Immunological Disorders and Immunotherapy
Open Access
ISSN: 2593-8509
ISSN: 2593-8509
Research Article - (2017) Volume 2, Issue 3
Introduction: Our study focused on the value of TCD8 cytotoxicity in susceptibility to severe malaria in endemic areas. The global purpose of the work hereby was to evaluate adaptive cellular immunity during Plasmodium falciparum malaria through TCD8+ cytotoxic lymphocytes.
Patients and methods: It was a prospective study, with analytical purpose that took place over a period of 8 months in the Pediatric Department of Hôpital Général d’Abobo and in the Immunology and Hematology Laboratory of CHU de Cocody. The study focused on 50 children (under 15 years of age) selected on the basis of WHO definition criteria for malaria infections (40 children with simple malaria and 10 severe malaria) a fact sheet and 10 witness persons. The samples carried were sent and processed in the said-laboratory.
Results: Among these 50 children, those under 5 and over 5 years accounted for 52% and 48% of the size respectively. Most of them were boys with a sex ration of 1, 77. In children under 5 years, the average rate of TCD8 was higher in simple malaria (6098.16 cells/ml) than in severe malaria (3915 cells/ml) with a statically significant difference. On the other hand, in children over 5-year-olds, the difference noticed was not significant despite relatively higher TCD8 rates. However, regardless of the age of the child, the rate of TCD8 cells was higher in malaria than in witness.
Conclusion: The study hereby shows a gradual stimulation of the specific immune system by Plasmodium falciparum. The proliferation of TCD8+ lymphocytes in the simple form could be due to the immune activity which protects against the severe form where a clonal contraction of TCD8+ could be observed. A study with a larger sample seems necessary to draw a conclusion to the comparison of these cells.
Keywords: Malaria; TCD8 lymphocytes; Adaptive immunity; Ivory Coast
Malaria, a disease caused by protozoan parasites of the genus Plasmodium, is a leading cause of morbidity and mortality in developing world, that is 429,000 of annual deaths due to malaria. The large majority is caused by Plasmodium falciparum infections [1]. In Ivory Coast, malaria is hyperendemic and is transmitted throughout the year with an increase rate during the rainy seasons. Its morbidity is 42.28% for a mortality of 15.29% in children under 5 years [2]. In the issue of protection against severe forms of malaria in endemic areas, pre-munition or semi-protection has been mentioned as a protection factor acquired in adults after multiple infestations for several years [3]. Multiclonal infections have been suggested to provide protection against malaria by preventing superinfection or by promoting tolerance to infection [4-6]. However, the precise immunological mechanisms underlying this protection association are unknown. Despite that relative protection, 2 to 3% of adults living in endemic areas immunologically competent are victims of severe malaria (unpublished data Ivory coast). That allow to raise the question of susceptibility and resistance toward severe malaria. According to Adam E and al in animals, the susceptibility in the young rat to Plasmodium berghei is related to a low cytotoxic lymphocyte rate [7]. According to Dassé et al, susceptibility in the young rat is related to the dendritic cell OX62, CD4- responsible for the humoral profile that appears to be ineffective in protecting the young rat against Plasmodium berghei while the dendritic cell OX62, CD4 + is more linked to resistance in adult animals [8] Those data should be confirmed in humans. The purpose of the work hereby is to evaluate the rate of TCD8 profile in children in the different clinical forms of malaria.
Type and place of study
It is a prospective study with analytical purpose, study conducted between May and December 2014 after obtaining authorization from the Ethics Committee. The purpose of the study was to explain to parents and patients the use to participate in that study. Those who agreed to participate in the study were given a fact sheet with information to be filled in. Ethical permission for this study was granted by the Ethical Committee of the Hospital Pedia´trico Abobo General in Abidjan, appointed by the Ivory Coast Ministry of Health. Written informed consent was obtained in all cases from study participants or parents of minors. Regular visits were made to the site, patients were recruited from the Pediatrico Department of hôpital Général d’Abobo and selected on the basis of WHO definition criteria for malaria infections in 2000.
Study population
The study focused on children aged from 0-15 sent to Paediatrics in hôpital Général of Abobo during 2014 in two age groups (less than 5 years and over 5 years). On the overall patients registered, 60 were selected for the study. There were 40 children who were referred to for uncomplicated malaria, 10 were hospitalized for severe malaria, 10 were uninfected (patients in malaria endemic areas without clinical signs), and other pathological infections were eliminated on the basis of NFS and clinical signs.
