Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2015) Volume 5, Issue 1
Keywords: Onygena corvina, Keratinase, Alkaline protease, Feather degradation
Poultry farms produce a huge quantity of feather waste annually [1]. The large amount of feathers discarded in garbage causes a serious local disposal and accumulative problem leading to environmental pollution. β-keratin, a kind of fibrous protein, is the most abundant (at 90% dry matter) and important structural protein found in feathers [2,3]. Other keratinaceous wastes, such as hair, bristle, wool, hooves and horns, are made primarily of α-keratin. These keratinaceous compounds are highly resistant to the action of weak acids, alkalis and organic solvents or hydrolysis by common proteolytic enzymes such as trypsin, pepsin and papain due to a high degree of cross-linking by disulfide bonds, hydrogen bonds and hydrophobic interaction [4]. The high content of cysteine residues (3-15%) contributes to keratin stability and recalcitrance by forming disulfide bridges between different twists in a single peptide chain and between chains in keratin [5]. It is reported that the content of cysteine in bird feathers can be up to 15% [6], indicating that feathers are very difficult to degrade. Applying physical and chemical methods to convert feathers into feather meal resulted in the loss of nutritionally essential amino acids and formation of nonnutritive amino acids [7]. Use of biological processes may therefore be a better approach for efficient conversion of keratin to animal feed. Several species of bacteria (Bacillus sp.) [8-13], actinomycetes [14] and fungi (Trichophyton sp., Microsporum sp., Aspergillus sp., Penicillium sp., Fusarium sp., Acremonium sp., Trichoderma sp. and Chrysosporium sp.) [15] are known for their ability to convert feather waste due to secretion of keratinolytic proteases.
Keratinolytic proteases, also known as keratinases, are robust extracellular induction enzymes and specific for highly recalcitrant keratinaceous substrates [16]. Microbial keratinases mainly belong to serine or metallo proteases [17]. Keratinases have multitude potential industrial applications in food and feed [18,19], in leather and fertilizer industries [20-22], in production of biohydrogen [23], in degradation of prions [24], for silver recovery from X-ray film [25] and also as detergent additives [26]. In the industrial enzyme market, proteases with high feather degradation capability are available mainly from Bacillus strains. Therefore, industrial application and commercial exploitation of microbial keratinases are still in the very early stages. Nevertheless, fungal keratinases have been studied increasingly for their technological advantages, such as high enzyme yield and easy downstream process, and for their role in medical and veterinary epidemiology [27,28]. However, most of these keratinolytic enzymes are derived primarily from dermatophytic ascomycetous fungi such as Arthroderma sp., Microsporum sp. and Trichophyton sp., [29,30] or from broad saprotrophic non-specialized ascomycetous fungi such as Trichoderma sp., Chrysosporium sp. and Aspergillus sp. [1,31,32]. The human dermatophytes are not acceptable for enzyme production at an industrial scale for safety reasons and approval for use of their proteases may also be difficult to obtain. By contrast, no single enzyme or blend of enzymes with strong keratin decomposition potential has been found among the saprotrophic non-specialized fungi.
Onygena corvina, belonging to the Onygenaceae family of Ascomycota, lives saprophytically on horns, hooves, feathers and animal hair. Figure S1 (Supplementary materials) shows O. corvina which is highly specialized to grow on horn (α-keratin). Many Onygenales were reported as keratinophilic fungi living either as saprophytes on keratin substrates or as pathogens on birds, mammals and humans [33]. However, there are relatively few reports on characterization of the keratinolytic proteases from O. corvina. As a non-pathogenic fungus, this species should be tested further for possible use for conversion of feathers and for biotechnological applications.
The present study aimed to investigate the capability of O. corvina to degrade feathers. This fungus is shown to produce alkaline keratinolytic proteases in liquid culture with duck feathers as sole carbon and nitrogen source. O. corvina readily grows on and degrades duck feathers and expresses high protease and keratinase activities. The protease and keratinase characterizations were further performed. The findings of this study suggest that O. corvina could be used for further converting feather waste into bioaccessible protein as valuable ingredients for animal feed.
Microorganism and growth conditions
O. corvina (strain number: CBS 281.48) and Trichoderma asperellum (strain number: CBS 131938) were obtained from the Central Bureau voor Schimmel cultures, Fungal Biodiversity Centre, Royal Netherlands Academy of Arts and Sciences (CBS-KNAW) and kept on potato dextrose agar plate at 4°C. Subculturing was done once a month.
