Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Editorial - (2012) Volume 1, Issue 6
High Mobility Group Box 1 (HMGB1) is highly conserved through evolution, and has 99% identity among all mammals. Out of its 215 amino acids, only two residues are substituted in the rodent and human versions. HMGB1 was originally described as a nuclear protein that binds DNA, and functions as a co-factor that is required for transcriptional regulation and gene expression.
<High Mobility Group Box 1 (HMGB1) is highly conserved through evolution, and has 99% identity among all mammals. Out of its 215 amino acids, only two residues are substituted in the rodent and human versions [1-3]. HMGB1 was originally described as a nuclear protein that binds DNA, and functions as a co-factor that is required for transcriptional regulation and gene expression [4]. Furthermore, HMGB1 is a potent proinflammatory cytokine and has been associated with a variety of inflammatory diseases, especially sepsis [5], and thus, it has been categorized as a danger signal. Numerous attempts have been made to treat sepsis by targeting well-known proinflammatory cytokines, including tumor necrosis factor (TNF). Although these approaches have led to the development of successful therapies for rheumatoid arthritis (RA) and Crohn’s disease, they have not proven effective regarding sepsis. A major obstacle in targeting TNF or other proinflammatory cytokines in the treatment of sepsis is that most of these cytokines are released early during the septic response, usually within hours after the exposure to the sepsis-inducing stimulus HMGB1- stimulation of neutrophils has been reported to induce an activated phenotype as determined by gene expression profiling, including increased production of TNF, IL-1 g and IL-8 [6]. Human umbilical vein endothelial cells (HUVEC) stimulated with full-length protein or the B-box of HMGB1 dose-dependently upregulated cell adhesion molecules such as ICAM-1, VCAM-1 and E-selectin and released IL-8 and G-CSF [5,7]. Human microvascular endothelial cells also responded with up-regulation of adhesion molecules and production of proinflammatory cytokines upon HMGB1-stimulation [7]. HMGB1 is thus able to induce production of multiple proinflammatory mediators by a variety of cells, and importantly it is possible that it might induce its own release from monocytes and macrophages. This implicates HMGB1 as a central player in proinflammatory reactions. Several proinflammatory activities of HMGB1 have been revealed from in vitro and in vivo. A picture has emerged of HMGB1, released into the extracellular milieu, having significantly proinflammatory functionality. HMGB1 is also a potent proinflammatory cytokine and associates with a variety of inflammatory diseases, especially sepsis (Figure 1).
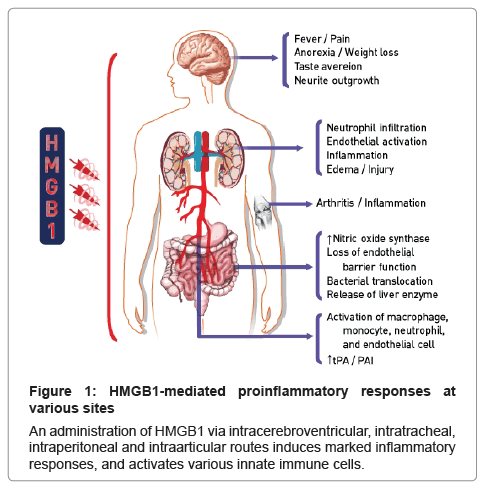
Figure 1: HMGB1-mediated proinflammatory responses at various sites
An administration of HMGB1 via intracerebroventricular, intratracheal, intraperitoneal and intraarticular routes induces marked inflammatory responses, and activates various innate immune cells.
With the discovery of HMGB1 as a potent mediator of inflammation and the presence of extranuclear HMGB1 in several inflammatory conditions, investigations of possible beneficial effects of HMGB1-targeted therapies were initiated. The appearance of HMGB1 much later than TNF and IL-1 in the inflammatory cascade suggested that HMGB1-targeted herbal therapies might be initiated even at late stages of acute inflammatory conditions. We have evaluated the immunomodulatory properties of chinese herbals. We recently showed that herbal single compound down-regulated the cell surface expression of the three receptors, TLR-2, TLR-4, and RAGE, which are known to bind HMGB1 to initiate pro-inflammatory responses in endothelial cells [8-14]. Similar to HMGB1, LPS also upregulated the expression of all three receptors on endothelial cells, and single compound from herbal medicines down regulated this effect [8-18], suggesting that herbal compounds can inhibit the amplification of pro-inflammatory pathways propagated by LPS and HMGB1 during infections. There are several potential mechanisms by which herbal compounds could interfere with HMGB1 functions. (1) Herbal compounds could inhibit the binding and interactions of HMGB1 to its receptors. This concept was supported by the finding that herbal medicine blocks chemokine binding to its receptors [8-14]. (2) Herbal compounds could block the downstream signaling pathway of receptors activation by HMGB1; this is also supported by the finding that recently we showed that herbal compounds inhibited LPS-stimulated NF-κB activation which is the down-stream transcriptional factor of HMGB1 [8,11,13-16,18-22]. (3) Herbal compounds could enter cells and interact with proinflammatory intracellular kinases or signaling pathways such as NF-κB, c-Jun N-terminal kinases (JNK1/2) or signal transducer and activator of transcription 3 (STAT3) and inflammatory intracellular molecules such as interleukin-6 (IL-6), IL-8, monocyte chemoattractant protein 1 (MCP-1), TNF-α, IL-1β or atrogin-1. For example, curcumin (from Curcuma longa), inhibited inflammatory signaling pathways such as NF-κB pathway, STAT and COX-2 signaling and it inhibited cell proliferation pathways such as phosphoinositide 3-kinase, protein kinase B and mammalian target of rapamycin [23]. Therefore, the potential molecular mechanisms for barrier protective effects against HMGB1 signaling might be mediated by inhibiting the interaction of HMGB1 to its receptors, inhibiting the inflammatory signaling pathway of HMGB1 or nuclear signaling pathways of HMGB1. In summary, beneficial compounds inhibit LPS-induced HMGB1 release and HMGB1-mediated secretion of TNF-α and NF-κB. More importantly, the inhibition leads to functional attenuation of barrier permeability, monocyte adhesion and migration through the HMGB1-primed endothelial monolayer by inhibitory effects of natural compounds on the expression of CAMs and HMGB1 receptors. Therefore, we conclude that natural compounds may contribute to the therapeutic potential of treating vascular inflammatory diseases.