Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2016) Volume 7, Issue 4
Reproduction of Mugil cephalus of the Pacific coast of Mexico was studied. Fish were captured with gill nets and cast nets; they are a common low priced product for local consumption. The study of the reproduction period and ages of first maturity helps manage the fishery. Fish were obtained from local commercial fishery from August to December 2007, January to March 2008 and November 2012 to October 2013. Size and weight, sex and gonad maturity were registered. The male:female ratio was 0.88:1. Mature organisms occurred all year round. Sexual maturation (L50) of males and females was observed at a mean size of 34.0 cm in males (4.64 years of age) and 35.0 cm in females (4.98 years of age). First maturity length (L25) was both 30.0 cm in males and females corresponding to 3.4 years of age in both cases. The allometric relationship with the hepatosomatic index was LW=4.00·10−3 · TL2.771 (r2=0.849). Condition factor indexes of Clark and Safran EW showed a maximum increment during June, August and December; Fulton and Safran TW in July and September to November. The gonadosomatic index showed its highest values from November to January. The hepatosomatic index reached its maximum values in June, July and August. The gastric repletion index reached its highest values in June, February and October. The mean oocytes diameter was 0.38 mm (range 0.22 to 0.52 mm, standard deviation=0.13). Fecundity ranged from 1’422,076 to 1’747,736 oocytes in females between ages 3 and 12 years old, and mean relative fecundity was 2,830 oocytes·g-1 (1,500 to 2,900 oocytes·g-1). This study is the base line for the fishery management of M. cephalus in Central Mexican Pacific, where the main regulations need information on the first maturity size and reproductive season.
Keywords: Fecundity; Maturity period; Fish reproduction; Gonadosomatic index; Hepatosomatic index; Gastric repletion index; Condition factor
The striped mullet Mugil cephalus Linnaeus 1758 (Figure 1) has a worldwide distribution between 42°N and 42°S [1]. Through the biogeographic areas and provinces of the American continent, the only exceptions are the cold temperatures of the Northeast Pacific in the Pacific Province and the Magellanic Province in South America [2]. In the Western Atlantic it distributes from Nova Scotia to Argentina including the Gulf of Mexico [3]. In the Eastern Pacific it distributes from California to Chile, including the Gulf of California and Galapagos Islands [4].

Figure 1: Stripped mullet Mugil cephalus.
New research made by Whitfield et al. establishes that M. cephalus is a cosmopolite species that flourishes in a high variety of habitats and that can be considered as a eurytopic species complex, and could be used as a biological marker in the health levels of different ecosystems where it inhabits [5].
This species is important for the meat consumption and the “roe” (female mature gonads), which reaches a higher price than the meat: the roe is $300.00 Mexican pesos per kilogram ($18.00 US dollars) and the meat $30.00 Mexican pesos ($1.80 US dollars) per kilogram.
M. cephalus ranks in 22nd place in Mexican fisheries, with a capture of 12 280 [6]. This species has been studied in many parts of the world where well established fisheries are located. In the case of Mexico, analysis has also been carried out on this fishery and biological aspects [7-20]. However, most of the studies in Mexico have been carried out in the Tamiahua lagoon, Tamaulipas, on the Atlantic Ocean, or in Mazatlán, Sinaloa and Nayarit on the northern Mexican Pacific Ocean.
In the coast of Jalisco and Colima, M. cephalus does not reach a high catch volume as in these other places, but it is part of a multispecific fishery and important to know the health status of their populations. Traditionally Mugil curema is fished in a higher amount in the coasts of Jalisco and Colima, than M. cephalus. In 2014, M. curema was fished up to 626 tons (79% of the total Mugilidae species), and 167 tons of M. cephalus (21% of the total Mugilidae species) [6]. Therefore M. curema has been analyzed and some studies of its population dynamics were done [21-26].
The objectives of the present study were to analyze monthly frequency of the gonadic maturity stages and massive spawning period; monthly values of the gonadosomatic and hepatosomatic index; monthly values of the gastric repletion index; monthly values of the condition factor of Fulton, Clark and Safran; values of total and relative fecundity; and to compare our results to those reported by other authors [27-29].
These studies will give a solid background for closed seasons and gill net mesh sizes, based on the minimum reproductive size. These fishing measures will allow the species to reproduce at least once, protecting the fishery from overexploitation.