For protease production, a 4 mm2 square of O. corvina mycelium from a PDA plate was inoculated in a minimal liquid culture medium containing 15 g/l duck feathers, 2 g/l KH2PO4, 0.15 g/l MgSO4 .7H2O, 0.3 g/l CaCl2, 3.3 g/l Tween 80, pH 8 and incubated at 25 °C on a rotary shaker (200 rpm) for eight days.
Duck feathers were obtained from Valbyparken, Denmark, in August 2012. The feathers were washed three times with tap water and distilled water. Then the feathers were cut into about 1 cm pieces and air dried. Before being used as sole carbon and nitrogen source in the minimal liquid medium, the feathers were further dried in an oven at 50°C to constant weight.
Optimization of protease and keratinase production conditions
Different duck feather concentrations (g/l: 5, 15, 25, 35) combined with different initial pH (pH 4, 5, 6, 7, 8, 9, 10, 11) of the duck feather degradation culture medium described under 2.1, with protease activity measurement after incubation for 0, 2, 4, 6, 8 and 10 days, were used to determine the optimal culture conditions for protease production.
Culture broths were centrifuged at 10000 × g for 10 min to remove residual feathers, and supernatants were used for estimation of protease and keratinase activities, soluble protein and thiol formation.
Determination of weight loss
Weight loss was estimated by the determination of the duck feather dry weight loss. Initial feather weight was determined as dry feather weight after dehydration at 50°C. Final feather weight was measured as the dry weight of the residual feathers after dehydration at 50°C. The weight loss in each experiment was determined using the formula:
Weight loss (%) = (initial feather weight – final feather weight)/ initial feather weight ×100
Determination of enzyme activity
Protease activity assay with azocasein: Protease activity was assayed using azocasein as substrate [34]. The reaction mixture contained 20 μl diluted enzyme solution and 20 μl 1.5 % w/v azocasein (Sigma-Aldrich) dissolved in 50 mM sodium carbonate buffer (pH 9.0). The reactions were carried out at 50°C for 60 min with constant agitation at 300 rpm using a TS-100 Thermo-Shaker, SC-20 (Biosan Ltd). After incubation, the reactions were stopped by adding 100 μl 0.4 M trichloroacetic acid (TCA) and incubating at 4°C for 30 min. Then the mixture was centrifuged at 16000 × g for 1 min to remove the substrate. 100 μl supernatant was transferred to a microtiter plate containing 25 μl of 1.8 M NaOH. Absorbance was measured at 405 nm using a plate reader. As a control, 20 μl 1.5% w/v azocasein dissolved in the same buffer (to that of the sample) was added to 100 μl 0.4 M TCA before the addition of 20 μl enzyme solution. Then the control mixture was incubated at 50°C for 60 min in the same way as the sample.
One unit (U) of protease activity was defined as the amount of enzyme resulting in 0.01 absorbance increase between the sample and control at 405 nm under the conditions given.
Keratinase activity assay: Keratinase activity was measured as described [35] with a few modifications. Keratin azure (Sigma-Aldrich) was used as the substrate. It was ground into a fine powder with mortar and pestle using liquid nitrogen. Then 0.4 g keratin azure powder was suspended in 100 ml 50 mM sodium carbonate buffer (pH 9.0). The reaction mixture contained 50 μl keratin azure suspension and 50 μl enzyme solution. Assays were performed at 50 °C for 24 h with constant agitation at 1000 rpm in a TS-100 Thermo-Shaker, SC-20 (Biosan Ltd). After incubation, the reactions were stopped by adding 100 μl 0.4 M TCA followed by centrifuging at 16000 × g for 1 min to remove the substrate. The supernatant was spectrophotometrically measured for release of the azo dye at 595 nm. As a control, 50 μl 0.4% w/v keratin azure suspended in the same buffer (to that of the sample) was mixed with 100 μl 0.4 M TCA before the addition of 50 μl enzyme solution and incubation at 50°C for 24 h. One unit of keratinase activity was defined as the amount of enzyme that resulted in an increase of 0.01 in absorbance at 595 nm under the above described.
Protein determination
The protein content was determined by the Bradford method with the BCA Protein Assay Kit (23227, Thermo scientific) and bovine serum albumin (BSA) as a standard.
