Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Mini Review - (2015) Volume 3, Issue 3
The subpopulation of cancer stem cells (CSCs) in tumor tissues drives tumorigenesis and causes the infiltration of cancer cells to surrounding tissues. CSCs are considered to induce tumor initiation, progression, and relapse. Conventional chemotherapies eliminate bulk tumors; however, CSCs evade most therapies. We recently showed that differentiation of CSCs in hepatocellular carcinoma diminished malignant phenotypes. The best strategy for CSC-targeting therapy is yet unknown, however, it appears that differentiation therapy would be effective for certain types of tumors. In this review, we discuss the characteristics of CSCs and prospective use of differentiation therapy for cancer stem cells.
Keywords: Cancer stem cells, Differentiation therapy, Chemoresistance, Hepatocellular carcinoma, SRY, OCT4, ABC transporter
Cancer stem cells (CSCs) are one of the promising areas of cancer research. Several researchers have focused on the understanding of CSCs after the existence of CSCs in leukemia was described in 1994 [1]. The concept of CSCs explains the phenomenon that malignant status correlates to the existence of CSC-like cells [2]. It is essential to understand the biology of CSC to develop a strategy to eradicate cancer. In various cases, malignant tumor tissues include CSCs, which overcome various anticancer therapies targeting specific proteins despite the temporary effect against the progression of cancer. Given that numerous genes are involved in cellular processes in CSC cell cycles, researchers are yet struggling to find promising cancer therapies.
It has been assumed that CSCs originate from dysregulated behavior of normal tissue stem cells and is limited to a hematopoietic or germ cell lineage [3], however, several types of solid tumors appear to include these aberrant CSCs [4-6].
In fact, non-CSCs and normal cells form CSC-like cells [7]. Several gene sets have been identified to have the potential to transform cancer cells into CSCs or maintain the homeostasis of CSCs. For example, epithelial to mesenchymal transition (EMT) inducers are representative functional markers for the acquisition of a stem-like state in cancer cells [8]. ATP-binding cassette (ABC) transporter proteins in CSClike cells function to secure genomic stability and prevent apoptosis by efflux of cytotoxic agents [9]. It is important to understand the highly complicated signaling mechanism, which sustains the CSC biology.
Inhibiting one of the important pathways for CSC causes the activation of the bypass pathway; therefore, some researchers are seeking a different approach. This approach originates from the ideas of differentiation, which causes leukocyte-initiating cells (LIC) to differentiate into terminally differentiated leukocytes [10,11]. The induction of differentiation has expanded to solid tumors [12]. In this manuscript, we discuss the induction of CSC differentiation based on our recent findings. We have demonstrated the multipotency of CSC in hepatocellular carcinomas (HCC), which implies that CSCs have a potential of multi-lineage differentiation. This strategy would provide novel CSC-targeting therapy.
Characteristics of cancer stem cells
Recent studies have demonstrated that cancer stem cells or cancerinitiating cells with tumor-initiating ability and self-renewal capacity are involved in tumor initiation, metastasis, and relapse [13,14]. It appears that the field of breast cancer contributed considerably to uncovering the biology of CSC [15]. Subpopulations with the cell surface marker CD44+CD24low were identified to be tumorigenic cancer cells [16]. EMT is the critical step for acquiring stem cell features [17]. Plasticity of CSCs between epithelial- and mesenchymal-like CSCs is involved in cancer progression by promoting tumor growth and metastasis [18]. In HCC, multiple CSC markers have been identified, including CD13, CD44, CD90, and CD133 [19-21]. Although it remains unclear when CSCs are generated, premalignant lesions of HCCs have CD44- positive CSC-like populations, which can initiate tumor formation by autocrine IL-6 signaling [22]; this suggests that they are required for tumor initiation of HCCs. With regard to the association between CSCs and metastasis, CSCs were found in the bloodstream of patients with HCC [21,23]. Circulating tumor cells (CTCs) are considered to be undergoing metastasis. CTCs are detected in early stages of prostate cancer in patients [24], and clusters of CTCs are more resistant to apoptosis and an increase in metastatic potential [25]. CTCs have been found in different types of cancer patients [26,27]. When combining all of these reports, it is possible that CSCs appear in relatively early stages of a tumor and contribute to both tumor initiation and metastasis.
