each cycle two molecules of catechol are oxidized and one molecular oxygen is reduced to water resulting in the formation of two quinone products [7]. The dioxygen (O2) binds to the copper metal center of the enzyme replacing the solvent molecule (H2O) bonded to CuA in the reduced enzyme form (Deoxy or reduced form). UV/Vis spectroscopy suggests that molecular oxygen binds first as a peroxide which is followed by the binding of the catechol substrate [7]. The catechol molecule undergoes deprotonation of one of the two hydroxyl groups and binds to CuB (Oxy-form). The transfer of two electrons from the substrate to the peroxide is followed by protonation of the peroxide group and cleavage of the O-O bond (Figure 3). Glutamate (at position 236) and the second non-coordinating hydroxyl group of the substrate donate a proton and promote the loss of water and the formation of the o-quinone product. Protonation of the bridging group by solvent brings the active site into the resting hydroxide-bridged dicupric state (met-form). Another molecule of catechol can then serve as ‘co-substrate’ reducing the hydroxide-bridged dicupric (CuII) state back to the dicuprous form (CuI). The dicuprous state Cu(II)-Cu(II) of active site again repeat the catalytic raction cycle [7] (Figure 3).
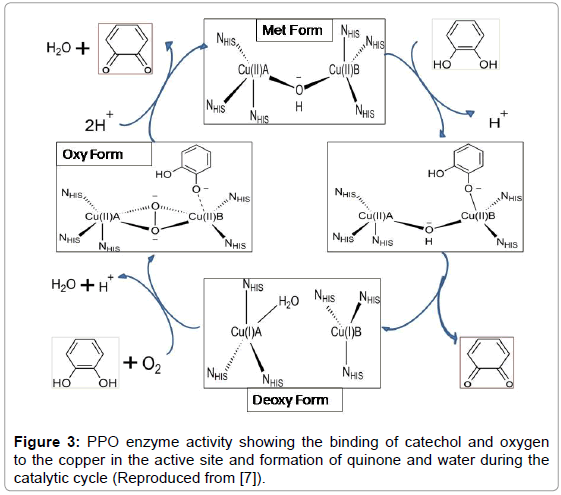
Figure 3: PPO enzyme activity showing the binding of catechol and oxygen to the copper in the active site and formation of quinone and water during the catalytic cycle (Reproduced from [7]).
Purification
The purification of PPO from many plants to the homogeneity has remained difficult possibly due to the high phenolic content and irreversible binding of phenolics to PPO during purification steps. Roudsaria et al. [8] reported only 15 fold partial purification using ion exchange and gel filtration chromatography. In potato, the PPO was 5 fold purified with 18% yield, whereas, in banana peel the PPO was 460 fold purified with 2.2% yield [9,10]. In loquat fruit the PPO was 422 fold purified with about 35% yield [11]. One isoform of native PPO was 259 fold purified from eggplant and found to be a homodimer of 112 kDa [12]. The enzyme showed very low Km (0.34 mM) and high catalytic efficiency (3.3 × 106) with 4-methyl catechol. The substrate specificity was of the order of 4-methyl catechol > tert-butylcatechol > dihydrocaffeic acid > pyrocatechol.
These variations in fold purification and yield could be due to differences in the concentration and types of phenolics affecting their overall binding to the PPO protein which has been found to interfere with protein purification in certain fruits and vegetables [2]. The ionic and hydrophobic characteristics of the enzyme change upon binding with phenolics resulting in variation in the elution behavior during purification steps [2]. The phenol binding agents like polyethylene glycol, polyamide, or polyvinyl pyrrolidone are sometimes used to inhibit the binding of phenolics to the enzyme during extraction [1]. However, some of these chemicals inhibit the enzyme irreversibly. The PPO existing in membrane bound form is insoluble in extraction solution. In such cases, solubility is increased by the addition of detergents or by use of the cold acetone.
Substrate specifity
The plant PPOs are usually characterized for their substrate specificity as depicted here in the case of eggplant using eleven well known PPO substrates [12]. In this study 4-methyl catechol has shown the maximum activity (Table 1).
 |
Functional group (R) at meta position of -OH group | Relative Activity*(%) |
|---|---|---|
| Substrate | ||
| p-Cresol | Nil, (-CH3 at para position) | 0 |
| Gallic acid | -COOH (and -OH at ortho) | 1.5h ± 0.3 |
| 3,4-dihydroxybenzaldehyde | -CHO | 5h ± 1 |
| Pyrogallol | -OH at ortho position | 17g ± 2 |
| Chlorogenic acid | 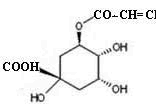 |
31e ± 2 |
| Caffeic acid | -CH=CH-COOH | 32e ± 2 |
| D-DOPA, L-DOPA | -CH2-CH(NH2)-COOH | 40d ± 2, 22f ± 2 |
| Pyrocatachol | Nil | 70c ± 3 |
| Pyrocatachol | Nil | 70c ± 3 |
| tert- butylcatechol | -C(CH3) | 83b ± 4 |
| 4-Methylcatechol | -CH3 | 100a ± 0 |
| * Relative activity was expressed with respect to 4-methyl catechol considering it as 100%. The values were expressed as mean ± SD of three parallel experiments. The values in column with different letter superscript are significantly different (P≤0.05). #Reproduced from [12]. | ||
Table 1: Substrate specificity of PPO purified from eggplant#.
