Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2014) Volume 2, Issue 1
Prebiotics have been found to have prophylactic potentials against colon cancer. Thus, the present study was designed to delineate the prophylactic role of prebiotics in experimental colon carcinogenesis. It was found that inulin+DMH-treated mice had significantly increased (p<0.05) body mass till 5th week and thereafter, decreased while in lactulose+DMH-treated mice the body mass increased significantly (p<0.05) from 5th week till end of experiment compared with DMH-only treated mice. Further, it was observed that mice belonging to inulin+DMH and
lactulose+DMH-treated groups had significantly (p<0.05) higher lactobacilli counts compared with DMH-only-treated mice. It was also observed that inulin+DMH-treated mice had significantly (p<0.05) decreased colonic pH compared with lactulose +DMH treated mice while no significant difference was found in the colonic mass. Short chain fatty acid mainly butyric acid increased significantly (p<0.05) in inulin+DMH and lactulose+DMH-treated mice compared with DMH-only-treated mice. Thus, it can be concluded that probiotic inulin had better anticarcinogenic property than lactulose in reducing the DMH induced colonic alterations.
Keywords: Inulin; Colon; Cancer
Colon cancer is the third most common cancer after lungs and breast cancer worldwide and is one of the major causes of mortality [1,2]. Colon carcinogenesis is a multistep process in which preneoplastic lesions accumulate in mucosal cells due to various genetic/molecular changes and diet has long been thought to have a role in its etiology particularly unbalanced diet, obesity, physical inactivity and unhealthy practices, such as smoking and alcohol consumption [3-5]. World Cancer Research Fund and American Institute for Cancer Research 2007, has emphasized that regular physical activity and a healthy diet can help to reduce the risk of colorectal cancer. Therefore, attempts are being made to assess the effect of various natural bio-interventions such as phytochemicals, probiotics and prebiotics on the occurrence and management of various lifestyle diseases including colon cancer. Prebiotics, the dietary modifiers are defined as the “non-digestible food ingredients that beneficially affect the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon” [6]. Since prebiotics modify the composition and metabolic activity of the human useful microbiota, they form the basis for their anticancer properties and can be referred as probiotic enhancers or modifiers [7,8]. Moreover, these carbohydrates are not digested in the small intestine and reach the ileocaecal region in a relatively unmodified form where they can act as dietary bulking agents or make up a percentage of available substrate for resident colonic bacteria, consequently, contributing towards a decrease in pH and Short Chain Fatty Acids (SCFA) production [9-11]. In our earlier studies, we have observed that supplementation of probiotics (Lactobacillus acidophilus, Lactobacillus rhamnosus) and prebiotic (inulin) separately to Sprague Dawley rats was effective in modulating the procarcinogenic markers i.e. aberrant crypt foci, fecal enzymes (nitroreductase, β-glucuronidase, β-glucosidase) in DMH- induced colon cancer but the prophylactic effect of prebiotic in terms of biochemical alterations in experimental colon cancer warrants further investigation [12,13]. Thus, the study was designed to delineate the biochemical modulatory potentials of prebiotic in experimental colon carcinogenesis.
Chemicals
1,2-Dimethylhydrazine dihydrochloride (DMH) and phenolphthalein-β-D- glucuronide were obtained from Sigma Chemical Company, St. Louis, MO, USA. Prebiotic inulin was procured from Hi-Media, Mumbai, India.
Mice
Male LACA mice aged 5-6 weeks old (20-25 grams) were procured from the Central Animal House, Punjab University, Chandigarh and were housed under the standard conditions of light and dark cycle. These mice were fed with the standard pellet diet, and water ad libitum. Care, use and disposal of mice were done in accordance with the guidelines of Punjab University Animal Ethical Committee (IAEC), Chandigarh and approved by the Committee for the Purpose of Control and Supervision on Experiments on Mice (IAEC/156; 25/08/2011).
Induction of colon carcinogenesis
1,2-dimethylhydrazine dihydrochloride (DMH) was dissolved in 1 mM EDTA saline and pH was adjusted to 7.0 with 1 mM NaOH. Mice were injected intraperitoneally with DMH (20 mg/kg body weight) once in a week [12].
Prebiotic
Inulin (2 mg/0.1 ml distilled water) and lactulose (2.8 mg/0.1 ml distilled water) were fed to mice Intraoesophageally via canula [13].
48 mice were divided into six groups.
Group I (Control)
These mice received a single dose of EDTA saline intraperitoneally once in a week for six weeks.
