Journal of Agricultural Science and Food Research
Open Access
ISSN: 2593-9173
ISSN: 2593-9173
Research Article - (2018) Volume 9, Issue 1
Tef is one of the most stable food crop originated and diversified in Ethiopia. It is highly preferred behind Ethiopian people for consumption as food, especially it has good nutrition content and free of protein known as gluten. In addition, tef used as sources of income so that most of farmers again prefer to produce tef. Thus, the improvement of tef is primary important to maximize productivity. Since tef breeding program was started during late 1950s. From the initial to date several attempts have been done with the satisfactory result. During this period over forty-two varieties were released and the yield was relatively increased. Conversely some little amount of constraints is still not solved. In the same way there are also many challenges in the improvement and development of tef in Ethiopia. Therefore, this review paper was present the progress, achievements and major challenges of tef improvement in Ethiopia.
<Keywords: Eragrostis tef ; Tef improvement; Achievements; Challenges; Tef genomics
Tef is a hugely important crop to Ethiopia, both in terms of production and consumption. In terms of production, tef is the dominant cereal by area coverage and second only to maize in production and consumption [1]. However, it has been historically neglected compared to other staple grain crops, yields are relatively low (around 1.26 tons/hectare), and some farmers under certain conditions sustain high losses which result in reduced quantity of grain available to consumers [2,3].
Tef is mainly serve as staple food, majority of people are preferring grain of tef for consumptions by making Enjera and local beverage. In a country of nearly 90 million people, tef accounts for about 15% of all calories consumed [4]. Tef is highly nutritious, excellent in amino acid composition, its lysine content is higher than that of all cereals except rice and oats [5], it has good mineral content and considerable amount of Iron content when compared with other cereal crops [6]. Tef is free of protein known as gluten which found in wheat, barley and rice, and can causes celiac disease by aberrant T-cell [7]. Tef is not only important for grain consumption but also its straw is highly nutritious and more palatable for livestock compared to straw of other cereals crop specially during dry season.
In Ethiopia tef breeding program is coordinated by Debre Zeit Agricultural Research Center (DAZRC) within the Ethiopian Agricultural Research Institute (EIAR). Debre Zeit Agricultural Research Center (DAZRC) is the centre of excellence for tef research or houses of tef improvement in terms of overall responsibilities for the development and execution of country wide tef research project. Under the umbrella of this institutional framework, the activities, particularly multi location variety trials, of the tef breeding program is carried out at various federal and regional research centers and testing sites, higher learning institutions, and on farmers fields [2].
A primary goal of the national tef breeding program in Ethiopia is to develop high yielding, stress tolerant tef varieties [2]. This depends on the availability of genetic resources with a range of desirable genes within the national germplasm collection, to develop desirable gene combinations in suitable genetic backgrounds. A program has been striving towards the collection, conservation and utilization of the native germplasm resources. From initial to date about 42 tef varieties were released from federal and regional research centres. They have been developed through direct selection from the types grown by the farmers and intraspecific hybridization of lines selected from the germplasm [8]. On experimental station, the yields of these varieties are in the range of 15 to 34 quintals per hectare and on farm level the range was 14 to 20 quintals per hectare [9].
Historical perspective of tef breeding
Scientific tef improvement research in Ethiopia was started in the late 1950s. The period in the late 1950's marked the beginning of tef improvement research at Jimma Agricultural Technical School and later moved to Debre Zeit. Selection of lines from land races was the first attempt made to improve tef. In the years that followed the quest for a hybridization technique continued for several years without success.
According to Assefa et al. the overall history of tef breeding since from initial to date distinguished as five inter-related phases [10]. First phase (1956-1974) was characterized by an emphasis on germplasm enhancement (collection/acquisition, characterization and evaluation, systematic and conservation), genetic improvement relying entirely upon mass and/or pure-line selection directly from the existing germplasm and initiation of induced mutation techniques. In 1972 scientists, briefly, resorted to mutagenesis as a means of creating additional genetic variability. Second phase (1975-1995) marked by the incorporation of intraspecific hybridization into the already preexisting breeding methods following the discovery of the chasmogamous floral opening behavior of tef flowers (from about 6:45-7:30 AM) and thereby the artificial crossing technique [11].