Higher enzyme activity was observed with substrates having catechol ring structure and a functional group (-R) at meta-position with respect to -OH group. A smaller functional group (-CH3) present in 4-methyl catechol probably rendered it with the maximum activity. With an increase in the size of the functional group (-R) the activity decreased as was observed in the case of tert-butylcatechol, chlorogenic acid, and DL-DOPA (Table 1). On the contrary, the absence of the functional group (-R) also decreased activity by 30% as observed in the case of pyrocatechol. These observations indicated the important role of the -CH3 group and its location in the structure of catechol in determining its specificity. The -R group position in the catechol ring also observed to be determinant of higher specificity. The active site of the enzyme could be interacting with these substrate molecules where -R group position and composition provided advantage for efficient catalysis. Most of the substrates with catechol ring structure and a functional group at other position of the ring (-OH at ortho position as in pyrogallol, gallic acid, and -CH3 at para position as in p-cresol) showed very less or absence of activity [12].
Copper as cofactor
Copper present in PPO is responsible for the oxidation-reduction processes that involve electron transfer, dioxygen chemistry, and reduction of nitrogen oxides. The metal cofactors are usually not covalently bound to the rest of the protein molecule, but are kept in place by interaction of amino acid side chains that have unpaired electrons (histidine, cysteine) with empty axial orbitals of the transition metal cofactor. Copper exists mainly in two oxidation states, Cu(I) and Cu(II), and often changes between these two states while catalyzing reactions. It can also, like other transition metals, serve as a structural support for the larger protein molecule that it is a part of, thereby affecting activity of a protein.
Enzyme structure
Many PPOs are believed to function in vivo as monomers, but a few fungal PPOs may associate to function as small oligomers (e.g. tetramers). The subunit molecular masses reported in the literature for plant and fungal PPOs vary considerably, even within a species. The monomers have independent active site and are enzymatically active, but for structural stability they often exist as oligomers. For a few species lower molecular mass subunits have been shown to result from proteolysis of a larger precursor form. The crystal structure of PPO from sweet potato (Ipomoea batatas) [13] and grapes (greenache; Vitis vinifera) [14] has been resolved and published (Protein Data Bank, PDB).
From sweet potato three structures are published, out of which two are in native Cu(I)-Cu(I) state (PDB accession no. 1BT1, 1BT3), one is in reduced Cu(II)-Cu(II) state (1BT2) and the other in complex with phenylthiourea (PDB accession no. 1BUG). In fungi two structures of mushroom (Agaricus bisporus) tyrosinase are published, both are in deoxy state, contain an unknown lectin-like subunit and bind to inhibitor tropolone [15]. The search of PDB for tyrosinase revealed bacterial PPO sources like Streptomyces castaneoglobisporus, Citrobacter freundii, and Bacillus megaterium. From Streptomyces as many as 22 different structures are in PDB including the tyrosinase caddie protein complex differently soaked in copper solution (8 nos. starting from 3AWS-Z and also 3AXO), in deoxy-from (4 nos. from 2ZWD-G, and 2ZMZ, 2AHL), met-form (2ZMX, 2ZMY), and oxy-form (1WX4, 1WX5) [16]. These structures are also published in copper free form (1WXC, 1WX5). These studies explained the flexible nature of the dinuclear copper centre in the active site during catalysis, which can accommodate both the monophenolase and diphenol oxidase activity. On the other hand the catechol oxidase from other sources including plants has rigid active site and do not show the monophenolase activity. In other study presence of six structures of PPO from Bacillus megaterium in the presence and absence of zinc ion and kojic acid inhibitor explained the plasticity of copper binding indicating the variation in the copper ion position within the active site [17]. The histidine residue position is also shown to play a role in accessibility of substrate to the active site of enzyme.
The PPO gene is transcribed to form the PPO mRNA which is translated to the precursor PPO protein inside cell. The PPO gene and its transcript have the following features.
Gene and protein sequences
The list of plants in which PPO has been sequenced and at least partly characterized is growing steadily. The nucleotide sequence of PPO is available in many plants including eggplant, potato, tomato, tobacco, sweet potato and cereals like wheat, and legumes like pea. In most of the dicot plants including tomato and potato the introns are reported to be absent. Monocots like banana and wheat, which have at least four and six distinct o-diphenol oxidase genes, respectively, show the presence of intervening sequences [18]. In pineapple (Ananas comosus) sequence analyses showed that both PPO genes contained single intron [19]. The presence of two introns was also reported in barley (Hordeum vulgare) PPO [20].
ClustalW sequence alignment of PPO nucleotide sequences showed considerable homology among close family relatives (Figure 4). The homology is limited among the phylogenetically distantly placed plants [21]. For example, potato nucleotide sequence (Genbank accession U22921.1) showed 89% homology with tomato (Z12836.1), whereas, the homology was 77% with relatively distant relative persimmon (Diospyros kaki) PPO (AF039165.1). PPOs of closely related plants, such as potato (AAA85122.1) and tomato (AAB22610.1) showed approximately 74% homology in the amino acid sequence. However, potato showed only 43, 44, and 42% homology with cotton, tea, and grape, respectively (Figure 4). Though the sequence length varies in different plants, in general the nucleotide sequence of PPO gene was about 1800 bp long with the translated PPO precursor protein about 600 amino acid long with a molecular weight of about 66 kDa.