Group II (DMH)
Mice were given a single dose of DMH (20 mg/kg body weight) intraperitoneally in a week and the treatment was continued till 6 weeks. Group III (Inulin): Mice were administered inulin (2 mg/0.1 ml distilled water) orally daily for 7 weeks. Group IV (Inulin + DMH): Mice received inulin (2 mg/0.1 ml) orally daily for a week. From second week onwards, mice were given a single dose of DMH intraperitoneally per week for 6 weeks. However, daily administration of inulin was continued till completion of the experiment. Group V (Lactulose): These mice were fed orally with lactulose (2.8 mg/0.1 ml D.W) daily for 7 weeks. Group VI (Lactulose+DMH): Mice received lactulose (2.8 mg/0.1 ml) orally daily for a week. From second week onwards, mice were given a single dose of DMH intraperitoneally per week for 6 weeks. However, daily administration of lactulose was continued till end of the experiment.
Follow up of mice
After respective treatment of mice, the body mass, lactobacilli count and short chain fatty acids in faeces were analysed. However, to assess the intestinal mass and pH alteration in the large intestine, mice were sacrificed by cervical dislocation after 7 weeks of treatment.
Determination of body mass
The body mass of mice were recorded once in a week by weighing on balance. (SD-300, S.D. Fine Chemicals Ltd, Chandigarh, India).
Enumeration of lactobacilli in faeces
To confirm that lactobacilli were able to survive the stress within the Gastrointestinal Tract (GIT), freshly voided faecal material (0.5 g/mouse) from each group was homogenised in normal saline and serially diluted. The diluted homogenates (0.1 ml) were spread plated on MRS agar incubated at 37°C for 48 hours and Colony Forming Units (CFU) on the plates were recorded. The lactobacilli count was performed once in a week for 7 weeks [14].
Determination of intestinal mass and pH of the colon
Mice were sacrificed after the completion of treatment after 7 weeks. On the day of sacrifice, these mice were fasted for 24 h, colon was removed, weighed on digital weighing balance [15] and intestinal pH was determined by using pH strips with 4.9-6.9 pH range.
Short chain fatty acids estimation
Short Chain Fatty Acids (SCFAs) were measured using method as described by Richardson et al. [16] with minor modifications of capillary Gas Chromatography (GC) method. In brief, 100 mg of fecal samples (stored at -20°C) were taken in Micro Centrifuge Tube (MCT) and homogenized by vigorous mixing on vortex and ultra-sonicated with 1 ml internal standard (100 mmol/l solution of 2-ethyl butyric acid in water). Sample in MCT was pelleted by centrifugation at 10,000 rpm for 1 min. To supernatant, 0.1 ml water, 0.5 ml hydrochloric acid and 2 ml ether were added and mixed by vortex, and centrifuged at 3,000 rpm for 10 min. The supernatant was dried using rotavapor and resultant nonvolatile substances were derivatized by adding 0.2 ml N-tert-butyldimethylsilyl-N-trifluoroacetamide (BSTFA). The samples were incubated at 37°C for 30 minutes and were injected into GC column (DB-5, GC-MS triple Quad, Agilent technologies, 7890A). Mass of the resultant molecules was also analyzed by Mass Spectrometry (MS-7000) and in silico analysis was done by software MassHunter and compared against the standards of formic acid, butyric acid, acetic acid, propionic acid and succinic acid. Values were presented as percent change compared to control samples.
Statistical analysis
Results were expressed as mean ± standard deviation (SD). The inter group variation was assessed by one way analysis of variance (ANOVA) followed by Post Hoc LSD Tests. Statistical significance of the results was calculated at p<0.05.
Body mass
The body mass of DMH-only treated mice decreased significantly (p<0.05) compared with control mice (Figure 1). A significant increase (p<0.05) in the body mass of inulin+DMH-treated mice was observed till 5th week, thereafter, decreased significantly (p<0.05) whereas lactulose+ DMH-treated mice had significantly (p<0.05) increased body mass from 5th week onwards till end of the experiment compared with DMH-only treated mice. More specifically, the body mass increased significantly (p<0.05) only in mice belonging to lactulose+DMH-treated compared with inulin+DMH-treated mice (Figure 1).