Tolerance against chemotherapy in cancer stem cells
Eliminating CSCs from tumor tissue is the most desirable cancer therapy because these cells play important roles in cancer progression. CSCs can survive in severe conditions, although most cancer cells die under these conditions. Shutdown of an important signaling pathway for cancer cells can regress bulk tumor; however, CSCs survive by releasing various agents to the extracellular environment. It has clarified that chemoresistance in CSCs depends on efflux by the ABC transporter gene family, MDR1 (ABCB1) and BCRP (ABCG2), which were originally found in hematopoietic stem cells by the side population (SP) assay [28,29]. The SP assay is the method for identifying CSCs using the efflux of the DNA-binding dye Hoechst 33342 [30,31]. The expression level of the ABC transporter gene correlates with stem cell compartment characterized by SP. MDR1 gene knockout mice were viable and fertile, but showed an increased sensitivity to drugs [32], suggesting that the ABC transporter is important for resistance to chemotherapy. However, the transporter is not critical for stem cell growth. BCRP is described as an essential multidrug transporter against mitoxantrone, doxorubicin, and daunorubicin in breast cancer [33]. To date, the subfamily of ABC transporter genes has been identified, including the ABCA, ABCB, ABCC, and ABCG subfamilies, and play an important role in the efflux of the vast number of anticancer drugs [34]. In melanoma, anticancer drug treatment with vemurafenib and dacarbazine resulted in the selection of ABCB5-expressing cells [35]. The nuclear membrane is important for securing genomic stability. ABCC2 is localized to the nuclear membrane [36] and involved in resistance to several types of chemotherapeutic agents [37-40]. These studies imply that current chemotherapy should focus on certain types of cancer cells, particularly CSCs.
Key signaling pathway in CSCs
Embryonic stem cell (ESC) signatures including OCT4, SOX2, and NANOG, are involved in the development of various types of tumors [41-44]. They play critical roles during embryonic development and are required for pluripotency of induced pluripotent stem (iPS) cells [45]. Because ESC signatures have several downstream genes, their target genes are widespread in oncogenes [46]. Oct4 upregulates CSC markers, and OCT4 and NANOG overexpression promote EMT by increasing the expression of Snail and Slug [47,48]. OCT4 is required for chemoresistance by regulating the ABC transporter [49- 51]. The expression of Sox2 is enriched in the CSC population with high expression levels of CD44 and directly binds to Snail, Slug, and Twist promoters, promoting EMT in pancreatic cancer cells [52]. SOX2 regulates self-renewal and tumorigenicity of CSC-like cells [53]. NANOG has a similar function for CSC and drug resistance, whereas NANOG interacts with CD44 and activates the STAT3 signaling pathway [54]. The reciprocal relationships among ESC genes, micro RNA, and polycomb complexes have also been investigated [55]. These findings provide the basis for the idea that dysregulation of ESC signatures can trigger multi-events important for cancer cells.
CSCs demonstrate that signal transduction pathways, activated in tissue development and homeostasis, have an effect on their state. Notch, Hedgehog (HH), and Wnt signaling pathways have been considered to be targets for overcoming CSC-associated primary or acquired resistance to cancer treatment [56]. In normal stem tissues, HH and Wnt feedback is associated with the proliferation of epithelial stem cells [57]. It is well known that stem cell compartments exist within stem cell niches, and these signaling pathways are important for stem cell niches in tissue homeostasis [58,59]. However, in cancer cells, Wnt/β-catenin signaling has been regarded as an important pathway for upregulating EMT-associated gene expression [60,61]. The HH signaling pathway plays an essential role in tumor initiation, metastasis, and maintenance of cancer stem cells [62,63]. In PDGF-induced gliomas, nitric oxide activity drives Notch signaling via the cGMP/ PKG pathway, promoting CSC-like characteristics [64]. Several studies have demonstrated that TGF-β functions in cancer progression. EMT induced during normal development and oncogenic transformation is mediated by TGF-β and Notch [65]. TGF-β upregulates the expression of EMT inducers including Snail and Slug, promoting EMT and the acquisition of CSC properties [17,66]. These reports suggest that CSCs utilize these signaling pathways for maintenance.
Significance of the sex determining factor
In our previous study, we have shown that SRY directly increases the expression of OCT4, and promotes self-renewal and chemoresistance of HCCs [67]. In rat HCCs, SRY upregulates SGF29, a component of STAGA histone acethyltransferease complex, and promotes HCC progression via c-Myc signaling pathway [68-71]. The increase in SRY expression in Sertoli cells during testes differentiation is a critical step for sex determination, and it induces sex reversal in female mice [72,73]. The timing of SRY expression determines the fate of the bipotential gonadal ridge [74,75], and SRY interferes with the Wnt signaling pathway required for female sex determination [76-78]. The functional region is limited to the HMG box, which has DNA binding and bending abilities to facilitate transcription [79,80]. A mutation within the HMG box provides a male to female specific phenotype [80]. SRY is involved in the gene network of EMT and serves to stabilize cell plasticity [81]. Several studies have demonstrated an association between SRY and tumor development [68,82,83]. Males have a higher cancer risk than females [84]; therefore, it is possible that SRY contributes to gender disparity in cancer occurrence and progression.
Potential of differentiation therapy
Determining the direction of differentiation would be helpful for certain types of disease, for which a therapeutic strategy has been found. Although it is possible that the differentiation from iPS cells would generate transformed cells, it does not have the potential to transform cells using a direct differentiation strategy. Direct differentiation could be a breakthrough for overcoming neurological disease. Because of four major reprograming factors, Oct4, Sox2, Klf4, and c-Myc, fibroblasts are transdifferentiated into neural progenitor/stem cells [85]. The minimal set of transcriptional factors, Mash1, Nurr1, and Lmx1a, are sufficient for inducing dopaminergic cells from mouse and human fibroblasts [86]. In addition to these methods, terminally differentiated hepatocytes are converted to functional neuronal cells by the combination of Ascl1, Brn2, and Myt1l [87]. Several studies have shown neural differentiation in vitro, and these techniques would provide a novel strategy for Parkinson’s or Alzheimer’s disease patients.