Blood collection
Blood was collected by puncture of a peripheral vein after cleaning the surface with alcohol buffer in children aged from 0 to 15 years for the dosage of CRP, glycemia. Another EDTA tube collection helped to carry out NFS to look for platelet, hemoglobin, and leukocyte levels to eliminate a bacterial infection. Finally, the last collection on EDTA tube allowed to count TCD8 cells. Blood smear and thick drop were carried out for species confirmation and parasite density.
Flow cytometry analysis
We have used isolation and counting technique, then immunostaining technique with BD reagents (Becton Diskinson) to obtain the different proportion of TCD8 cell rates. 3 ml of whole blood diluted in half in PBS buffer after reconstruction was based on a tube containing 5 ml of ficoll. After centrifugation at 4°C and 3000 tours per minute for 5 minutes, 4 layers were formed, the red cell layer, the layer containing ficoll solution, the mononuclear cell layer and the serum layer. Mononuclear cell layer was re-collected, washed 3 times, then the pellet was taken up in 1 ml of PBS and finally counted on the Mallasez cell.
Then, we tagged those cells with specific antibodies.
• Anti CD3 FITC for T lymphocytes (including TCD8+).
• Anti CD8PE for TCD8 lot 4059802 reference ST / A1406109.
The reading was then done with flow cytometry model: Facs calibur® serie E34297300578 version 002 manufacturer BD Company and the analysis of the data acquired was carried out by Cell Quest Pro® software.
Statistical analysis
Data was entered using Microsoft Excel software Statistical analysis of those data was carried out using the Epi Info 2008 software (Version 3.5.1). For quantitative variables, our results were converted in averages. The test of Student was needed to compare the averages. We converted our results in numbers and percentages for qualitative variables. The comparison of percentages was carried out with Chi-2 Pearson test. The result was considered significant at the 5% level.
Epidemiological characteristics of the population
Repartition of patients based on age: Patients less than 60 months (5 years) followed by 60 to 120 months (5 to 10 years) have a higher frequency (Figure 1).
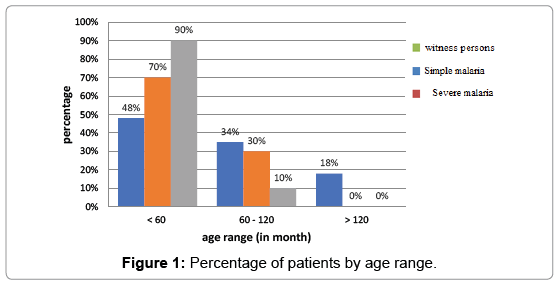
Figure 1: Percentage of patients by age range.
Repartition of study population based on sex: Within the study population, the sex rate is 1.77 in favor of men (Figure 2).
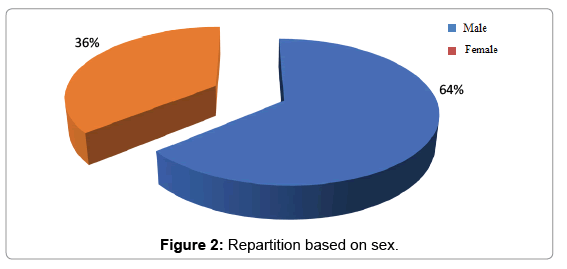
Figure 2: Repartition based on sex.
Biological characteristics
Comparison of TCD8 cell rate in the healthy person in endemic (witness person) area based on age: The average level rate of TCD8 + cells in witness persons over 5 years is meaningless compare to witness persons less than 5 years of age as shown in Figure 3 below.
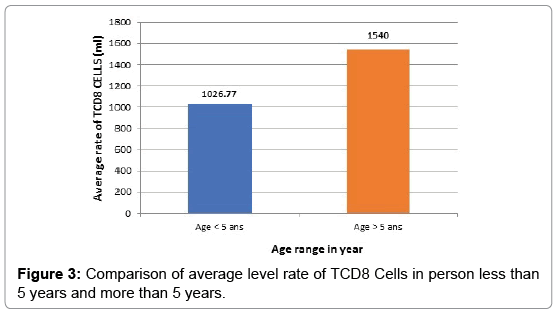
Figure 3: Comparison of average level rate of TCD8 Cells in person less than 5 years and more than 5 years.