Thiol formation determination
Free thiol groups were analyzed as described [36] with a few modifications. 20 μl NH4OH, 100 μl of 0.5 g/l NaCN and 100 μl MilliQ water were added to 100 μl culture broth supernatant. The mixture was incubated for 20 min at 25°C, followed by the addition of 20 μl of 0.5 g/l sodium nitroprusside. Absorbance at 530 nm was measured within 2 min.
Amino acid determination
Concentrations of amino acids in duck feather degradation culture broth supernatant were determined by Hitachi automatic amino acid analyzer L-8900 at the State Grain Administration, Chengdu food feed quality progress and testing center, Chengdu, Sichuan, China.
Characterization of the enzymes produced by O. corvina
Influence of temperature and pH on enzyme activity: To investigate the optimal temperature, enzyme reactions were carried out at different temperatures (30, 40, 50, 60°C) as described in section 2.4.1. In order to study thermal stability, the enzyme supernatant was preincubated for 60 min at 30, 40, 50, 60°C in 50 mM sodium carbonate buffer (pH 9), then the residual activity was measured as described in section 2.4.1.
Determination of the optimal pH of the enzyme supernatant was carried out over a pH range of 4-11 at 50°C. To study pH stability, the enzyme supernatant were incubated in buffers of different pH (2 × McIlvaine Buffer (pH 4, 5, 6, 7 and 8), 50 mM sodium carbonate buffer (pH 9, 10 and 11)) for 60 min at 4°C. Residual protease activity was then determined as described in section 2.4.1.
Effects of metal ions, protease inhibitors, organic solvents, detergents and reducing agents on enzyme activity: The effects of various metal ions, inhibitors, organic solvents, detergents and reducing agents on protease and keratiase activity were investigated by assaying the enzyme activity as described above after pre-incubation with each chemical for 15 min at room temperature. The following chemicals were used: CaCl2, MgCl2 .7H2O, CuCl2, ZnCl2, FeCl2 and MnSO4 (1 mM), phenylmethylsulfonyl fluoride (PMSF) (1 and 5 mM), β-mercaptoethanol (1 %) and ethylene diamine tetraacetic acid (EDTA) (1 mM), sodium dodecyl sulfate (SDS) , dithiothreitol (DTT) (5 mM), ethanol, methanol, isopropyl alcohol, glycerol, triton x-100, tween-20, tween-80 (1 %).
SDS-PAGE electrophoresis and zymography: SDS-PAGE was performed with 12% polyacrylamide gels. PageRuler™ Prestained Protein Ladder (10-170 kDa, 26617, Thermo scientific) was included as standard. After electrophoresis, the gel was silver stained following Pierce Silver Stain Kit (24612, Thermo scientific) protocol.
To prepare a zymogram assay [34], protease sample was mixed with native electrophoresis sample buffer (3.1 ml 1 M Tris-HCl buffer (pH 6.8), 5 ml 50% glycerol, 0.5 ml 1.0% bromophenol blue and 1.9 ml MilliQ water) in a 4:1 ratio without heat denaturation. SDS-PAGE was carried out at 4°C at a constant voltage of 80 V in 12% polyacrylamide gel. The gel was first washed with 2.5% (v/v) triton x-100 in 50 mM Tris- HCl buffer (pH 9) for 30 min and then washed with 50 mM Tris-HCl buffer (pH 9) for 30 min. Casein (1% w/v) in 50 mM Tris-HCl buffer (pH 9) was then poured onto the gel. After 60 min incubation at 37 °C, the gel was stained with Page Blue Protein Staining Solution (24620, Thermo scientific) for 15 min. When protease bands appeared on a blue background, the gel was washed using MilliQ water.
Effect of duck feather concentration and initial pH on the degradation of duck feathers by O. corvina
Highest protease activity occurred when O. corvina was grown for eight days at 25ºC, 200 rpm in duck feather degradation medium containing feathers at 5 g/l and pH 6 (Figure 1A). However, the highest feather decomposition was measured as soluble protein, thiol formation and keratinase activity when duck feather concentration was 15 g/l.