Some studies have shown that the significance of differentiation of cancer cells into normal tissue cells, which contributes to tumor regression, is induced by some factors, including nutrient conditions, chemicals, and genetic processes. In prostate cancer, androgen deprivation and chemical compounds induced differentiation into neuroendocrine cells [88,89]. Tumor differentiation factors produced by the pituitary gland and secreted into the bloodstream induce the differentiation of prostate and breast cancer [90-92]. In HCC, knockdown of Yap restores hepatocyte differentiation and tumor regression [93], suggesting that restoration of normal function or differentiated phenotypes in cancer cells are related to tumor suppressive function.
Accumulating research has shown the possibility of a differentiation capacity of CSCs. In 1980, differentiation therapy had been adapted to leukemia CSCs (L-CSCs) to induce retinoic acid (RA)-induced differentiation in certain hematopoietic cells [10,11]. It has been well identified that RA, the active form of vitamin A, plays an important role in the induction of neuronal differentiation in mouse embryonal carcinoma lines, P19 and F9 [94-96]. We have recently demonstrated the possibility of differentiation therapy against purified CSCs from HCCs, which differentiated into hepatocytes and neuronal cells induced by BMP4 and RA, respectively [67]. We have confirmed that in the presence of lower concentrations of serum, a high differentiation efficiency is observed, suggesting that lower nutrients affect the status of CSCs. The differentiation capacity of CSCs is well characterized in several types of tumors. For instance, CD133 positive CSCs in HCCs differentiate into hepatocytes induced by BMP4 [97]. The SP fraction of a C6 glioma cell line can produce neuronal and glial cells [98]. Spheroids from ovarian carcinomas show stem cell like properties and have translineage differentiation capacities [99]. Reprogramed sarcoma cells in the pluripotent state, induced by defined reprograming factors, induce terminal differentiation into multiple tissue types, although it is a different germ layer [100]. Glioblastoma stem-like cells generate tumor endothelium, suggesting that they control their plasticity for tumor progression [101]. Subpopulations of tumor tissues may eventually acquire stem-like properties via certain nuclear reprogramming factors or some signaling pathways, which in turn, could be therapeutic targets. In summary, two types of therapy have been considered to be effective for tumor tissues (Figure 1). Differentiation therapy could be valuable for CSCs. Matured/differentiated cancer cells may be targets for direct differentiation via the set of lineage-specifying transcriptional factors and conventional chemotherapy.
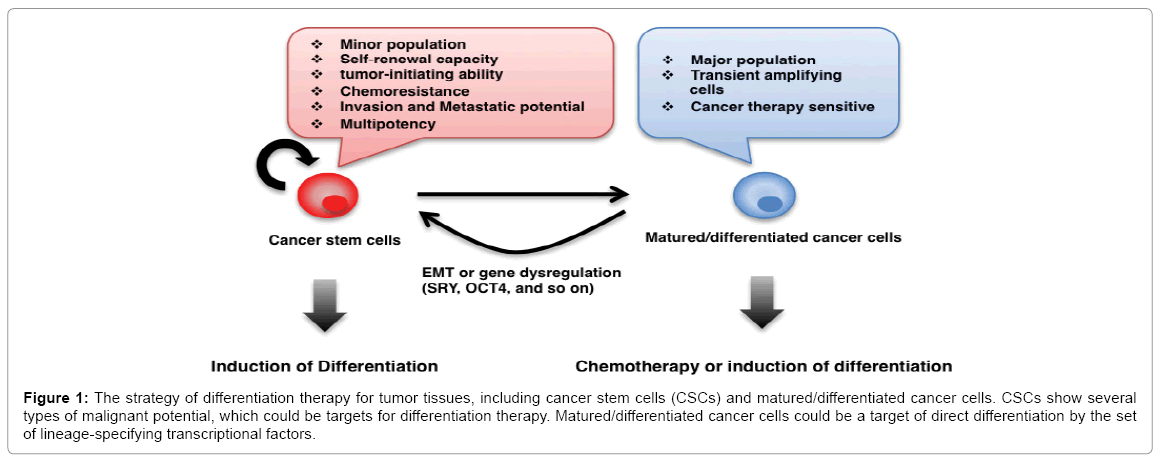
Figure 1: The strategy of differentiation therapy for tumor tissues, including cancer stem cells (CSCs) and matured/differentiated cancer cells. CSCs show several types of malignant potential, which could be targets for differentiation therapy. Matured/differentiated cancer cells could be a target of direct differentiation by the set of lineage-specifying transcriptional factors.
Our studies show the high potency of differentiation therapy against the CSC population in HCC. However, at present the differentiation is limited in some tumors and this technique is yet poorly developed. Furthermore, we have no options in the late stages of cancer such as drug refractory and metastasized cancers, which contain considerable CSCs. Therefore, the differentiation strategy requires improvement.