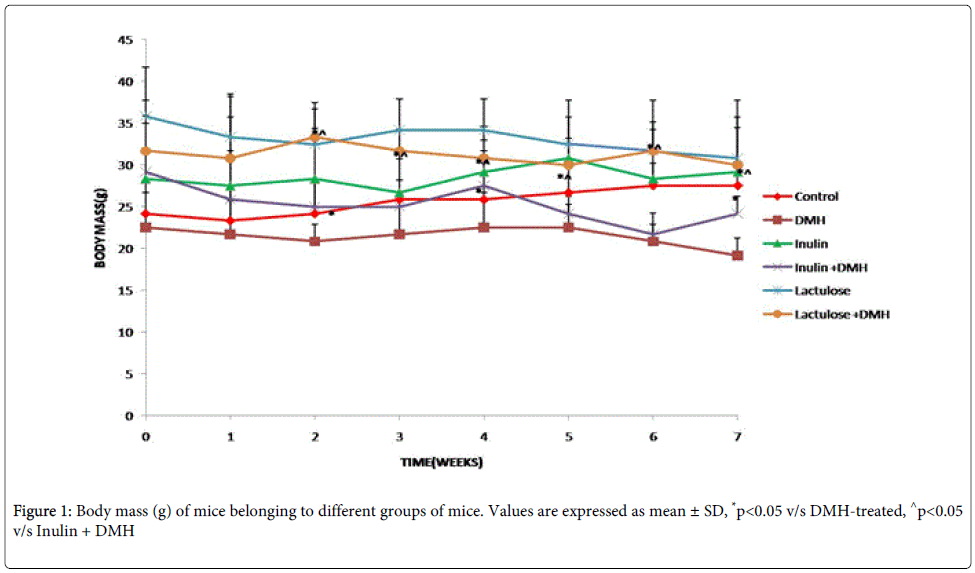
Figure 1: Body mass (g) of mice belonging to different groups of mice. Values are expressed as mean ± SD, *p<0.05 v/s DMH-treated, ^p<0.05 v/s Inulin + DMH
Lactobacilli count
It was interesting to note that mice belonging to inulin+DMH and lactulose+DMH-treated groups had significantly (p<0.05) higher lactobacilli compared with DMH-only-treated mice (Figure 2). Further, it was observed that inulin treated mice had significantly (p<0.05) increased lactobacilli count from 3rd week onwards till end of the experiment compared with lactulose+DMH- treated mice (Figure 2).
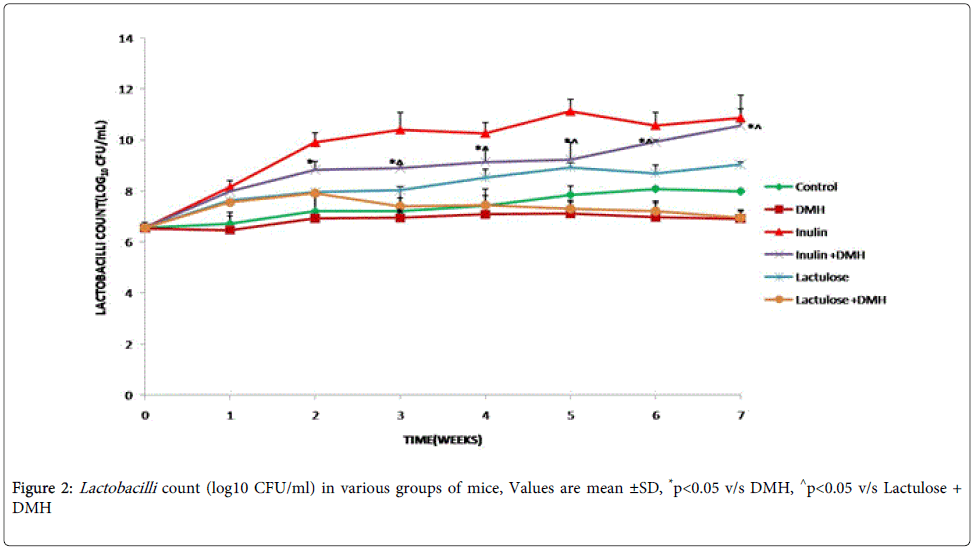
Figure 2: Lactobacilli count (log10 CFU/ml) in various groups of mice, Values are mean ±SD, *p<0.05 v/s DMH, ^p<0.05 v/s Lactulose + DMH
pH and mass of the colon
It was observed that supplementation of prebiotic, inulin to mice for 7 weeks led to significant decrease (p<0.05) in colonic pH compared with DMH-only treated mice. However, mice belonging to inulin+DMH-treated had significantly (p<0.05) decreased colonic pH compared with lactulose + DMH treated mice (Figure 3). Further, it was observed that mice administered only with inulin and lactulose had significantly (p<0.05) lower colonic mass while no significant difference was observed in the colonic mass of mice belonging to inulin + DMH and lactulose + DMH- treated groups compared with DMH-only treated mice (Figure 4).
