Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Review Article - (2013) Volume 1, Issue 3
Keywords: Colon cancer; Arachidonic acid; COX-2; Wnt signaling; Probiotic; PPARγ
All over the world, cancer is the most common cause of death and is rapidly overtaking Cardiovascular Diseases (CVD) as the prime cause of mortality in many countries [1]. Mortality from colon cancer is second only to that of lung cancer in men and breast cancer in women. Molecular studies indicate that colon carcinogenesis is a multistep (accumulated genetic alterations), multipath (multiple functional pathways) and multifocal process, frequently driven by genetic instability. Colon tumorigenesis involves activating mutations in protooncogenes as well as genetic inactivating mutation in tumor suppressor genes including Adenomatous Polyposis Coli (APC) gene, p53. These genetic events lead to changes in signal transduction pathways which regulate apoptosis, cell differentiation and proliferation. Several signaling pathways are related to colon tumorigenesis such as APC, β-catenin [2], Cyclooxygenase-2 (COX-2) [3], Nuclear Factor (NF-κB) [4] and Phosphatidyl inositol 3-kinase (PI3)/AkT [5]. Therefore, therapeutic agent that interacts with different targets is likely to enhance the efficiency of cancer treatment and chemoprevention.
Several studies on mucosal immune responses against pathogenic microorganism have created interest in the commensal microflora of the gut, which has emerged as a field of scientific challenge in recent time. There are several ways through which host getting advantage from gut microbiota such as metabolism of nutrients, production of health benefit compounds, development of intestinal epithelium, protection against pathogenic micro-organism by contending for colonization [6]. However, colon cancer has been linked with altered homeostasis in gastrointestinal tract (GI tract) [7]. Dysbiosis, a state in which guts microbiota (commensal bacteria) population are altered or imbalanced results dysfunctional immune responses. Altered homeostasis caused by dysbiosis may be due to chronic infections, use of antibiotics, immunosuppressant drugs etc. This altered physiological environment of the gut possibly leads to colon cancer development. Altered colonocytes were found able to escape from apoptosis and attained extensive proliferation (Figure 1).
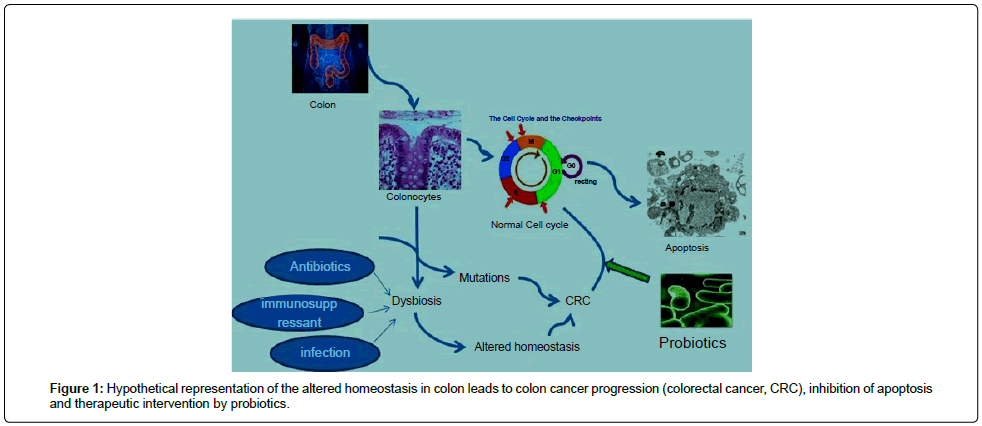
Figure 1: Hypothetical representation of the altered homeostasis in colon leads to colon cancer progression (colorectal cancer, CRC), inhibition of apoptosis and therapeutic intervention by probiotics.