Third phase (1995-1998) featuring initiation of molecular approaches including development of molecular markers and genetic linkage maps, and analyses of molecular genetic diversity. Fourth phase (1998-2003) marked by further incorporation of in vitro culture techniques and inter-specific hybridization along with re-appraisal of induced mutagenesis particularly for lodging and leaf rust disease resistance; and Fifth phase (from 2003 till present), featuring introduction of participatory breeding approaches in the pre-existing overall tef genetic improvement ventures and continued extensive molecular and genomic research approaches [12].
Development of tef genomics
The utilization of the molecular marker technology oers a wide range of novel approaches to enhance the conventional breeding strategies of crops. The use of molecular marker technology in tef breeding is a relatively recent occurrence, and it is still at an infantile stage compared to the conventional breeding. However, there have been significant, on-going eorts on developing the pre-requisites towards the use of the molecular techniques and biotechnological tools to complement and hasten the conventional tef breeding [2].
Molecular markers development: These are markers that work based on naturally arising polymorphism in DNA sequence which are employed for analysis of variation and valuable tools in markers assisted selection and breeding program. So far different markers systems have been studied and applied to tef genetic and genomic studies. Those are random amplified polymorphic DNA (RAPD) [13], amplified fragment length polymorphism (AFLP) [14,15], restriction fragment length polymorphism (RFLP) [16], ISSRs [17] and SSRs [18]. However, there is a low level of diversity detected in cultivated tef using RAPDs and RELPs. In contrast, SSR markers have been shown to be highly polymorphic with in tef germplasm. According to Zeid et al. the overall tef molecular characterization shown that the level of diversity observed has been much improved by the utilization of SSR than RAPDs technology [19].
Development of genetic linkage maps: Genetic maps show the position of the molecular markers and QTLs relative to each other in terms of recombination frequency and are used to find genes responsible for traits of interest [20]. Genetic linkage mapping endeavours in tef started a decade ago and have evolved from the amplified fragment length polymorphism (AFLP) maps in 1999 [21] to simple repeat (SSR) maps in 2011 [18]. Bai et al. developed the first genetic map of tef with an intra-specific cross between the ‘Kaye Murri’ and ‘Fesho’ cultivars and contained 211 AFLP markers in 25 linkage groups [21]. This linkage map was not dense enough to be used in trait mapping due to the low level of polymorphism (6.1%) observed between the two tef varieties. The same group later produced an RFLP linkage map using 116 RILs from the cross of ‘Kaye Murri’ with E. pilosa [16]. This inter-specific cross produced far more polymorphisms; however, the level of polymorphism was still smaller than that of other grasses. The RFLP map Molecular genetic diversity demonstrated better genome coverage (88%) as compared with previous AFLP map (81%) [16]. Another map from an inter specific cross [E. tef (DZ-01-2785) and E. pilosa (30–5)] was developed by Chanyalew et al. by utilizing a combination of dierent marker types namely; AFLP, ISSR, rice EST-SSR markers and tef specific EST-SSR markers. The map was based on 124 F8 RILs and covered 78.8% of the genome [22].
QTL analysis: The first QTL mapping in tef was carried out by Chanyalew et al. who mapped QTLs for grain yield, lodging resistance, seed weight, and shoot biomass and plant height [22]. The authors used 124 F8 recombinant inbred lines (RILs) from an interspecific cross between E. tef and E. pilosa based on AFLP, ISSR, EST-SSR and SSR markers. The percentage of phenotypic variance explained by the QTLs in this study ranged from 12.4% for a QTL associated with grain yield to 63.9% for a QTL associated with days to heading. The second QTL map of tef was constructed by Yu et al. using 94 F8 recombinant inbred lines (RILs) from the same mapping population [22,23]. Ninety-nine QTLs were identified in this study three times more than were identified previously. The most recent QTL map of tef was developed with 151 F9 recombinant inbred lines and PCR-based marker system. This marker system in which 83 QTLs were mapped on 30 linkage groups suffers from small marker density, and the QTLs are not validated and hence are of little use in initiating marker-assisted selection (MAS) in tef [18,22].