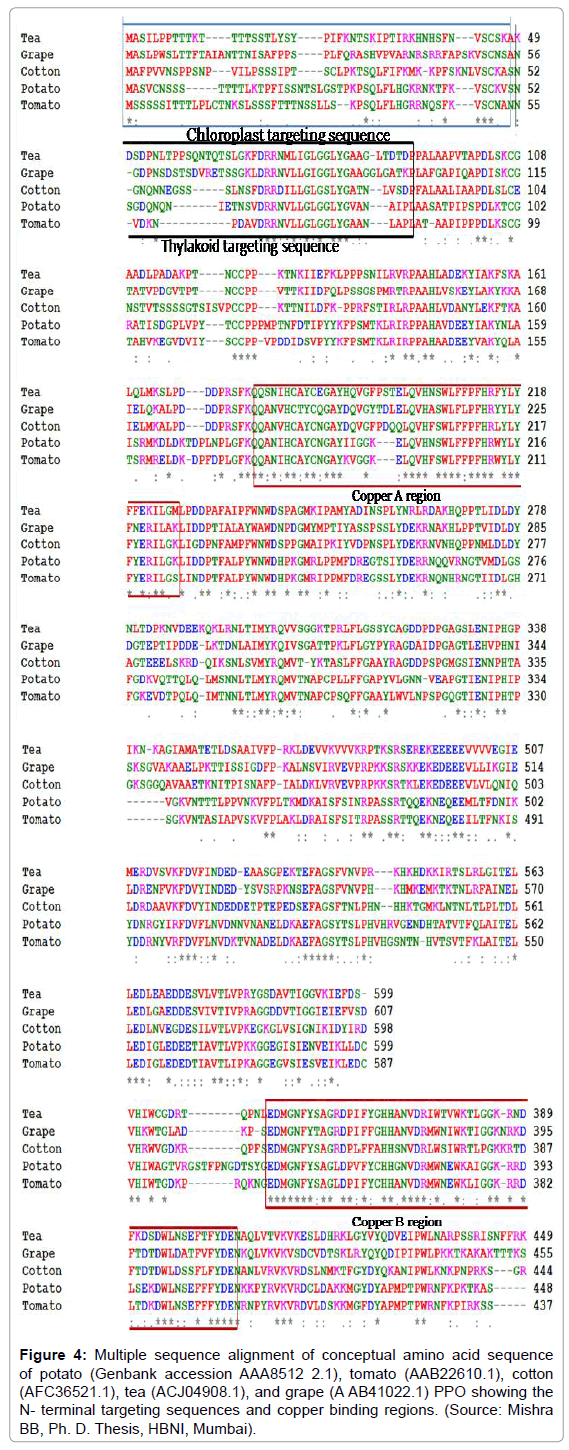
Figure 4: Multiple sequence alignment of conceptual amino acid sequence of potato (Genbank accession AAA8512 2.1), tomato (AAB22610.1), cotton (AFC36521.1), tea (ACJ04908.1), and grape (A AB41022.1) PPO showing the N- terminal targeting sequences and copper binding regions. (Source: Mishra BB, Ph. D. Thesis, HBNI, Mumbai).
The protein sequence of PPO can be classified into three regions. The N-terminal is the chloroplast targeting sequence region and spans about 80 amino acids. The next is the copper binding region (dinuclear copper centre) which again divided into Cu A (170-220 aa) and Cu B (350-410 aa) region. The remaining part of the sequence up to about 600 residues constitutes the C-terminal region. The comparative analysis of PPO protein sequence in plants and fungi has been reported [22], where a number of important structural features were found to be conserved in the N-terminal domains including a tyrosine motif which can be considered as a landmark indicating the beginning of the linker region connecting the N-and C-terminal domains. All species possess the six histidine residues that associate with the two copper ions of the active site. The first three histidine residues encompass a region termed the CuA site, and the following three encompass the CuB site. An unusual thioether bridge was identified between a cysteine residue and the second histidine of the CuA site. This covalent bridge appears to have a structural role in which it puts additional structural restraints on the CuA center. In sweet potato (Ipomoea batatas) PPO, the cysteine involved in the thioether bridge is located four residues after the first histidine of the CuA site. Due to the conservation of these cysteines, it is likely that all plant and fungal PPOs have a thioether bridge as a structural component of their CuA sites [22]. The Phenylalanine (at 261 position) of sweet potato PPO has been termed the gate residue because of its location above the active site, partially blocking the substrate entrance. The bulky phenylalanine side chain must rotate to allow substrate binding to occur. Other plants either have phenylalanine or leucine at this position, whereas, fungal PPO have leucine or proline at this position. Immediately after the CuB region, a tyrosine motif (Y/ FxY) is located (at 331-333 amino acid position in sweet potato PPO). The hydroxyl group of tyrosine side chain form hydrogen bonds to an aspartate side chain located four residues after the third histidine of the CuB site. This aspartate (at 288 position in sweet potato PPO) is reported to be conserved in all plant and fungal PPOs [22].