Probiotics, an important component of the functional food are known to affect gastrointestinal tract immune response against colon cancer but the mechanism behind this is still poorly understood. Researchers suggested that there are several ways through which probiotics exhibit anti-cancer activity. It is documented that probiotics produces some health promoting metabolites during fermentations on dietary fibers in GI tract which have anti-colon cancer activities. These metabolites such as conjugated linoleic acid (CLA), butyrate and secreted proteins have ability to interact with several components of the cell signaling such as PPARγ, COX-2,Akt, MAPK and NF-κB which either are upregulated and/or downregulated during colon cancer development [2-5]. This interaction leads to the modulation of the signaling pathways and hence mitigation/reduction of colon cancer.
The immune response and their interaction in cell signaling have been thought to play important role in colon cancer progression, although little is known regarding the basic molecular mechanism of probiotic regulation/modulation of cancer signaling pathways. Extensive research on probiotics and immune responses indicates that probiotics and their cellular components could be able to induce cytokine production such as TNF-α, IL-12, IL-18 in human immune cells [8,9]. Lactobacillus casei shirota induces the NF-κB, STAT3, COX-2, PI3/Akt, and DNA binding activity [10]. These findings clearly suggest that probiotic strains can directly activate host immune cells as well as cancer cell signaling pathways involved in the generation of colon cancer. However, the exact mechanisms by which probiotic can modulate these cross talk remain obscure. In this review, we attempted to summarize how probiotics might play therapeutic roles in regulating or/and modulating colon cancer cell signaling.
Colon carcinogenesis is a multistep process in which several factors such as genetic mutation in tumor suppressor genes (p53, p15, p21, APC etc.), loss of DNA repair genes, and genomic instability are involved. These lead to the activation of several cell signaling pathways such as β -catenin in Wnt-signaling, COX-2 in arachidonic acid (AA) pathway, NF-κB and PI3/Akt pathways. These pathways cross talk each other through which cancer cells become everlasting [2-5,10].
Adenomatous Polyposis Coli (APC) is considered a tumor suppressor protein that acts as an antagonist of the Wnt signaling pathway. Defects in this gene cause Familial Adenomatous Polyposis (FAP), an autosomal dominant premalignant disease that usually progresses to malignancy and inhibits apoptosis of the colonocytes and causes the initiation of the colon carcinogenesis [11]. APC negatively regulate the concentration of β -catenin in cytoplasm by forming the degradation complex with GSK-3β , Axin, CK-1 (casein kinase-1), which lead to the phosphorolytic degradation of β -catenin. Loss of function of APC deregulate Wnt-pathways which further activate several down effectors (like E2F, Tcf, cycline D, c-myc etc.) and ultimately enhances the cell proliferation resulting accumulation of tissues with cancerous in nature (Figure 2).
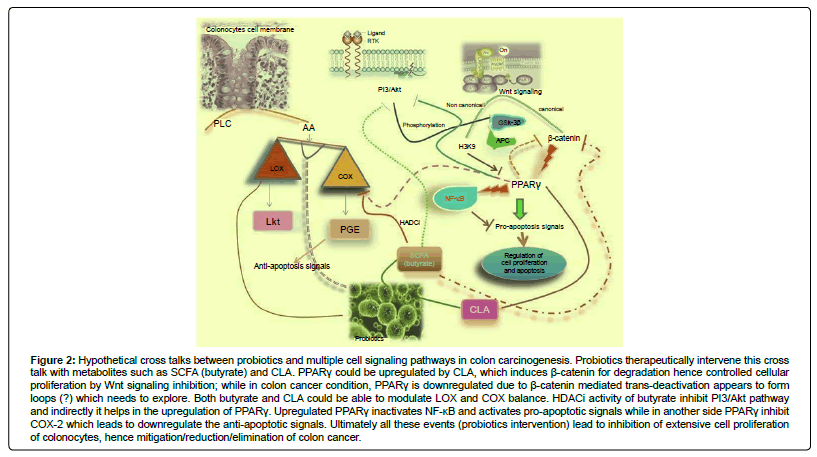
Figure 2: Hypothetical cross talks between probiotics and multiple cell signaling pathways in colon carcinogenesis. Probiotics therapeutically intervene this cross talk with metabolites such as SCFA (butyrate) and CLA. PPARγ could be upregulated by CLA, which induces β-catenin for degradation hence controlled cellular proliferation by Wnt signaling inhibition; while in colon cancer condition, PPARγ is downregulated due to β-catenin mediated trans-deactivation appears to form loops (?) which needs to explore. Both butyrate and CLA could be able to modulate LOX and COX balance. HDACi activity of butyrate inhibit PI3/Akt pathway and indirectly it helps in the upregulation of PPARγ. Upregulated PPARγ inactivates NF-κB and activates pro-apoptotic signals while in another side PPARγ inhibit COX-2 which leads to downregulate the anti-apoptotic signals. Ultimately all these events (probiotics intervention) lead to inhibition of extensive cell proliferation of colonocytes, hence mitigation/reduction/elimination of colon cancer.