From August to December 2007, January to March 2008 and November 2012 to October 2013, M. cephalus samples were monthly collected in the Cuyutlan Lagoon, Colima, Mexico (103°57’-104°19’ W and 18°57’-19°50’ N) and in Cruz de Loreto Lagoon, Jalisco, Mexico (105º27’-105º33’ W and 19º58’-20º05’ N) (Figure 2). The fishing gears were gill-nets of 2.0, 2.5, 3, 3.5 and 4 inches mesh size (5.08, 6.35, 7.62, 8.89 and 10.16 cm). Total length was measured to the nearest millimeter (TL, cm) from the snout tip to the caudal fin extreme in 262 organisms (fishermen deliver this species intact); the total (TW, g) and eviscerated (EW, g) weight of 784 specimens (weighed to the nearest 0.1 g) were measured.
Figure 2: Study area of Colima and Jalisco.
The function W=a.Lb was used to obtain weight-length relationship and sex was recorded macroscopically for each specimen. Sexual maturation was determined in visu on fresh organisms taken to the laboratory the same day they were caught. Sokolov and Wong, Holden and Raitt, Aboussouan and Lahaye and Espino-Barr suggest a scale to determine sexual maturity and describe the stages as follows [30-33]:
• Phase I: Sexually immature organisms, in which sex cannot be distinguished, the gonads are very fine filaments.
• Phase II: Organisms have not yet matured sexually, the testis start to develop and are light colored and ovaries are pale pink, and oocytes cannot be observed.
• Phase III: Sexual maturity begins; sexual glands can be perfectly identifiable. Oocytes are beginning to form and are opaque, the color of the ovaries start to turn dark pink, testis also show darker and opaque color.
• Phase IV: Mature, sexual glands are well developed, ovaries are rose-orange, oocytes are big and transparent, and testes are whitish.
• Phase V: This stage corresponds to the spawning, both, ovules and sperm that are expelled if the visceral cavity is pressed, gonads show and intense blood supply and both ovaries and testes show brighter colors.
• Phase VI: Corresponds to the post-spawning, sexual products have been expelled, both ovaries and testes are empty, gonads coloring tend towards a dark pink.
Sparre and Venema suggest that the first spawn length is calculated as the 50% of the accumulative frequency (L50) of phases IV and V of the sexual maturity scale mentioned above, considering that the lowest spawning length (L25) is also registered, to compare with data reported in other studies [34,35]. This kind of analysis are carried out for both males and females and both (L25) as (L50) are useful because deliver information on the stages close to the reproduction. Gaertner and Laloe and Sparre and Venema represent this function by the equation [34,36]:
Hp=1/[1+ea+b·TL],
where: HP=the percentage of mature organisms (males or females) and a and b are constants.
Transforming this equation logarithmically, we obtain:
ln 1(1/Hp-1)=a-b · TL, and the length at which 50% of the population is sexually mature (L50) corresponds to: L50=a/b.
To include L50, the original equation is modified:
Y=1/[1 + a (1-TL/L50)]
The minimum TL of first spawning (L25) was also recorded to be compared with other authors’ findings.
The formula mentioned by Rodríguez-Gutiérrez to calculate the gonadosomatic index (GSI) for males and females of M. cephalus [35], considers the gonad weight (GW) in relation to the fish total weight,
i.e., GSI=100·GW/TW (TW=total weight).
To measure physical fitness of fish, we obtained the condition factor
K=(EW·TL-3)·100,
K=(TW·TL-3)·100 and
a=TW·TL-b and
The hepatosomatic index (HSI), expressed as the percentage of liver weight (LW) with respect to the total weight was calculated as:
HSI=100·LW/TW [35].
The stomach repletion index is the relation between the stomach weight and the body weight, calculated individually and averaged monthly.
The gravimetric method was used to calculate total fecundity (F) and relative fecundity (Fr) using the weight of 45 females in phase V of gonadic maturity. To estimate F, two subsamples of 0.1 g were obtained of each individual and put in modified Gilson fluid for preservation [37]. All oocytes were counted with the help of a stereoscopic microscope and measured with an ocular micrometer.
The following formula was used to determine fecundity:
F=n ? Gi/gi,
where n=number of oocytes in the subsample; Gi=weight of the gonad (g) and gi=weight of the subsample (g) [31].
The relationship between fecundity and total length and weight was calculated with the formula
F=a · xb,
where x=individual weight or length, a=intercept or initial number of oocytes, b=slope or oocyte number changing rate.
The data obtained by Espino-Barr et al. in the study of otoliths, were used to obtain –for each age- the relations between TL, TW, LW, testis weight (TeW), ovary weight (GW), and fecundity [19].
M. cephalus sex cannot be differentiated by their body morphology, so organisms have to be opened and eviscerated; 262 individuals were sexed. The use of gill nets of different size (2.0-4.0 μn) in commercial fishery allowed catching individuals of various age groups.
Once organisms begin their gonadal maturity, sex identification is performed quite easily. Ovaries are elongated and during the season close to spawning, oocytes are easily observed, with a bright yellow pink coloration.