The glycosylation sites could not be predicted with confidence for plant and fungal polyphenol oxidases. However, such sites were recently predicted in a cultivar of eggplant [18]. The primary sequence of the linker region is much longer for plant and fungal PPOs, ranging from 52 (in rice), 80 residues in potato and faba bean, and 59-98 residues for fungi [22].
Gene families
PPO gene families have been described in several species, with individual PPO genes typically showing distinct expression patterns. They have been most extensively investigated in wheat (6 genes), potato (5 genes), tomato (7 genes), eggplant (5 genes), poplar (9 genes), faba bean, red clover (6 genes), banana (4 genes) and apple (2 genes) [18,23-26]. These genes were isolated from diffrenet parts of the plant such as fruit, flowers, and leaves [27-29]. They have also shown the presence of intervening sequences [30,31]. Although genes homologous to o-diphenol oxidase are absent in Arabidopsis genome, laccases (17 genes) are present [32]. Hexaploid wheat kernels have six genes coding for PPO, of which at least three are expressed during development of the kernel. These six genes fall in two clusters with three similar sequences in each.
Chromosomal location
In wheat (Triticum aestivum) the genetic mapping of PPO exclusively performed and has implications for selection for wheat variety with low PPO activity [33]. The Quantitative Trait Loci (QTL) for PPO in T. aestivum indicated that number of PPO genes are present on different chromosomes. One gene from of T. turgidum coding for PPO was mapped to chromosome 2D [33]. In Barley the PPO or phenol reaction (Phr) gene has been mapped to the long arm of barley chromosome 2H [20]. In grape variety (Pinot nori) the search of the complete genome showed the presence of four homologues on chromosome 10 [14]. The search for PPO gene in Solanum Genomics Network (SGN) resulted in the finding of the physical locus of PPO gene on chromosome number 8 both in potato and tomato.
Further, in comparison to tomato chromosome 8, the chromosomal rearrangements (inversions and translocations) are absent in eggplant which indicated the similar localization of PPO.
In-vivo expression profile
The presence of multiple genes of PPO and its differential expression in different parts of plant and at different stages of development is one of the most important phenomena reported [24]. Differential tissue specific expression of six genes coding for PPO in potato, and seven genes in different tissues of tomatoe has been reported [24]. Apple PPO is encoded by a multiple gene family, whose expression is up-regulated by wounding of the tissue. Two different genes are expressed at different stages of apple flower development, one gene coding for PPO being expressed only at the post-anthesis stage [34]. There are few evidences that implicate PPO as defence genes possessing both constitutive and inducible modes of expression and having multiple functions. The constitutively abundant PPO in glandular trichomes of Lycopersicon spp. and Solanum spp. is reported to be responsible for entrapment of small-bodied insects via oxidative polymerization of trichome exudate. Systemic induction of PPO expression might act as an additional line of defense to protect the growing parts of plants against further attack by pathogens and insects [29]. Recently, it has been shown that antisense down regulation of constitutive and inducible PPO activity resulted in pathogen hypersusceptibility, suggesting that PPO may possess a key role in plant defence systems. Vickers et al. [35] manipulated the levels of PPO in transgenic sugarcane using constructs of sense and antisense to the native PPO gene and found change in the degree of browning.
In-vitro activation
Different injuries, salicylic acid, ethylene, and jasmonates elicited developmental stage specific patterns of PPO expression. Fungal PPO as plant PPO, can be present in latent form which is activated by SDS or proteolysis or acid shock. SDS activation of PPO in beet root is reported to be reversible [5]. Further insight into the activation of a latent PPO comes from the work of Kanade et al. [36] on Dolichos lablab, which shows that both acid and SDS change the environment of a single glutamic residue, close to the di-copper active site. As a result the active site is unblocked or opened and enzyme activity enhanced. Similarly wounding or methyl jasmonate could cause localized acidification which results in conversion of a latent to an active enzyme.
Transcriptional regulation
The promoter studies of PPO have been reported in tomato [29] and pineapple [19]. In pineapple southern blot analyses suggested that pineapple contained two PPO genes, PINPPO1 and PINPPO2. Analysis of expression of PINPPO1 promoter GUS fusion constructs showed this promoter had a low basal activity and was cold and wound inducible. Striking homologies to gibberellin response complexes (GARC) were observed in sequences of both the PINPPO1 and PINPPO2 promoters, which indicated these promoters respond gibberellins. In tomato, analysis of PPO promoters as GUS fusion constructs showed that mechanical wounding and infection by fungal and bacterial pathogens induced transcription of one isozyme of PPO (PPOf). The PPO transcript levels systemically increased in young leaves when mature leaflets were injured. It was demonstrated in their study that cis-elements are sufficient for PPO induction and reside in the 5’ flanking region. Again Newman et al. [23] reported similar results in another isozyme of tomato PPO (PPOB). The PPOB promoter was sequenced (1975 bp 5’ upstream) and subjected to homology analysis. Sequence similarity of this 5’ upstream sequence was observed with nucleotide sequences of genes encoding enzymes/proteins active in phenylpropanoid biosynthesis, signal transduction and responsiveness to hormones and stresses. The putative cis-elements were also reported. Chimeric gene fusions were constructed linking PPOB 5’ regions to the reporter gene, β-glucuronidase (GUS). The resultant transgenic plants were analyzed for GUS activity in various vegetative and reproductive tissues. It was shown that PPO B expression was tissue specific, developmentally regulated, and localized to mitotic or apoptotic tissues. In barley (Hordeum vulgare), two PPO enzymes were identified and their expression was differentially expressed in seed coat [37]. An insertion of a hAT-family transposon in the promoter region of PPO2 was indicated to be responsible for different expression patterns of the duplicate PPO genes in barley.