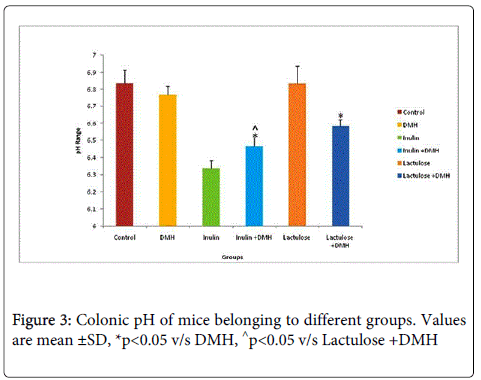
Figure 3: Colonic pH of mice belonging to different groups. Values are mean ±SD, *p<0.05 v/s DMH, ^p<0.05 v/s Lactulose +DMH
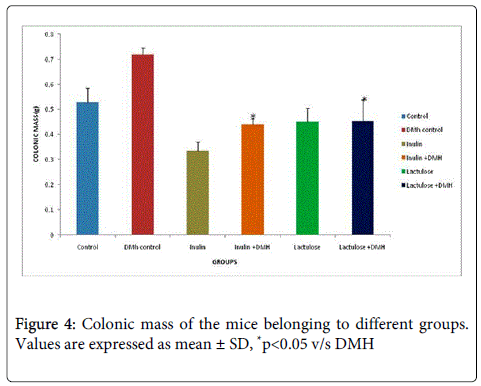
Figure 4: Colonic mass of the mice belonging to different groups. Values are expressed as mean ± SD, *p<0.05 v/s DMH
Short Chain Fatty Acids Estimation
Short chain fatty acids are the common metabolites produced during the end term digestion of food by gut microbe’s metabolic activity in the colon. It was observed that acetic acid content increased (p<0.05) in DMH-only-treated mice with no significant difference in inulin+DMH and lactulose+DMH-treated mice (Figure 5a). Interestingly, butyrate levels increased significantly (p<0.05) in both inulin+DMH and lactulose+DMH-treated mice compared with DMH-only-treated mice (Figure 5b). However, no significant difference was found in isovaleric acid and propionate levels in mice belonging to DMH-only-treated, inulin+DMH and lactulose+DMH-treated groups (Figures 5c and 5d)
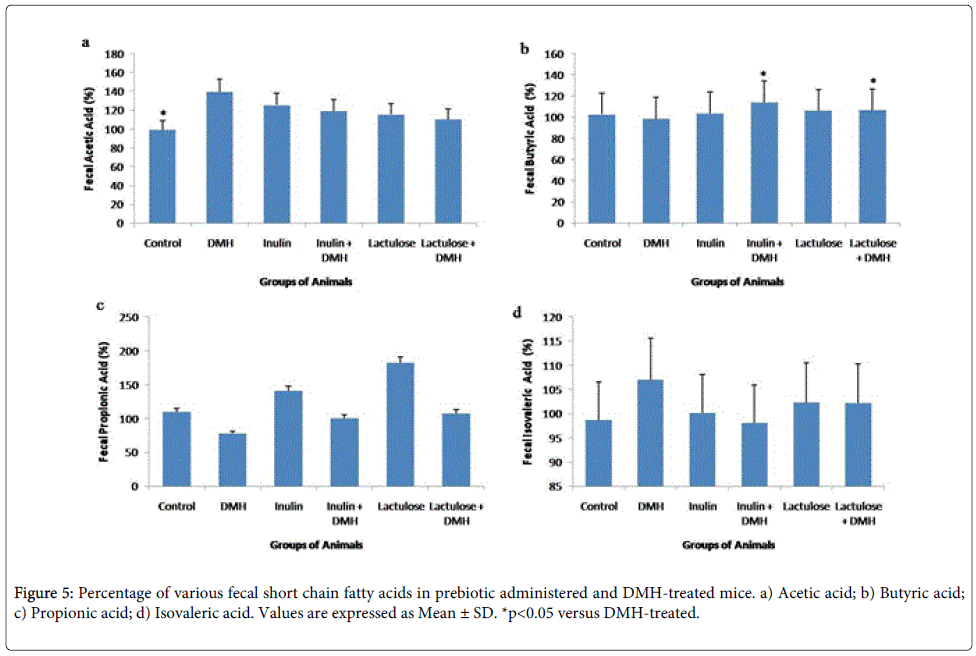
Figure 5: Percentage of various fecal short chain fatty acids in prebiotic administered and DMH-treated mice. a) Acetic acid; b) Butyric acid; c) Propionic acid; d) Isovaleric acid. Values are expressed as Mean ± SD. *p<0.05 versus DMH-treated.