Sequence-based diversity: Around the same time, sequencing of single genes and small genomic regions was also employed to measure diversity and genetic relationships. Sequence analysis of non-coding regions of chloroplast DNA, 18S rDNA, and the transcription factor VP1 did not show significant intra-specific variation among six tef cultivars [24]. In addition, two rht1 (reduced height) gene homologs and three sd1 (semi-dwarf) genes were later sequenced for 31 accessions of tef [25].
Genome size and ploidy estimation: Genome size and ploidy have been described as fundamental genetic traits that have important breeding, physiological and genomic implications [26]. One of the methods used to estimate genome size is flow cytometry. This technique involves preparation of aqueous suspensions of intact nuclei whose DNA is stained using a DNA fluorochrome for estimating DNA quantity in a cell nucleus [27]. The technique has been used successfully to determine relative DNA content and ploidy levels in various plants [28]. The genome size of tef was estimated to be between 714 and 733 Mb with flow cytometry based on four cultivars [29] and between 648 and 926 Mb based on ten cultivars [30]. These estimates have been used to validate the genome size estimate obtained from the tef genome sequencing initiative [20].
Genome sequencing and mutation: In recently the genome and the transcriptome of the tef genotype Tsedey (DZ-Cr-37) were sequenced by the Tef Improvement Project at the Institute of Plant Sciences, University of Bern [31]. The draft genome contains 672 Mbp representing 87% of the genome size estimated from flow cytometry and other is sequencing of two transcriptomes; one from a normalised RNA library and another from an unnormalised RNA sequencing (RNASeq) library collected from tef seedlings subjected to various moisture regimes. It will enable future exploration on genetic diversity in tef natural accessions and mutagenised populations [32]. Genome sequencing is especially important for the isolation of homologous copies of each sub-genome for techniques such as Targeting Induced Local Lesions IN Genome (TILLING) which require genome specific primers [33-35]. The tef TILLING project based at the University of Bern in Switzerland is recently initiated with financial support from Syngenta Foundation for Sustainable Agriculture and University of Bern, and scientific collaboration with University of Georgia, FAO/ IAEA Programme, and Ethiopian Institute of Agricultural Research. The main goal of the project is to obtain semi-dwarf tetraploid tef lines which are resistant to crop lodging. So far, the project has generated over 4,000 M2 mutagenized lines and utilized in TILLING [34]. From these an ethyl methane sulphonate (EMS)-mutagenized population, the first semi-dwarf lodging-tolerant tef line, called kegne, was developed and characterized [36].
In vitro techniques: Plant regeneration in tissue culture is a crucial aspect in plant biotechnology as it facilitates the production of genetically engineered plants, the release of disease free plants from meristem cultures and the rapid multiplication of difficult species to propagate them [37]. Establishment of plant cell and tissue culture techniques for tef is a prerequisite to successful plant transformation and regeneration. This was done by using seed culture [38], young leaf base [39], anther culture [40], root culture [41], immature embryos [42]. The report shows that tef plant regeneration through in vitro culture is high regeneration (87-93%) from seed compared to 2-3% from young leaf [39] and root basis [41] mainly due to differences in the explant types, media compositions and genotypes used.
Genetic transformation: Attempts to improve tef using transgenic methods are recent. In the early 1990s, genetic transformation of tef was attempted without success [43]. A decade later it was reported a successful genetic transformation and regeneration of transgenic tef plants obtained by biolistic and agrobacterium-mediated gene transfer [37]. Very recently, Gebre et al. also reported the first stable transformation of tef with GA inactivating gene PcGA2ox under the control of CaMV 35S promoter using Agro bacterium transformation procedure [44]. Despite the works, there is no well defined and reproducible transformation protocol developed for tef and this needs further research. The tef research group at the University of Bern is working to establish a reproducible transformation protocol [32].