Posttranslational modification
The synthesis of PPO and its transport to its site in chloroplasts, where plant PPOs is thought to be located, is a complex process, but it has the general features of import of nuclear coded proteins into sub-cellular organelles. The PPO is translated as a precursor protein and undergoes N-terminal and C-terminal processing along with phoshorylation, N-myristoylation, and O-glycosylation sites. In greenache grapes amino acid sequence of leaf PPO was deduced together with predicted sites of posttranslational modification [14]. Possible phosphorylation sites are abundant and consistent with some reports on phosphorylation of PPOs. However, the mature active form that was crystallized was devoid of phosphorylation and glycosylation. The occurrence of six potential myristoylation sites in this grape was also predicted which indicated the probable association with membrane [14].
Targeting of PPO and N and C-terminal processing
The synthesis of PPO and transport to its site in chloroplasts is a complex process, but it has the general features of import of nuclear coded proteins into sub-cellular organelles. Plant PPOs contain a bipartite N-terminal plastidic transit peptide directing the protein to the plastid lumen. This N-terminal transit peptide is cleaved off in two steps to generate the mature protein. The pathway by which plant PPO reaches the chloroplast was investigated by Sommer et al., [37]. Using an in vitro system it was found that tomato PPO, coded by cDNA, was processed in pea chloroplasts in two steps during its import. The precursor PPO with Mw, 67 kDa was imported into the stroma of the chloroplasts by an ATP-dependent step. It was then processed into a 62 kDa form by a stroma peptidase. Subsequent transport into the lumen was light dependent and resulted in the mature 59 kDa form. Apparently, such processing is a feature of all chloroplast located PPOs. The precursor protein contains a transit peptide, which must be removed in order that the PPO reaches its site in the chloroplast. The processing is carried out by a stromal peptidase [38]. The peptide bond cleaved by the processing proteases was identified as alanine-serine or alanine-alanine in most plant PPO examined so far. The transit peptide is characterized by the presence of three domains: The N-terminal 25 amino acids contain many hydroxyl containing amino acids, the middle part is more loosely conserved and called the ‘n’-region, while the C-terminal 25 amino acids form a hydrophobic domain, which is the thylakoid transfer domain (lumen targeting). The import and processing did not require Cu2+, but import was inhibited by micromolar concentrations of Cu2+. Further studies revealed that this inhibition was probably due to this inhibition of the stromal peptidase involved in the processing of the precursor protein [37]. N-terminal protein sequencing by Edman degradation of the mature protein has established the transit peptide cleavage site for PPO from sweet potato (I. Batatas), tomato (L. esculentum; PPOE), potato (S. tuberosum; POT32) tubers, spinach (S. oleracea) thylakoid, wheat (Triticum aestivum), bean (V. Faba), and grape (V. vinifera). The transit peptide cleavage site for many other plants has been predicted based on homology.
The location of fungal PPO is not entirely clear. Generally it appears to be a cytoplasmic enzyme. The products of fungal and bacterial genes lack the N-terminal domain, since these organisms also lack the plastids to which the plant PPO is targeted. However, a PPO from Pycnoporus over produced in Aspergillus niger, could be targeted to the extracellular growth medium [39]. An additional PPO is present in the mycelium of Pycnoporus saguineus, which has a very high tyrosinase activity and is able to convert coumaric acid to caffeic acid in vitro [39]. An interesting feature of tyrosinase from Agaricus is that it contains putative glycosylation and phosphorylation sites, although no glycosylation or phosphorylation has so far been reported for fungal PPO [40]. Although fungal PPOs do not have transit peptides, two reports indicated that PPO from some fungi do undergo N-terminal proteolysis of a few residues [40]. N-terminal protein sequencing of the active enzymes from Pycnoporus sanguineus and Trichoderma reesei revealed that 4 and 18 residues had been cleaved from their N-termini, respectively. In the case of T. reesei PPO, these 18 residues constitute a signal peptide. Unlike all fungal PPO studied to date, T. Reesei PPO is excreted. On the other hand, N-terminal protein sequencing of Neurospora crassa PPO indicated that this protein did not undergo any N-terminal proteolysis except the initial methionine.
Most of the PPOs that have been studied are known to undergo significant processing to cleave the C-terminal domain, and gene sequences suggested the existence of similar processing sites in a host of related proteins. The C-terminal cleavage is thought to be carried out by enzymes in the thylakoid lumen. The C-terminal processing takes place in the linker sequence (WLPKNTKAKAK), which begins shortly after a tyrosine motif (YQY). A proline residue in this linker sequence was found to be the actual site of proteolysis [14]. For V. faba and V. vinifera PPOs the 40 kDa active form has been shown to result from proteolytic cleavage of a C-terminal fragment from the 60 kDa latent form [40]. C-terminal processing occurs for the PPO from the sweet potato (I. batatas) as well. The crystal structure has been determined for an active 39 kDa form of I. batatas PPO which is missing a C-terminal fragment, apparently as a result of in vivo proteolysis. Similarly, the 75 kDa precursor of tyrosinase from the ascomycete N. crassa is C-terminally processed to a 46 kDa active form [40].