The relationship from each age group to the values of the total length (TL, cm), total weight (TW, g), eviscerated weight (EW, g), liver weight (LW, g), gonad weight (TeW, g and GW, g), and fecundity (number of oocytes) are shown in Table 1.
| Age | TL (cm) | TW (g) | EW (g) | LW (g) | TeW (g) | GW (g) | F (eggs) |
|---|---|---|---|---|---|---|---|
| 0 | 15.62 | 40.000 | 33.000 | 0.767 | - | - | - |
| 1 | 20.43 | 89.000 | 74.000 | 1.604 | - | - | - |
| 2 | 24.71 | 156.000 | 130.000 | 2.706 | - | - | - |
| 3 | 28.53 | 239.000 | 198.000 | 4.018 | 1.440 | 2.379 | 1,422,076 |
| 4 | 31.94 | 333.000 | 277.000 | 5.481 | 2.966 | 4.216 | 1,485,136 |
| 5 | 34.98 | 436.000 | 362.000 | 7.037 | 5.308 | 6.684 | 1,537,943 |
| 6 | 37.69 | 544.000 | 451.000 | 8.640 | 8.557 | 9.756 | 1,582,684 |
| 7 | 40.11 | 653.000 | 542.000 | 10.253 | 12.743 | 13.374 | 1,620,990 |
| 8 | 42.26 | 762.000 | 633.000 | 11.836 | 17.799 | 17.426 | 1,653,846 |
| 9 | 44.18 | 869.000 | 722.000 | 13.374 | 23.653 | 21.827 | 1,682,328 |
| 10 | 45.90 | 973.000 | 808.000 | 14.855 | 30.202 | 26.489 | 1,707,202 |
| 11 | 47.42 | 1,072.000 | 890.000 | 16.248 | 37.203 | 31.245 | 1,728,711 |
| 12 | 48.79 | 1,165.000 | 967.000 | 17.571 | 44.642 | 36.097 | 1,747,736 |
Table 1: Length (TL, cm), total weight (TW, g), eviscerated weight (EW, g), liver (LW, g), testis weight (TeW, g), ovary weight (GW, g) and fecundity (number of oocytes) for each age group (years).
Testes are cream colored long triangular tape type and smaller than ovaries.
Oocyte diameters were 0.38 mm (from 0.22 to 0.52 mm ± 0.12 mm standard deviation: SD). The f ecundity values oscillated from 1’422,076 to 1’747,736 oocytes in females from 3 to 12 years of age, length from 28.5 cm to 48.8 cm, and total weight 239 g to 1,165 g (Table 1). Relative fecundity showed average values of
Fr=2,830 oocytes?g-1 (ranging from 1,500 to 2,900 oocytes?g-1).
The sample size was of 262 organisms of M. cephalus, of which 123 were female (46.95%), 109 male (41.60%), and 30 undetermined (11.45%). The male: female proportion was 0.88: 1.0.
According to the values obtained of the monthly frequencies for the gonadal maturity scale, it was observed that immature male organisms corresponding to Phase II prevailed from March to August in 100% with high values also in September and October. Immature females were present with more than 50% in February, May, August and December (Figures 3a and 3b). Phase III: Maturing, females were not present and males showed up during January and November, while phase IV, mature, was observed in July, September and October for females and in January for males. Phase V or spawning stage was observed in females in November, January and February. Males were in phase V in September and from November to January. Phase VI, post-spawning, females were observed during all year round, with high values in March, April, June, July, September and October, while males were present in February, October, November and December (Figures 3a and 3b).
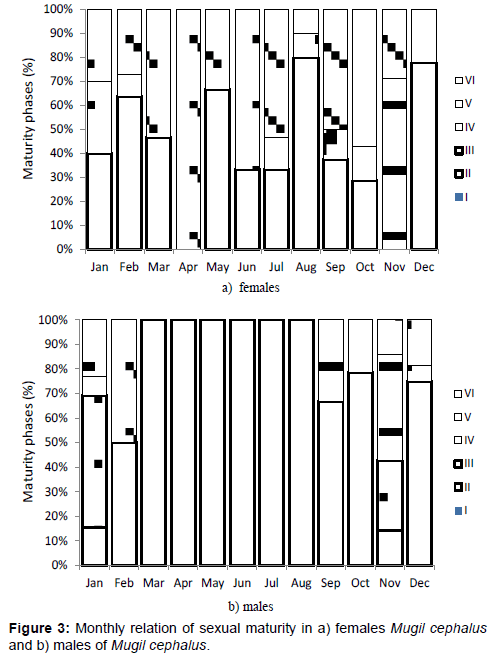
Figure 3: Monthly relation of sexual maturity in a) females Mugil cephalus and b) males of Mugil cephalus.