In AA metabolism pathway, phospholipidic enzyme release AA from membrane which undergoes two major pathways, lipoxygenases (LOX) and cyclooxygenase (COX). LOX, in turn produces leukotrienes and COX, cyclooxygenase-2 which in turn gives various types of prostaglandins. Aberrant expression of COX- 2 was found in several malignancies [12]. Constitutive enzymatic expression of LOX and COX leads to cancer progression (Figure 2). Therefore balance between these two pathways is primary requirement for the maintenance of the homeostasis in GI tract. LOX pathway product 15-hydroxyeicosatetraenoic acid (15-HETE) which inhibits proliferation in PC3 prostate carcinoma cells, possibly through activation of peroxisome proliferative activated receptor-γ (PPARγ). Induction of PPARγ by LOX metabolites and COX-2 inhibitors and PPARγ effects on COX-2 activity, are evidence of crosstalk between effectors of AA pathway and PPARγ activity.
COX-2 is known to be a downstream target of the Wnt signaling pathway. The Wnt pathway is highly conserved evolutionarily and has been implicated in many biological processes. Signaling through the Wnt pathway increases expression of COX-2 mRNA, COX-2 protein, and PGE2 levels [13]. Few NSAID (non-steroidal anti-inflamatory drugs) such as Troglitazone and Rosiglitazone which act as ligands for PPARγ [7] and are able to cross talk with Wnt/β -catenin signaling in a number of cancers such as Hepatocellular, Breast, NSCLC, Neuroblastoma, Astrocytoma, Glioma, Prostate, Thyroid, Gastric, and Pancreatic [14]. These clearly show that Wnt pathways are interconnected with AA pathways and hence crosstalk.
Wang et al. [15] has demonstrated the involvement of PI3-kinase/ Akt pathways in cell cycle arrest and differentiation of colon cancer cell lines, HT29 and Caco-2. These researches further showed that Akt phosphorylates Glycogen synthase kinase-3β (GSK-3β ) which is involved in the inactivation of β -catenin. Thus GSK-3β might be involved in cross talk and anti-neoplastic activity with regulation of Wnt/β -catenin pathway [15]. Akt pathways deliver the cell survival signals by inhibiting apoptosis signals. NSAID inhibits the colon carcinogenesis by targeting the Wnt/β -catenin pathway and PI3/Akt pathways. As mentioned earlier that PPARγ ligands can cross talk with Wnt/β -catenin pathway which causes a decrease in β -catenin expression in adipocytes in In vitro and in normal intestinal mucosa in mice [15]. These findings indicate towards the convergence of several pathways and their cross talk. Therefore, the therapeutic agent which is able to arbitrate this crosstalk, could be a promising candidate for colon cancer modulation and mitigation.
Probiotics are efficient to exerts health beneficial effects by immunomodulation of GALT (gut-associated lymphoid tissue) and maintain immune homeostasis [6]. Among intestinal microbiota, probiotics are health promoting mucosa adherent species. According to WHO/FAO [16], probiotic are “live microorganism which when administered in adequate amount confers a health benefit on the host”. Probiotics is an important component of functional food, and their health benefit effects are unique and strain specific. Research evidences reveal the interactions between the enteric or gut microflora, host genotype and host immune system. Majority of bacterial gut microbiota possibly are probiotic in nature and hence enable to sustain the host homeostasis. Thus, this may be hypothesized that the interaction (negative) between colonocytes and colonized microbiota may be decisive for carcinogenesis at colon.