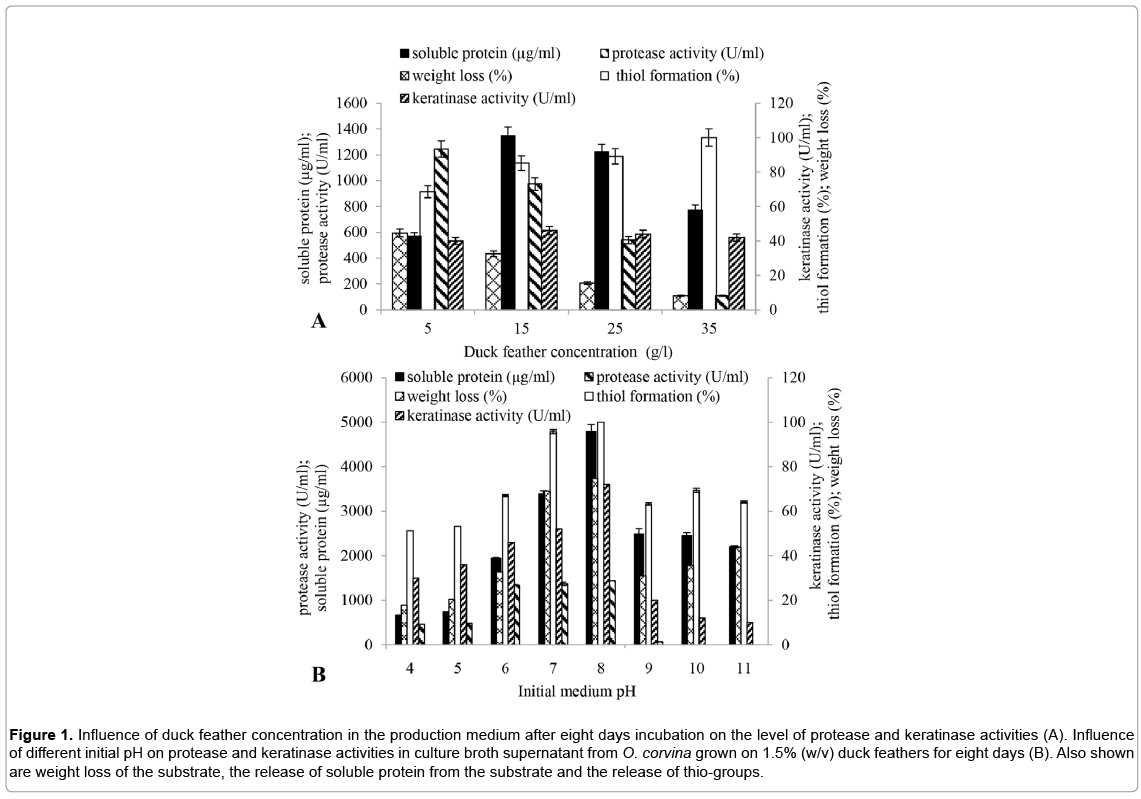
Figure 1: Influence of duck feather concentration in the production medium after eight days incubation on the level of protease and keratinase activities (A). Influence of different initial pH on protease and keratinase activities in culture broth supernatant from O. corvina grown on 1.5% (w/v) duck feathers for eight days (B). Also shown are weight loss of the substrate, the release of soluble protein from the substrate and the release of thio-groups.
Suitable initial pH for duck feather degradation by O. corvina was investigated at different initial pH values from 4 to 11 (Figure 1B). The highest protease and keratinase activities were obtained when the initial pH value was 8. Interestingly, the final pH increased to about 8.4 when the initial pH was between 4 and 8, whereas the final pH decreased to 7.0, 7.4 and 8.5, respectively, when the initial pH was 9, 10 and 11. Very low protease and keratinase activities were detected in the high pH media although the duck feathers were partially degraded. However, the alkaline environment contributed slightly to keratinolysis but it also seemed to make the feathers more accessible for degradation by the fungal proteins. This may be due to partial dissolution of the feathers by the alkaline environment. However, O. corvina does not grow well in the medium with initial pH 9 to 11. At pH 8 weak alkaline medium, partially dissolved feathers stimulated the keratinase resulting in complete degradation of the feathers. The results suggest that the tendency towards increase in pH of the acidic medium may be due to the keratinolysis of feathers. Kavitha and Hilda [37] have demonstrated that the change in pH at the end of fermentation is an indicator that keratinolysis has taken place.