Breeding methodology of tef
Breeding methodology of tef employed in tef is generally aimed to the development of high yielding and tolerant variety for different agroecology. Since genetic variation is basic for breeding, the development of tef variety is primarly depends on germplasm enhancement or utilization and conservation of the existed variation or creating variation. This germplam enhancement is through collection and characterization indogeneous germplasm, inra- and inter spesific crossing and induced mutation techniques [2]. Crosses with high genetic variability were considered for further selection and generation advancement. Subsequent segregating populations are handled using the modified pedigree or bulk methods of breeding [2,45] (Figure 1).
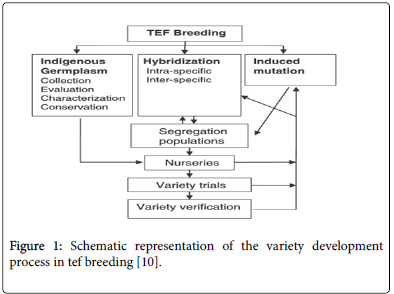
Figure 1: Schematic representation of the variety development process in tef breeding [10].
Despite the indispensable importance of the crop, scientific tef improvement research program in Ethiopia was started only after the late 1950’s to increase production and productivity through genetic improvement and appropriate cultural practices [46]. From initial to date, over 40 improved varieties have so far been released for production by different Federal and Regional Agricultural Research Centers of the country [9]. In addition, several agronomic recommendations have been developed [10]. As tef is native to Ethiopia, the source of genetic variation for effective breeding is limited to landrace collections and crossing of selected parents from the landraces with little or no opportunities of introduction and acquisition of breeding materials and other germplasm from foreign sources [2,3].
Besides to the development of varieties, several agronomic recommendations [10] and scientific study have been done, especially studies on the molecular and biotechnological aspect are provided good knowledge and bench mark for further improvement of tef with the integration of convectional breeding (Table 1).
| No. | Variety | Year of release | From | Altitude | Annual rainfall | Maturity date | Productivity (t/ha) | |
|---|---|---|---|---|---|---|---|---|
| ARC | On farm | research | ||||||
| 1 | DZ-01-99 (Asgori) | 1970 | Debrezeit | 1500-2400 | 300-700 | 80-130 | 1.7-2.2 | 2.4-3 |
| 2 | DZ-01-196(magna) | 1970 | Debrezeit | 1500-2400 | 200-700 | 80-113 | 1.4-1.6 | 1.8-2.2 |
| 3 | DZ-01-354(Enatit) | 1970 | Debrezeit | 1600-2400 | 300-700 | 85-130 | 1.7-2.2 | 2.4-3.2 |
| 4 | DZ-01-787(walankomi) | 1978 | Debrezeit | 2800-2500 | 400-700 | 90-130 | 1.7-2.2 | 2.4-3.0 |
| 5 | DZ-Cr-44 (menagasha) | 1982 | Debrezeit | 1800-2500 | 400-700 | 125-140 | 1.7-2.2 | 2.4-3.0 |
| 6 | DZ-Cr-82(melko) | 1982 | Debrezeit | 1700-2000 | 300-700 | 112-119 | 1.8-2.2 | 2.4-2.8 |
| 7 | DZ-Cr-37(Tseday) | 1984 | Debrezeit | 1500-2200 | 150-200 | 82-90 | 1.4-1.9 | 1.8-2.8 |
| 8 | DZ-Cr-255(gibe) | 1993 | Debrezeit | 1700-2000 | 300-700 | 114-126 | 1.6-2.2 | 2.0-3.0 |
| 9 | DZ-01-974(Dukem) | 1995 | Debrezeit | 1400-2400 | 150-700 | 76-138 | 2-2.5 | 2.4-3.4 |