Cell free supernatants of Bifidobacterium lactis 420, Lactobacillus acidophilus, and L. salivarius were able to modulate the gene expression involved in tight junction and regulate the expression of COX-2 and NF-κB [17], thus suggested that probiotic produced bioactive metabolites. Probiotics hence could be able to induce or modify the gene expression in GI tract.
Several researches show that probiotics are able to produce natural ligands by their metabolic activity like CLA (conjugated linoleic acid) [18]. CLA is the natural ligands for PPARγ and has better therapeutic prospect than NSAID used in cancer. CLA is a mixture of positional and geometric isomers (9-cis, 11-trans and 10-trans, 12-cis) of linoleic acid which is preferably present in dairy products. CLA is a powerful anticarcinogen in rat mammary tumor model with an effective range of less than 1% in diet [6,8,19]. Specific probiotic strain has the ability to biohydrogenate linoleic acid to CLA and can thus function as a PPARγ ligand [18,19]. By keeping PPARγ in the center of this cross talk, probiotics and its metabolites are able to modulate or regulate several signaling pathways. However, the detail mechanism is yet to be established by further research.
Probiotics have been reported for its inhibitory effect on the expression of COX-2, NF-kB, nitric oxide and cytokine expression [20]. Over expression of COX-2 in colonic cells gives colonocytes anti-apoptotic potential and cell proliferation ability. The probiotic mixture, VSL#3 includes L. casei, L. plantarum, L. acidophilus, L. delbrueckii subsp. bulgaricus which could be able to inhibit colonocytes proliferation by inducing proapoptotic pathway by regulation of the concentration of COX-2 in colon [8]. Putaala et al. [17] demonstrated that the probiotics are involved in the inhibition of colon carcinogenesis by the down regulation of COX-2 [17]. In another study, Otte et al. [20] showed that probiotics were able to regulate COX-2 gene expression significantly in comparison to heat inactivated probiotics. This study revealed that probiotics-metabolites are more prominent in immunomodulation and gene expression than their cellular components. Involvement of probiotics in signaling pathways also was shown in a study where probiotics acted as a suppressor of cytokine signaling (SOCS) in Helicobacter pylori infection in gastric cancer [9]. In general, several commonly used non steroidal anti-inflammatory drugs such as COX-2 inhibitors are used to inhibit colon carcinogenesis. However, NSAID have some detrimental effects associated with its persistent use, especially on mucosal integrity and microbiota. Due to obvious health beneficially effects of probiotics over mucosal immunity as well as anti-carcinogenic effects, it may be considered as future drug for such treatment replacing NSAID.
Metabolites like short chain fatty acids (SCFA) such as butyrate and propionate produced by fermentation activity of probiotics could be able to regulate COX-2 expression in Caco-2 cell lines [21] and this capability is strain specific. Butyrate is a potent histone deacetylase inhibitor (HADCi). HDACi represent a class of epigenetic agents that regulate gene expression by modifying chromatic structure. HDACi have been shown to induce G1-phase cell cycle arrest with up-regulation of p21 (gene silencing). During tumorigenesis, histone hypoacetylation can result in silencing of genes involved in regulation of cell growth, differentiation, and apoptosis. It also can take their DNA methylation ability to methylate COX-2 promoters for the regulation of COX-2 in intestinal epithelial cells (IEC). Thus, the probiotic metabolite, butyrate envisages its therapeutic property in cancer therapy. It is therefore, important to evaluate such potential in each individual organism (strain) carefully both In vitro as well as in vivo studies. Some strains produces CLA which efficiently binds and activates the PPARγ, and this upregulation causes the downregulation of COX-2 concentration in colonocytes. It appears that there might be two pathways: one is butyrate or/and propionate dependent and another via CLA, through which probiotics could be able to regulate COX-2 concentration in IEC (Figure 2). These two capabilities (CLA and SCFA) may or may not be present in each individual probiotic strain. The probiotic strains which have both the properties could be considered as most promising therapeutic candidate against colon cancer.