Comparison of TCD8 cell rate based on clinical status and age:
1. In children under 5 years of age (Figure 4)
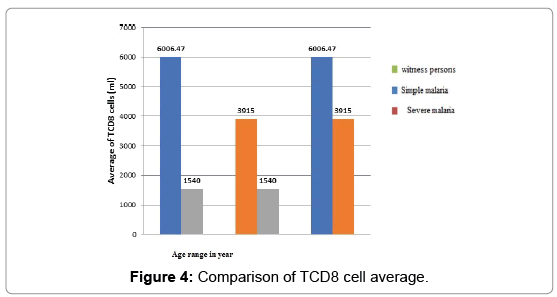
Figure 4: Comparison of TCD8 cell average.
2. There is a statistical difference between the levels of TCD8 cells depending on clinical status.
3. That rate is higher in simple malaria than in severe malaria compared to witness person.
For children over 5 years of age (Figure 5). In persons older than 5 years, the average number of TCD8 does not differ significantly in the two clinical forms of malaria (simple and severe malaria).
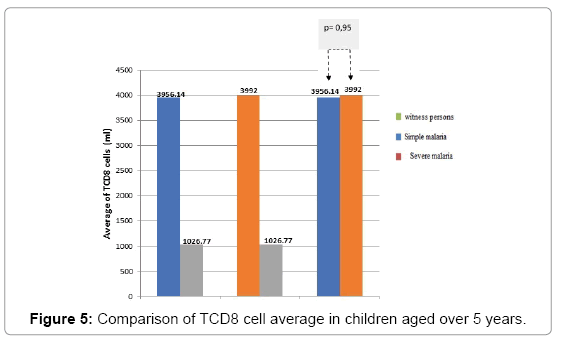
Figure 5: Comparison of TCD8 cell average in children aged over 5 years.
Epidemiological characteristics
Study based on age: 70% of severe forms of infection were more common in children under five years of age according to clinical status (Figure 1). That situation could be explained by the immaturity of the immune system at this age as evidenced by the studies on immunogenesis [9] i.e. the role of antibodies (humoral immunity).
As for the simple form, 18% of children over 5 years are victims. That case could be explained by the progressive introduction of a premunition well shown on the diagram (Figure 1). That part showed a decrease in the proportion of severe forms as the age of children increased.
The simple form persisted for up to 15 years, thus demonstrating the progressive introduction of clinical tolerance, materialized by the absence of the severe form 15 years.
Study based on sex: The repartition of our patients based on sex with a sex ratio of 1.77 in favor of men was superimposed on that of the population received in hôpital général of Abobo at the same period (report of activity of the hospital non-published). That shows the representativeness of our sample (Figure 2).
Analysis based on immunological characteristics
The profile of TCD8 cells based on age: If the acquisition of a clinical tolerance takes place during the years of life in endemic area as well shown here, it is at the price of a gradual introduction of semiprotective Antibody [9] and cellular immunity. In the study hereby, we first compared the average level rate of TCD8 cell based on age in healthy children. In witness persons, the rate of TCD8 is higher in persons over 5 years (1540 cells/ml) than in persons less than 5 years (1026.77 cells/ml). In person less than 5 years, the rate of TCD8 is higher in simple malaria (6006.47 cells/ml) than in severe malaria (3915 cells/ml). TCD8 cells appear to have a much higher in children over 5 years with severe malaria (3992 cells/ml) than in simple malaria (3956.14 cells/ml), but the statistical difference is not significant, in children under 5 years old it’s the contrary. Cerebral malaria (CM) is a severe and often fatal, complicated form of Plasmodium falciparum malaria affecting mostly children in endemic regions [10,11] However, one factor remaining implicit are CD8 T cells [12,13] although the exact mechanisms that drive their pathogenesis in CM remain unclear. In particular, type I IFN was shown to be involved in the priming of CD8 T cells [14,15] and in the generation of robust response to infection [16,17]. In contrast, the role of CD8T cells in CM is unequivocal [18,19]. Numerous studies have demonstrated that CD8T cell depletion [17,20] or ablation of effector functions [21,22] completely abrogates the disease. Furthermore, parasite-specific CD8 T cells can mediate CM in the absence of bystander T cells [15]. Despite the critical role played by CD8 T cells during CM, little is known about the dynamics, kinetics, anatomical localization, and function of these cells in vivo. A strong consensus exists in the literature among several studies showing that CD8T cells play an essential role in CM pathogenesis [15,17,20-27] and our data support this conclusion. That difference in TCD8 cell rate could also be explained by the high prevalence of viral infections of the ORL sphere which remain frequent after 5 years. Those infections induce chronic stimulation of cellular response [28]. In that study, we did not evaluate the ORL viral infection in our patients and witness persons. However, in the case of malaria, there has been a particular evolution of the levels of TCD8 cells, cytotoxic cells according to the clinical status and the age of the child. Thus, regardless of the age of the child, a higher rate of TCD8 cells was observed in malaria than in witness persons. Then, depending on the clinical status, the rate of these cytotoxic cells is higher in simple malaria than in severe malaria. These data show a stimulation of the specific immune system by Plasmodium falciparum.