| 10 | DZ-Cr-358 (ziqala) | 1995 | Debrezeit | 1400-2400 | 150-700 | 75-137 | 1.8-2.4 | 2.1-3.6 |
| 11 | Dz-01-2053(holeta key) | 1998 | Holetta | 1900-2700 | - | - | - | - |
| 12 | Dz-01-1278(ambo toke) | 1999 | Holetta | 2200-2400 | 700-800 | 125-140 | 2.7 | 3.6 |
| 13 | DZ-01-1281(gerado) | 2002 | Debrezeit | 1850 | >600 | 73-95 | 1.6-2.0 | 2.0-2.2 |
| 14 | DZ-01-1285(koye) | 2002 | Debrezeit | 1900-2200 | 300-700 | 104--118 | 1.8-2.5 | 2.4-3.6 |
| 15 | DZ-01-1681(kay tena) | 2002 | Debrezeit | 1600-1900 | 300-500 | 84-93 | 1.6-2.0 | 2.0-2.2 |
| 16 | Dz-01-2054(Gola) | 2003 | Sirinka | 1450-1850 | 660-1025 | 68-100 | 1.6 | 1.1 |
| 17 | PGRC/E205396(Ajora) | 2004 | Areka | - | 900-1200 | 85-110 | 1.0-1.4 | 1.0-3.1 |
| 18 | Dz-01-2675(Degatef) | 2005 | Debrezeit | 1800-2500 | 997-1200 | 112-123 | 1.6-2.0 | 1.8-2.8 |
| 19 | Dz-01-1868(Yilmana) | 2005 | Adet | 200-2600 | >600 | 108 | 1.63 | 2.32 |
| 20 | Dz-01-2423(Dima) | 2005 | Adet | 2000-2600 | >600 | 105 | 1.68 | 2.46 |
| 21 | Dz-01-146(genete) | 2005 | SARC | 1450-1850 | 660-1025 | 78-85 | 1.55 | 2.17 |
| 22 | Dz-01-1821(zobel) | 2005 | SARC | 1450-1850 | 660-1025 | 78-85 | 1.51 | 2.07 |
| 23 | Ho-cr-136(Amarach) | 2006 | Debrezeit | 1600-1700 | 500-850 | 63-87 | 1.2 | 1.3 |
| 24 | DZ-Cr-387/RIL-355(quncho) | 2006 | Debrezeit | 1500-2500 | 300-700 | 80-113 | 2.0-2.2 | 2.4-2.8 |
| 25 | Dz-01-1880(Guduru) | 2006 | Bako | 1850-2500 | 1000-1200 | 132 | 1.4-2.0 | 1.5-2.3 |
| 26 | Acc.205953(Mechere) | 2007 | Sirinka | 1450-1850 | 660-1025 | 79 | 1.79 | 2.06 |
| 27 | Dz-cr-387 RIL#127(Gamechis) | 2007 | Melkassa | 1450-1695 | 690-965 | 62-83 | 1-1.4 | 1.3-2.0 |
| 28 | Dz-01-3186(Etsub) | 2008 | Adet | 1800-2600 | 1230 | 92-117 | 1.6-2.2 | 1.9-2.7 |
| 29 | DZ-01-899(gimbichu) | 2005 | Debrezeit | 2000-2500 | - | - | 1.6-1.8 | 1.8-2.0 |
| 30 | 23-tafi-adi-72(Kena) | 2008 | Bako | 1850-2400 | 1000-1200 | 110-134 | 1.3-2.3 | 1.7-2.7 |
| 31 | Dz-Cr-385 RIL295(simada) | 2009 | Debrezeit | Low | 300-700 | 88 | 1.7-2.0 | 1.8-2.2 |
| 32 | SR-RIL-273(Laketch) | 2009 | Sirinka | 1450-1850 | 660-1025 | 90 | 1.3-1.8 | 2.24 |
| 33 | DZ-Cr-409/RIL50d(Boset) | 2012 | Debrezeit | - | - | - | 1.6-2.0 | 1.9-2.8 |
| 34 | Dz-Cr-438(Kora) | 2014 | Debrezeit | - | - | - | - | - |
| 35 | Acc. 21476A(Workiye)* | 2014 | Sirinka | - | - | - | - | - |
| 36 | DZ-Cr-438 RIL91A (Dagim)* | 2015 | DebreZeit | - | - | - | - | - |
| 37 | Abola* | 2016 | Adet | - | - | - | - | - |
| 38 | DZ-Cr-429 RIL.125 ( negus)* | 2017 | DebreZeit | - | - | - | - | - |
| 39 | DZ-Cr-442 RIL77C (Felagot)* | 2017 | DebreZeit | - | - | - | - | - |
| 40 | DZ-Cr-457 RIL-181( Tesfa)* | 2017 | DebreZeit | - | - | - | - | - |
| 41 | Adet New* | 2017 | Adet | - | - | - | - | - |
| 42 | Areka New* | 2017 | Areka | - | - | - | - | - |
Table 1: Tef varieties released from initial to date (1970-2017) [9,47].'*' Detailed specifications could not be obtained for the most recently released varieties, ARC-Agricultural Research Centre.
Nature of the crop