Prebiotics provide an exciting new concept in human nutrition and digestive function for which many physiological and health beneficial effects have been claimed [17,18]. Our previous studies have also shown that prebiotics and probiotics have the potentials to reduce procarcinogenic markers (aberrant crypt foci, fecal enzymes) and retain the colonic cellular morphology in six weeks of DMH-treated mice [12,13]. Thus, the present study was designed with an aim to further study the effects of lactulose and inulin in six weeks DMH-treated mice with special reference to colonic mass, pH, intestinal microbiome and SCFA production.
The increased body mass in mice belonging to either inulin/lactulose groups may be due to improved gut microbiota as evident by increased lactobacilli counts. However, mice belonging to inulin+DMH treated group had decreased body mass in later phase of experiment that may be due to the initiation of colon carcinogenesis induced by DMH, altered biochemical, physiochemical activity and production of short chain fatty acids and corroborates with earlier studies [19]. These scientists have also observed decrease in body mass of Fisher 344 male rats when administered with different inulin fractions. However, the increased body mass in inulin+DMH treated mice compared with DMH-treated animals corroborates with earlier observations where it has been demonstrated that administration of prebiotics (Synergy1-Beneo) increased the body mass significantly in rats administered with azoxymethane [20,21]. The decreased body mass observed in prebiotic inulin group of mice compared with control mice may be due to short chain fatty acids (propionate) produced by colonic fermentation that may have hypolipidemic effect due to decreased lipogenesis, thereby reducing the concentration of plasma Very Low-Density Lipoproteins (VLDL) [22,23]. We also observed that supplementation of lactulose has no effect on body mass and is consistent with the earlier observations [24,25].
It was interesting to observe that supplementation of inulin and lactulose increased the counts of lactic acid bacteria in feces after 2 weeks inspite of DMH treatment but decreased the colonic mass and pH. This may be due to colonization of lactic acid bacteria in colon and their enhanced metabolic activity as both inulin and lactulose are dietary fibres that favors the growth of resident microbiota. These lactic acid bacteria ferment prebiotics resulting in production of short chain fatty acids that in turn decrease the luminal pH and is in accordance with earlier studies [26,27]. These researchers have also found that lactate production by lactic acid bacteria decrease the luminal pH. The observed increased colonic mass in DMH-treated mice may probably be due to the DMH which may have led to the formation of Aberrant Crypt Foci (ACF).
In our earlier study also we have found that prior administration of prebiotic inulin was better than lactulose in reducing ACF counts as it is degraded slowly and passes further along the colon before being completely degraded [13,28]. Further, it was interesting to note that administration of either prebiotic inulin or lactulose increased the levels of butyric acid in feces that may have inhibited the proliferation of colorectal tumor cells probably by decreasing the intestinal pH which may have in turn favored the growth of colonic microbiota and this observation is observed by earlier studies where butyrate has been shown to inhibit the proliferation of colorectal tumor cells [29,30]. Moreover, butyric acid, decreases the intestinal pH that is beneficial for the colon since high colonic pH has been shown to promote colon cancer [30]. Based on the observations of the present study the underlying proposed anticarcinogenic mechanism of inulin was due to its ability to modify the colonic conditions like colonic pH which affected the growth of colonic microbiome leading to enhanced lactobacilli that produces short chain fatty acid (butyrate) by fermentation which in turn inhibited the cell proliferation.
Taken together, the observation of the present study further strengthens our earlier observation of anticarcinogenic nature of prebiotic which provides a solid practical basis of prebiotic application that helps in reducing and even reversing the colon cancer. As inulin was found to have better anticarcinogenic property than lactulose due to its ability to reach distal colon, produce short chain fatty acid (butyrate) in the colon that acidify the environment resulting in decreased colonic pH and thus enhancing the growth of lactobacilli and reducing the levels of procarcinogenic markers i.e. aberrant crypt foci and procarcinogenic fecal enzymes. Thus, it can be said that diet rich in fibres modulates the gut microbiome subsequently affecting the metabolic profile and human health. However, these observations needs to be further correlated with clinical studies.