O. corvina was cultivated in liquid medium (initial pH 8), containing 15 g/l duck feathers as sole carbon and nitrogen source. After incubation at 25 ºC, 200 rpm for eight days, the pH value of the culture broth increased to 8.5 and the protease activity and keratinase activity were 1435 and 72 U/ml, respectively (Figure 2). Soluble protein and relative thiol concentration increased from day 2 to 10. Furthermore, the amount of insoluble non-degraded feathers gradually decreased. Increased keratinase activity appeared to be related to an increase of soluble protein, which indicates that keratinase activity depends on O. corvina growth. During the cultivation, an accumulation of soluble proteins may be caused by both enzyme secretion and keratin solubilization. Keratinolysis is not only accomplished by keratinase but also by disulfide reduction mechanisms, such as through disulfide reductases, sulfide or thiosulfate chemical mechanisms, or by a cell-bound redox system [17,38]. So the increase in relative thiol concentration during cultivation may be attributed to the disruption of disulfide bridges. Gram-negative feather-degrading bacteria also increased the amount of thiol groups in growth medium [34].

Figure 2: Time course recordings of development of protease and keratinase activities in culture broth supernatant from O. corvina growing on 1.5% (w/v) duck feathers. Release of soluble protein from the substrate and of thio-groups is also given in the graph.
Different keratinolytic capability between O. corvina and T. asperellum
Other non-pathogenic fungi with high potential for biotechnological conversion of feathers into feather meal are Trichoderma species [1]. In the present work, T. asperellum was chosen as a positive control for duck feather degradation. T. asperellum was incubated for eight days at 25°C in duck feather medium, which resulted in a partial degradation of duck feathers. However, O. corvina was able to degrade most of the duck feathers. There was no degradation of the feathers in the negative control without fungus (Figure 3A). The weight loss of duck feathers incubated with O. corvina was 75% whereas T. asperellum only gave a weight loss of 23%. Moreover, O. corvina increased the soluble protein concentration, relative thiol concentration and the protease and keratinase activities far more than T. asperellum (Figure 3B). The O. corvina hydrolysate was also found to be higher in total amino acids concentration as well as in essential amino acids threonine, valine, isoleucine, leucine and phenylalanine concentrations when compared to T. asperellum (Table 1). Avinash et al. [39] reported that keratin is rich in amino acids like cysteine, leucine and serine. In our study, relatively high concentrations of threonine, glutamic acid, glycine, valine, isoleucine, tyrosine and phenylalanine were also detected in feather degradation culture broth inoculated with O. corvina. The results indicate that the non-pathogenic fungus O. corvina has high potential for feather decomposition and could be relevant for bioconversion of the feather keratin into high value-added products.
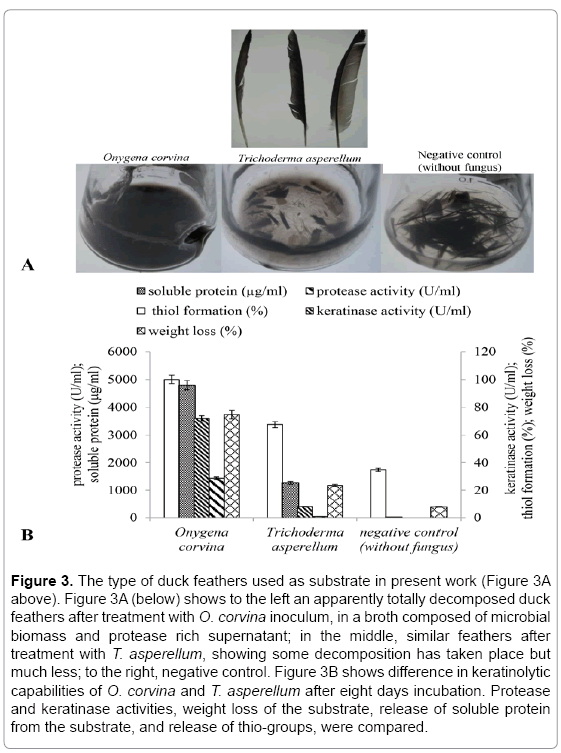
Figure 3: The type of duck feathers used as substrate in present work (Figure 3A above). Figure 3A (below) shows to the left an apparently totally decomposed duck feathers after treatment with O. corvina inoculum, in a broth composed of microbial biomass and protease rich supernatant; in the middle, similar feathers after treatment with T. asperellum, showing some decomposition has taken place but much less; to the right, negative control. Figure 3B shows difference in keratinolytic capabilities of O. corvina and T. asperellum after eight days incubation. Protease and keratinase activities, weight loss of the substrate, release of soluble protein from the substrate, and release of thio-groups, were compared.