Seed size: The small size of tef seed poses several problems during sowing, and indirectly during weeding and threshing. Tef is often lost in the harvesting and threshing process because of its size [48]. In their study on the tef value chain, Bekabil et al. [49] reported that tef yields are relatively low (around 1.2 t/ha) and high loss rates (25-30% both before and after harvest). It reduces the quantity of grain available to consumers by up to 50% [49]. This mainly because of the smallness seed. At sowing the very small seed size makes it difficult to control population density and its distribution [3]. Seed size is one of crop character so that improvement may necessary. However, so far, no studies have been done to increase its seed size because of nature of the crop (Figure 2).

Figure 2: Teff and durum wheat grain size. Adapted from Kamies [50].
Shattering problem: Shattering is also causes significant yield loss in Tef production, the crop needs to be harvested on time. Shattering begins right after the crop starts to dry and can still occur during harvesting, gathering, piling and threshing. Results from the field visits revealed that farmers report shattering can account for losses of about 20% of the harvested amount in the Adaa and Bachoo areas and 10% of the harvested amount in the Shashemene area. Under the incidence of rain, the yield loss due to shattering is reported to reach up to 30% in the Dejen area [49].
Difficulty of crossing technique and its floral biology: Tef floret is hermaphrodite with three stamens and a pistil with two stigmas. For many years tef was believed to be cliestogamous, (flowers do not open for self-pollination) [51]. However, Berhe, in 1975 discovered that tef flowers open early at the day break. Then he developed a technique for hybridization of tef for the first time. However, crossing of tef is very cumbersome and time consuming. The main obstacle of tef crossing attributed to its unique pollination habit and size of the flower. The flower of tef is very small, on the average 1- 2 mm and it requires a surgical emasculation using needles and fine forceps and pollination with the aid of a low power stereomicroscope at a magnification of X30-X50 [37].
Besides, flower opening, and self-pollination occurs early at morning, around 6:00-7:00 a.m. Moreover, once the flowers opened, pollen sheds within 30-40 minutes. Therefore, no fresh supply of pollen will be found for hand pollination [52]. Emasculation is essential in tef crossing since there is no male sterility trait found in nature or artificially. It is done by using fine forceps and very tidies and difficult because of flower size. However, Ketema has reported that male sterility may induced by hot water treatment [52]. He observed that the most satisfactory results (high degree of male sterility) were obtained when he treated flower with hot water at 44°C for nine minutes and 48°C for three minutes. But extended treatment beyond 48°C induced female sterility [37]. Even though the existence of such challenges crossing has been forwarded by skilled persons (Figure 3).

Figure 3: Photograph showing crossing of tef under microscope.