The profile of TCD8 cells based on clinical status: In children over 5 years of age particularly, if the TCD8 + cell rate remains the same in simple and severe malaria, the small sample size of witness could not allow us to compare the rate of those cells between witness persons and case of simple malaria. A study with a larger sample seems necessary to draw a conclusion from the comparison of those cells. Nevertheless, the proliferation of TCD8 + lymphocytes in the simple form could be due to the important immune activity which protects against the severe form where a clonal contraction of the TCD+ could be observed. In addition, those data were confronted by the work of HILL et al. who have shown the relative protection of severe malaria in children with a major histocompatibility complex I type B 53 [24] (CMHI) in West Africa. It appears that specific form of MHC may show to TCD8 cells a particular peptide of the plasmodial protein LSA- 1 (« liver stage antigen-1 »). TCD8 cells can inhibit the development of sporozoites and delete serious complications [25]. Researchers on the other hand believe that TCD8 cells could eliminate the parasite without harming the hepatocytes by secreting cytokines such as INFgamma [26]. More recently depletion studies and adoptive transfer experiments have demonstrated that CD8 T cells have a leading role in protection against sporozoite challenge [27,28]. The critical role of CD8 T cells in protection against liver –stage malaria has been confirmed upon immunization with different formulations, such as recombinant vaccines and genetically attenuated parasite [29,30]. Despite the strong evidence for the role of CD8 T cells in sterile protection against malaria, critical qualitative and quantitative characteristics of the protection response and effector mechanisms engaged by CD8 T cells remain incompletely understood. The modest progress in this field is strongly influenced by the extreme genetic plasticity of the parasite, its complex life cycle and the paucity of defined antigenic targets for CD8 T cells. Filling these knowledge gaps is of an utmost importance as this information would facilitate the development of successful preerytrocytic vaccine candidates. Various effector molecules, such as IFN-y, TNF-ɚ, perforin FasL, and TRAIL, are utilized by memory CD8 T cells in protection against different infections [31,32]. Upon parasite clearance in humans CD8T cell responses are clearly directly elicited by liver stage infection [26,33-37] Activation by blood – stage infections by pass the liver stage. Prior studies suggest that murine CD8T cells recognize blood- stage rode malaria parasite [38,39].
We evaluated the rate of status of TCD8 cells in child malaria based on age range less than 5 years, over 5 years and clinical status. Our results revealed that once malaria is evolved in severe form, the effector defense by specific cellular immunity seems to take over through the TCD8 lymphocytes that we noted higher in the simple form (6006.47 cells/ml) in children under 5 years of age. Those data must be confirmed by a study that will take into account a wider sampling and will extend over a long period. Those studies should integrate the analysis of the immune response as a whole, taking into account the antibodies (IgM, IgG, IgE), cytokines during the infection of the child by Plasmodium falciaprum. Those collected data could provide insight into the understanding of immune response and antimalarial pathways to theurapeutical perspectives vaccination by inducing and stimulating that immune response.
We are grateful to the patients and their relatives for the active participation and continued collaboration in this work. The authors thank all the present and former members of Harty laboratory and all of our collaborators for their contributions to the work described herein.
The authors do not have any conflict of interest.