| free amino acid concentration (%) | |||
| amino acid | O. corvina | T. asperellum | negative control (without fungus) |
| Asp | 0.003 | n.d. | n.d. |
| Thr | 0.044 | n.d. | n.d. |
| Ser | 0.054 | 0.001 | n.d. |
| Glu | 0.025 | n.d. | n.d. |
| Gly | 0.030 | n.d. | n.d. |
| Ala | 0.032 | n.d. | n.d. |
| Cys | 0.024 | 0.015 | 0.003 |
| Val | 0.054 | 0.001 | n.d. |
| Met | 0.001 | n.d. | n.d. |
| Ile | 0.038 | n.d. | n.d. |
| Leu | 0.055 | n.d. | n.d. |
| Tyr | 0.032 | n.d. | n.d. |
| Phe | 0.038 | n.d. | n.d. |
| Lys | 0.001 | 0.001 | n.d. |
| His | 0.002 | n.d. | n.d. |
| Arg | 0.019 | n.d. | 0.001 |
| Pro | 0.014 | n.d. | n.d. |
| Trp | n.d. | n.d. | n.d. |
| Total | 0.466 | 0.018 | 0.004 |
Table 1: Free amino acid concentration after feather degradation by the enzyme composition from O. corvina and T. asperellum n.d. not detected.
Characterization of the O. corvina feather degrading enzymes
Influence of temperature and pH on enzyme activity: Figure 4 shows that the protease from O. corvina was active at a broad range of pH (pH 6 to 11) and temperature (40-60ºC). Such wide pH and temperature range might be useful for industrial applications. Maximal protease activity was obtained at pH 9 and 50ºC, respectively. The protease was stable at pH 5-11 at 4ºC, and more than 71% residual activity was retained throughout the pH spectrum. The enzyme was stable for one hour at a temperature of 30 ºC, while the residual activity at 40ºC was 48%.
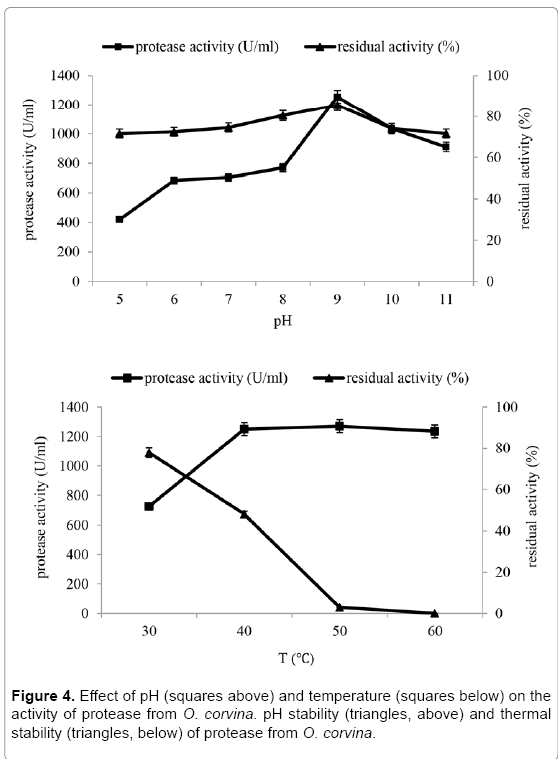
Figure 4: Effect of pH (squares above) and temperature (squares below) on the activity of protease from O. corvina. pH stability (triangles, above) and thermal stability (triangles, below) of protease from O. corvina.
Effects of metal ions, protease inhibitors, organic solvents, detergents and reducing agents: The keratinase from O. corvina was partially inhibited by Mg2+, Cu2+, Zn2+ and Mn2+ and stimulated by Ca2+ and Fe2+ (Table 2). Generally, Ca2+, Mg2+ and Mn2+ metal ions stimulate keratinase activity, and Cu2+, Zn2+ metal ions inhibit keratinase activity. But for O. corvina, Mg2+ and Mn2+ had negative effects on keratinase activity. Fungal keratinases mostly belong to the class of serine proteases, which are inhibited by PMSF and EDTA [17]. In the present work, keratinase activity was partially inhibited by 1 mM EDTA and 1 mM PMSF, and completely inhibited by 5 mM PMSF. This indicates that most keratinases from O. corvina may belong to serine proteases, but this result needs further investigation. Different organic solvents such as ethanol, methanol, isopropyl alcohol and detergents such as tween-20 and tween-80 inhibited keratinase activity to some degree. Similar results were found for keratinase from Bacillus subtillis [35]. But the enzyme from O. corvina was stable in the presence of glycerol; while enzyme from Bacillus subtilis was not. The keratinase activity of O. corvina was decreased by SDS and triton x-100 detergents, as was the enzyme from Streptomyces pactum [40]. Reducing agents like DTT and β-mercaptoethanol generally enhance keratinase activity because the addition of reducing agents can break disulfide bonds to help sulfitolysis [35]. But keratinase activity of O. corvina was partially inhibited by DTT and β-mercaptoethanol after pre-incubation for 15 min.