Mutation in APC gene may result colon cancer. It was shown that oral administration of microencapsulated L. acidophilus preparation (109-11cfu/ml) daily reduces the multiplicity, tumor size and cancer progression in mice model. The study thus clearly showed that probiotic could be able to modulate the gene expression of APC in colonocytes [22]. Kumar et al. [23] demonstrated that bacterialepithelial interactions could modulate ubiqutin-mediated degradation of important signaling intermediates, such as β -catenin and NF-κB [23]. Probiotic supplementation was found to modulate Wnt signaling pathway and concluded that butyrate was involved in cell cycle arrest of aberrant colonocytes and apoptosis [24]. In addition, biohydrogenation ability of probiotics enables to produce CLA, resulting upregulation of PPARγ. Upregulated PPARγ leads regulation of Wnt signaling pathway in APC gene mutated (Figure 2) colon cancer model interacting with β -catenin and T cell transcription factor (Tcf) -4. This inhibited Wnt pathways further causes the regulation of cellular proliferation molecules such as cyclin D and c-myc of colonocytes which ultimately results the inhibition of colonocytes proliferation [25].
Takada et al. [26] showed that Wnt signaling activation via different pathways (canonical and noncanonical) led to different effectors such as suppression of transactivation of PPARγ in canonical Wnt-β -catenin pathway. In contrast signaling via non canonical Wnt pathways activates the histone methyltransferase that represses PPARγ transactivation through histone H3K9 methylation of target genes [26]. Probiotics which could be able to produce metabolites such as butyrate and CLA, considered as more promising agents in the inhibition or modulation of Wnt signaling pathway. Both butyrate and CLA showed synergistic effects over the inhibition of Wnt pathway: butyrate inhibit the histone methyltransferase activity while CLA induced the downregulated PPARγ expression (Figure 2). So it is necessary to carry out research in this direction.
Several critical events such as apoptosis, inflammatory responses, and angiogenesis during colon cancer progression are mediated by PI3/ Akt pathways [27]. Regulation of PI3/Akt pathways is considered as an important therapeutic target in cancer therapy. Experimental study, conducted by Dai et al. [27] in dextran sodium sulphate (DSS) induced colitis (mice model) found that VSL#3 exerts anti-inflammatory activity by inhibiting PI3K/Akt and NF-κB pathways (Figure 2) which maintain COX-2 concentrations and decreases cytokines IL-6, TNF-α and increases the concentration of IL-10 and correlated with antioxidant properties of VSL#3. In addition to this VSL #3 were found able to intrude the cross talk between NF-κB, COX-2 and PI3/Akt signaling pathways [27].
In vitro studies indicate that structural components (glycoprotein, lipotechoic acids) of probiotics such as Lactobacilli, Bifidobacteria, S. thermophilus, L. lactis and E. faecalis could be able to bind mutagenic compounds and reduces their bioavailibity to cause mutagencity in GI tract, hence preventative in nature. The mutagen binding with cellular component is strain dependent, varies with pH and was found similar in both live and dead bacteria [28,29]. The capacity to bind mutagens is not much clear in human but some animal experiments strongly supports this view [29]. Moreover, recent assays performed with the colon cancer epithelial HT29 cell line have shown that some probiotic bacteria (belonging to the species L. helveticus, Bifidobacterium spp., S. thermophilus and L. bulgaricus) or a compound they produce can interact directly with colonic epithelial cells, decreasing growth rate and induced differentiation [29].
Current researches suggest that probiotics are able to produce SCFA (butyrate as an HADCi) and CLA (ligand for PPARγ) through which they exhibit several modulatory activities in colon cancer cell signaling. Metabolically produced compounds could be able to modulate the gene expression in IEC and maintain homeostasis in GI tract. But if probiotics have any regulatory or modulatory role in the above discussed cross talk in colon cancer progression, is a thrust area of research. Thus exploration of the role of probiotics in the multiple cross talk in colon cancer will improve our knowledge and also will help to provide new site of action targeting multiple signal transduction, which could also open new door for other types of cancer where these pathways cross talk.