In potato (Solanum tuberosum) and pokeweed (Phytolacca americana) two PPO cDNAs clones from leaves were reported which were used for nucleotide sequence characterization [27]). In grape berry the PPO cDNA clone was constructed from the sequence of two overlapping cDNA clones using 5’ and 3’-RACE (Rapid Amplification of cDNA Ends) PCR [41]. In case of apple (Malus pumila) PPO gene was expressed in E. coli, and the gene product (56 kDa) without a transit peptide was immunochemically detected and was of the same size (ca. 65 kDa) as the main PPO of apple fruit determined by SDS-PAGE [42]. The DNA coding region for one of the PPOs from apple fruit was cloned and expressed in E. coli. The PPO contained a transit protein and was processed to a mature PPO, Mw 56 kDa. Although the protein expressed in E. coli, Mw 56 kDa, was detected using antibodies, the gene product was enzymically inactive [42]. Molecular cloning and characterization of PPO gene has been reported in apricot (Prunus armeniaca) fruit [43], hybrid poplar (Populus trichocarpa × Populus deltoides) plant [3] and red clover (Trifolium pretense) leaves [26]. However, the overexpression studies in E.coli were not reported. In poplar the PPO cDNA clone was isolated from cDNA library and the probe was used in northern hybridization to study the expression of PPO in response to wounding and herbivory. In red clover grass leaves PPO cDNA was prepared, cloned and expressed in E. coli. However, the expressed protein in E. coli formed insoluble inclusion bodies. This PPO gene when expressed in alfa alfa (Medicago sativa) surprisingly showed significant enzyme activity. In tomato, overexpression of polyphenol oxidase in transgenic tomato plants resulted in enhanced bacterial disease resistance [44,45]. In order to investigate the role of PPO in plant disease resistance, transgenic tomato (Lycopersicon esculentum) plants that overexpressed a potato cDNA under control of the cauliflower mosaic virus 35S promoter. The transgenic plants expressed up to 30-fold increase in PPO transcripts and 10 fold increases in PPO activity and immunodetectable PPO. Arican and Gozukirmizi [46] reported reduced PPO activity in transgenic potato plants associated with reduced wound inducible browning phenotypes. Wounded transgenic plants with reduced PPO activity exhibited a great amount of reduced browning when compared with control plants. Wang and Constabel [46] studied the defensive role of leaf PPO in transgenic poplar plants overexpressing PPO from another variety. The transgenic plants showed high PPO enzyme activity and resistance to forest tent caterpillar larvae. Thipyapong et al. [29] reviewed the experiments in which the roles of PPO in disease and insect resistance were investigated using transgenic tomato (Lycopersicon esculentum) plants with modified PPO expression levels (suppressed PPO and overexpressing PPO). These transgenic plants showed normal growth, development and reproduction under laboratory conditions. Antisense PPO expression dramatically increased susceptibility while PPO overexpression increased resistance of tomato plants. The PPO gene from cultivar ‘Kalpatharu’ of eggplant was cloned using PET 28a expression vector downstream of lac I promoter (IPTG inducible) in E.coli (BL-21). The IPTG induction showed overexpression of a protein of 66 kDa size, observed to form inclusion body. This insoluble PPO was solubilized and purified but showed very poor enzyme activity, which could be attributed to the absence of post translational modifications in E. coli (Mishra, BB, PhD thesis HBNI, Mumbai).
Inhibition of PPO and enzymatic browning
Many fruits and vegetables including potato, sweet potato, breadfruit, jackfruit, yam, mushroom, apple, avocado, banana, grape, peach are prone to postharvest and processing induced browning and therefore causes considerable economic losses [6]. The control of browning from harvest of the vegetable to the consumer is therefore very critical for minimizing losses and maintaining economic value. Browning also adversely affects flavour and nutritional value. The phenomenon of enzymatic browning and the approaches used for its prevention have been elaborately discussed by Marshall et al. [47]. Various methods have been developed for inhibiting or preventing PPO activity over the years for the control of this undesirable enzyme activity. These techniques eliminate one or more of the essential components (oxygen, enzyme, copper, or substrate) required for PPO reaction.
a) Browning of the cut fruit occurs rapidly upon exposure to oxygen. The elimination of oxygen from the cut surface of the vegetables prevents the browning reaction, which is also possible by immersion in water, syrup, brine, or by vacuum treatment [47].
b) Copper is present in the active site of PPO enzyme. Elimination of copper by using chelating agents makes PPO inactive and browning can be prevented [6].
c) Inactivation PPO by physical treatments (high pressure, steam blanching) is used for the control of browning in canned or frozen fruits and vegetables. However, heat treatments are not practically applicable in the storage of fresh produce [47].
d) PPO catalyses the oxidation of phenolic substrates such as caffeic acid, protocatechuic acid, chlorogenic acid, and tyrosine. Chemical modification of these substrates can prevent their oxidation and browning [48].
e) Certain chemical compounds react with the products of PPO activity and inhibit the formation of the melanin like pigments resulting in inhibition of browning [6].