Lodging: Lodging is the major constraint to yield increases in tef, while a number of genetic and agronomic factors are involved [53]. Tef plant is so weak in nature and cannot withstand several internal and external factors such as ample application of N2 (Urea) fertilizer application, wind, heavy rain, heavy seedlings and tillers, lack of potassium and phosphorus, high yield per panicles [3,52,54]. Tef ’s estimated yield loss due to lodging can be as high as 30% and reduce the quality of seed in terms of germination capacity, color and nutritional value [2,3,52]. Teklu and Tefera showed that over the years of breeding from 1960-1995, the yield potential of tef increased from about 3300 kg ha-1 to 4350 kg ha-1 when lodging was avoided [55].
Breeding program of tef has been highly tried to combat lodging problem. However, no tef technology (variety) developed until recent years in conventional method. Breeders screened thousands of tef germplasm accessions and found no tef lodging resistant tef types. The challenges of improving lodging resistance by conventional approaches is the lack of sufficient genetic variability in lodging resistance and the apparent unfavorable correlation between lodging resistance and yield related traits such as plant height, panicle length, panicle form, grain and shoot biomass yield [2].
Recently tef breeding program with collaboration of University of Bern applied high throughput technique known as TILLING (Targeting Induced Local Lesions in Genomes) and Eco TILLING with the initial objective of developing dwarf tef plants resistant to lodging (http://www.syngentafoundation.org) [34]. Several semi dwarfs and lodging tolerant candidate lines have been developed and evaluated in the field [32,34]. For instance, the first semi-dwarf lodging-tolerant tef line called kegne, was developed from an ethyl-methanesulphonate (EMS)-mutagenized population [36].
Pre and post-harvest mechanization
Mechanization is the most important and supportive in tef breeding and production systems. So far, a limited attention has been paid to mechanization, processing and storage. The Tef mechanization group, Melkassa Agricultural research station is focusing on this important topic and has developed and tested one of several prototype row planters. But because of its costs most of growers cannot used [49]. In general, the failure to develop affordable and scalable processing technology (especially threshers) are mainly due to nature of the crop, since the seed size is very minutes which is easily lost during sowing to harvesting and packing. In addition, lodging, which makes unfavorable condition for mechanization [3,56].
Limited focus
Tef is an orphan crop which is native to Ethiopia and Eritrea. The attention given to tef research and development is lacked and a few foci has been restricted to only Ethiopian for last century. Initially even government by itself belief that tef is not amenable for improvement because of tef is considered as reliable crop and adaptable to different condition [10]. So that, the support provided to tef research program was not enough and satisfy the gap of research. In addition, since tef is not well kwon internationally as other crop like maize, wheat, sorghum and rice and has not been considered as an important food crop by the international scientific community or funding agencies for a long period of time [49]. Thus, no or little fund to support breeding program of tef [10]. With this situation attempts have been forwarded by a few breeders and other researchers to popularize tef until to a few years before and after a few years focus and support given to tef research and development from government and internationally is improved and with this some satisfactory results have been achieved.
Low capacity building
Limited scientifically-trained human resources, inadequate facilities and infrastructure, and insufficient financial resources are constraints that limit tef research. Breeding is currently the best-functioning approach to tef research, but even this field is under-capacitated. Moreover, there is limited or absent applied research in many areas, such as agronomy, soil, physiology, crop protection, and pre-harvest mechanization.
Tef is among the most widely grown cereals in Ethiopia. The crop is a staple diet of most of the population and the most widely planted by farmers. Tef research was started in early nineteen century and so far, about 42 varieties have been released by using conventional breeding methods and sufficient agronomic recommendation have been developed. In addition to this, several basic studies specially in molecular and biotechnology have been done which are very important for tef improvement. However, there is also many challenges have been existed during this period. Such challenges are nature of crop like smallness of seed size, difficulties of crossing, shattering and lodging; Limited focus, mechanization problems and capacity building. Some of these challenges are relatively solved and some are persisted.