| concentration | protease residual activity (%) | keratinase residual activity (%) | |
| Ca2+ | 1 mM | 96 | 136 |
| Mg2+ | 1 mM | 100 | 67 |
| Cu2+ | 1 mM | 107 | 67 |
| Zn2+ | 1 mM | 100 | 44 |
| Fe2+ | 1 mM | 104 | 142 |
| Mn2+ | 1 mM | 108 | 92 |
| EDTA | 1 mM | 37 | 75 |
| PMSF | 1 mM | 41 | 33 |
| PMSF | 5 mM | 13 | 0 |
| ethanol | 1% | 96 | 58 |
| methanol | 1% | 99 | 44 |
| isopropyl alcohol | 1% | 98 | 56 |
| glycerol | 1% | 110 | 94 |
| triton x-100 | 1% | 74 | 39 |
| tween-20 | 1% | 86 | 33 |
| tween-80 | 1% | 104 | 67 |
| SDS | 1 mM | 95 | 47 |
| DTT | 1 mM | 99 | 61 |
| β-mercaptoethanol | 1% | 24 | 31 |
Table 2: Effect of several compounds on the crude protease and keratinase activities of O. corvina.
SDS-PAGE electrophoresis and zymography: The zymogram of the crude enzyme preparation revealed two clear bands (Figure 5), which suggest that the molecular weight of the secreted proteases was mainly estimated to 35 and 20 kDa, respectively, when using casein as substrate. It was reported that the molecular weight of keratinases ranges from 18-200 kDa [17], with most being around 20-60 kDa. Some of the secreted proteases of O. corvina, when grown on duck feather medium, may have very similar molecular weights and cannot be separated on the SDS-PAGE gel; this occurs especially with the proteases with molecular weights around 35 kDa, which merge into a strong band in this study (Figure 5).
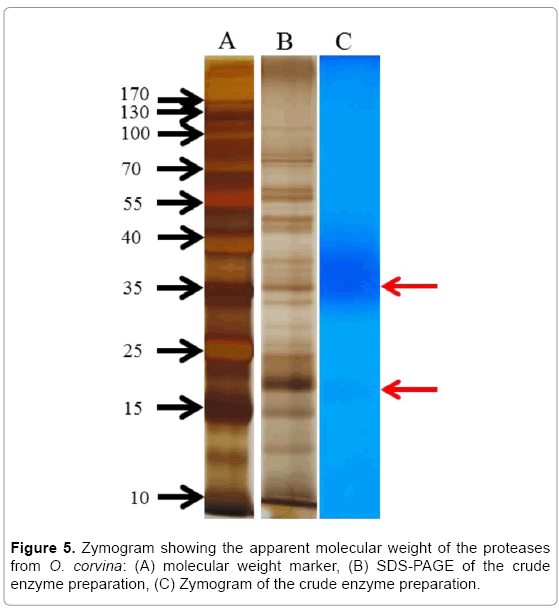
Figure 5: Zymogram showing the apparent molecular weight of the proteases from O. corvina: (A) molecular weight marker, (B) SDS-PAGE of the crude enzyme preparation, (C) Zymogram of the crude enzyme preparation.
In the present research, O. corvina, a non-pathogenic ascomycetous fungus, secreted highly active keratinolytic proteases when grown on duck feather medium. Duck feathers were completely degraded due to the high protease and keratinase activities (1435 and 72 U/ml, respectively). The proteases of O. corvina were active over a broad range of pH values (pH 6-11) and temperature (40-60ºC). The keratinase was sensitive to serine protease inhibitors and organic solvents and inhibited by most metal ions, but was stable in presence of glycerol and stimulated by Ca2+ and Fe2+. Compared to the potential feather degrading fungus T. asperellum, the enzymes of O. corvina could be of relevance for biotechnological processes for converting feathers into important bioaccessible protein rich animal feed.
This study was supported by the Sino-Danish Center (SDC) and Keratin2 Protein.