Physical methods
Different kinds of physical treatments have been reported to control enzymatic browning in different fruits and vegetables. Among this heat treatment is the most widely utilized method for stabilizing foods because of its capacity to destroy microorganisms and to inactivate enzymes. Low temperature can also inhibit browning by inhibiting PPO activity [6]. However, fruits like banana undergo chilling injury mediated browning under refrigerated storage. Low dose gamma radiation also reported to inhibit browning in potato tubers and tropical fruits [49]. High pressure treatment has also been reported to inactivate PPO in apple, grape, avocado and pear at room temperature (25 °C) at 600, 700, 800 and 900 Mpa, respectively [50]. Ultrafiltration (UF) is used for white wines and other fruit juices as it removes PPO [51]. UF was used for improving the colour stability of banana juice without the application of heat, which is known to alter its flavor [52].
The mechanics of cutting and further processing were found to have very profound effect on the browning process. A method was reported where fine cutting (blade thickness, 0.04 mm) and immediate washing in water inhibited browning in fresh cut ready-to-cook (RTC) eggplant [52-54]. The scanning electron and fluorescence microscopic examinations showed that fine cutting caused lesser physical injury and cellular death, resulting in reduced leaching of phenolics and PPO activity and hence lesser browning. For commercial acceptability of the technique, storage studies indicated that fine cut samples could be stored up to 5, 12, and 16 days at ambient (26 ± 2 ), 10 and 4°C, respectively and found to be microbiologically and organoleptically acceptable [53].
Chemical methods
The use of browning inhibitors in food processing is restricted due to concerns like toxicity, effect on taste, flavour, and texture [48]. Browning inhibitors may be classified in accordance with their primary mode of action. PPO inhibitors could be grouped in six catagories based their mode of inhibition of enzymatic browning (Table 2).
| Reducing agents/antioxidants | Sulfiting agents, ascorbic acid and analogs, cysteine, glutathione, phenolics |
|---|---|
| Chelating agents | Phosphates, EDTA, organic acids |
| Acidulants | Citric acid, phosphoric acid |
| Enzyme inhibitors | Aromatic carboxylic acids, aliphatic alcohol, anions, peptides, substituted resorcinols |
| Enzyme treatments | Oxygenases, o-Methyl transferase, proteases |
| Complexing agents | Cyclodextrins |
| Transgenic approach | Introduction of antisense gene |
| #Reproducd from [56] |
Table 2: Approaches to inhibit post-cut enzymatic browning in fruits and vegetables#.
Reducing agents/Antioxidants: Reducing agents prevent enzymatic browning either by reducing o-quinones to colourless diphenols, or by reacting irreversibly with o-quinones to form stable colourless products. Such compounds are effective in controling browning and include sulfiting agents, ascorbic acid and analogs, cysteine, and glutathione [5].
Regulatory restrictions are applied globally to the use of sulfites in foods since sulfite allergies pose a health risk. According to the United States Food and Drug Administration (US FDA) Federal Register foods containing ≥10 ppm of sulfiting agent must declare the content on the ingredient label [47]. As a result, there has been a considerable focus on identifying appropriate sulfite substitutes for use in foods. Ascorbic acid is a reducing compound and is acidic in nature, forms neutral salts with bases, and is highly water-soluble. L-ascorbic acid and its various neutral salts and other derivatives have GRAS (Generally Recognized as Safe) status and now in use for prevention of browning in juices, fruits and vegetables. It inhibits browning by reducing the o-quinones to their precursor diphenol substrates [55,56]. Cysteine is an effective inhibitor of enzymatic browning and is more effective than sodium bisulfite as an antibrowning agent [57]. Cysteine hydrochloride, potassium metabilsulfite, ascorbic acid, erythorbic acid, resorcylic acid and kojic acid were compared for their PPO inhibition activity in eggplant, which showed that cysteine hydrochloride was found to be the most effective inhibitor with a low inhibitor constant of 1.8 μM [12]. In addition phenolic antioxidants are also reported to preserve food by retarding oxidative changes thereby preventing deterioration, rancidity or discolouration. Antioxidants inhibit oxidative processes by reacting with free radicals, through metal chelation, and by scavenging singlet oxygen [6]. Both synthetic and naturally occurring phenolic antioxidants are used in food applications. Several synthetic antioxidants, such as butylated hydroxytoluene (BHT), tertiarybutyl hydroxyquinone (TBHQ) are permitted for use in food.
Chelating agents: PPO enzyme has copper metal ions at its active site. Chelating of copper ions can inactivate the enzyme. Such agents used in the food industry include sorbic acid, polycarboxylic acids (citric, malic, tartaric, oxalic, and succinic acids), polyphosphates (ATP and pyrophosphates), macromolecules (porphyrins, proteins), and EDTA [55].
Acidulants: Acidulants are generally applied in order to maintain the pH well below that required for optimum catalytic activity of an enzyme. Acidulants such as citric, malic, and phosphoric acids are capable of lowering the pH of a system, thus rendering PPO inactive [58]. Ionizable functional groups of the enzyme structure are affected by the pH of the food medium. These groups must be in appropriate ionic form in order to maintain the conformation to catalyse the enzymatic reaction [6]. Changes in the ionization status of enzymes are generally reversible. Citric acid is the one of the most widely used acidulants in the food industry. It is typically applied at levels ranging between 0.5 and 2 percent (w/v) for the prevention of browning in fruits and vegetables. In addition, it is often used in combination with other antibrowning agents such as ascorbic or erythorbic acids and their neutral salts. Citric acid inhibits PPO by lowering the pH as well as by chelating the copper at the active site of the enzyme [59].
Enzyme inhibitors: Many specific inhibitors of PPO enzyme have been reported in literature. Four-hexylresorcinol (4-HR) has been used in pharmaceuticals and is considered to be safe and effective in use as an anti-browning agent [60]. It has several advantages over sulfites when applied in the control of browning in foods. Four-hexylresorcinol was reported to control browning in fresh and hot-air dried apple slices as well as in apple juice [58]. Several studies have shown the effectiveness of 4-HR in controlling enzymatic browning in shrimp, mushroom and apple slices [47]. Four-hexylresorcinol (HR) is a chemically stable, water-soluble compound. Toxicological, mutagenic, carcinogenic, and allergenic studies have shown that there are no risks associated with the levels of 4-HR used in the treatment of shrimp (Mayer, 2006). Fourhexylresorcinol has been given GRAS status from the United States Food and Drug Administration, for use on shrimp [60].
Halide salts: Inorganic halides are known to inhibit PPO, for example NaF is the most potent inhibitor of apple PPO, followed by NaCl, NaBr, and NaI [6]. The inhibition of enzymatic browning by halides decreases with increase in pH. Sodium chloride and calcium chloride at concentrations of ranging between 2 and 4 percent (w/v) are most commonly used in the food industry for the inhibition of browning [47]. Amino acids, peptides and proteins inhibit PPO either by inhibition of the enzyme or by reacting with the product quinones [55]. Proteins, peptides and amino acids are capable of forming stable complexes with Cu2+. In addition, they are also capable of chelating copper at the active site of PPO. Histidine and cysteine have high affinities for Cu2+. Imidazole ring of histidine and thiol group of cysteine, both have metal binding capacity [48]. Aromatic carboxylic acids such as benzoic acid and cinnamic acid are structurally similar to phenolic substrates and known to inhibit PPO. These acids inhibit PPO by forming complex with copper at the active site. Cinnamic acid at levels of 0.01 percent was observed to be effective in long term inhibition of PPO in apple juice [56]. Benzoic acid and its derivatives showed inhibitory effect on PPO activity in mushrooms [61].
Complexing agents
Cyclodextrins and Chitosan: The cyclodextrins (CD) are highly insoluble and inhibits browning of fruit juice by binding with PPO substrates. Polyphenols can be removed by CD and by insoluble polyvinyl polypyrrolidone or polyethylene glycol [2]. The adsorption of flavour or colour compounds by cyclodextrins poses a major drawback to their use in food systems. The applicability of cyclodextrins in fruit and vegetable juices has been patented [47]. However, cyclodextrins have not yet been approved for food use by the United States FDA. Edible Coatings minimize undesirable changes due to minimal processing and has been reported for several commodities [4]. Carboxymethyl cellulose/soy protein coating formulations containing 0.5 percent ascorbic acid applied to freshly cut apples were more effective in antibrowning activity than aqueous solutions of 0.5 percent ascorbic acid alone [47].
Enzyme treatments: Certain enzymes can be used to control browning either by substrate and/or product modification, and direct inactivation by use of proteases. Apple juice treated with the bacterial enzyme protocatechuate-3, 4-dioxygenase prevented browning by modifying its phenolic constituents which resulted in PPO enzyme deprived of substrates. The plant proteases including ficin, papain and bromelain are reported to be very effective inhibitors of browning [47]. Ficin was observed to be effective in preventing black spot formation in shrimp under refrigerated conditions.
Transgenic approach
Antisense gene introduction: One of the recent approaches for controlling browning is inhibition of gene expression in plants by introduction of antisense genes [47]. It involves insertion of a gene or a significant part of it, into the cell in a reverse orientation. Messenger RNA encoded by this antisense gene undergoes hybridization with mRNA encoded by the endogenous PPO gene, preventing production of the protein product. Gene silencing or the elimination of expected phenotypic characteristics, through antisense techniques has received much attention in recent years. Bachem et al. [62] reported that the expression of PPO in potatoes was decreased through the use of vectors carrying antisense PPO cDNAs. Approximately 70 percent of the transformed plants had lower PPO activity than controls, and on visual scoring, a significantly low level of discolouration. Insertion of PPO in the sense orientation resulted in very high PPO activity in the lines expressing the construct. This technology of genetically preventing enzymatic browning in a wide variety of food crops can be used as an effective alternate to chemical and physical treatments [47].
Considerable work on biochemal and molecular studies of PPO enzyme from various sources has resulted in current understanding of its structural and functional characteristics. The food industry has adopted methods to control the enzyme action in critical steps of processing in case of many food items. However, the problem is not solved in many ready to eat fruits and ready to cook vegetables where the browning takes place immediately after cutting. Transgenic plants are reported to have controlled browning due to reduced PPO activity. However, its wider acceptability remains to be achieved. Parallely there is a scope for using specific GRAS inhibitor(s) in food applications.