Length at first maturity was L25=30.0 cm in females (Figure 4a) and in males (Figure 4b), corresponding to 3.4 years of age. First reproduction length was L50=35.0 cm in females (Figure 4a) and L50=34.0 cm in males (Figure 4b), which correspond to 4.98 and 4.64 years of age, respectively.
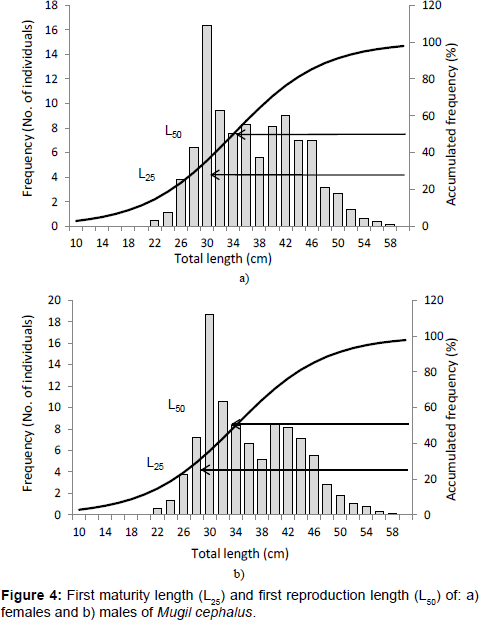
Figure 4: First maturity length (L25) and first reproduction length (L50) of: a) females and b) males of Mugil cephalus.
The gonadosomatic index (GSI) showed the highest values in November and January for total length and total and eviscerated weight (Figures 5a and 5b), followed by a very light increase of GSI values during September and October only observed for total length and total weight. GSI values were very low the rest of the year.
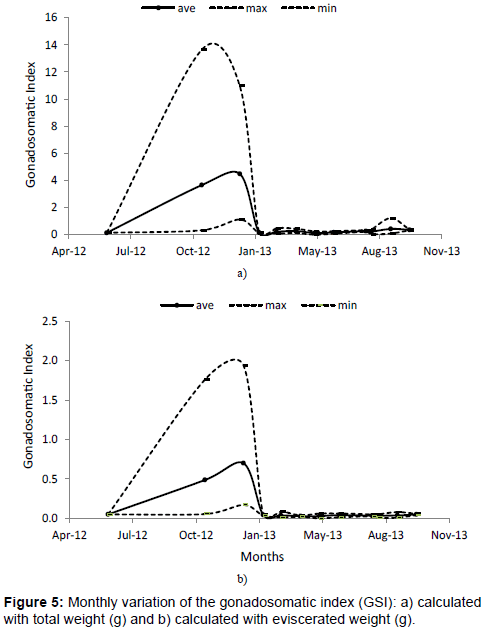
Figure 5: Monthly variation of the gonadosomatic index (GSI): a) calculated with total weight (g) and b) calculated with eviscerated weight (g).
We obtained the following allometric relationships of the hepatosomatic index (HSI) LW=4.00·10−3 · TL2.771 (r2=0.849). The index b shows that, in terms of length, the liver weight is lower than a cubic proportion, which results in a negative allometric growth of the fish and a decrease of its fatty reserves as it ages. HSI variations are shown in Figures 6a and 6b; maximum values are observed in May, June and July and lower values in February and March.
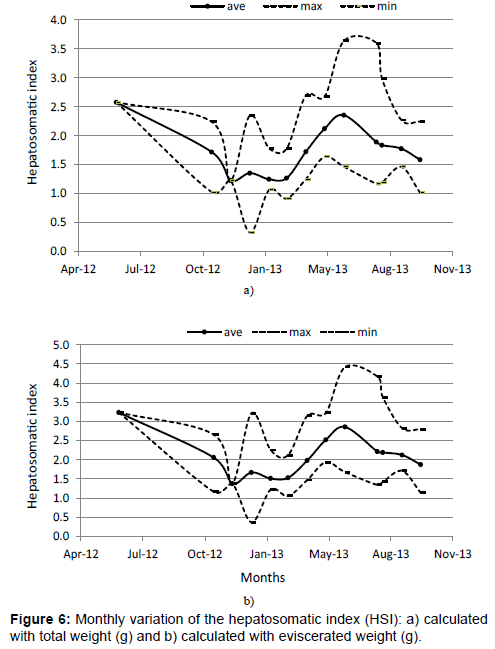
Figure 6: Monthly variation of the hepatosomatic index (HSI): a) calculated with total weight (g) and b) calculated with eviscerated weight (g).
The differences in the stomach repletion index (Figures 7a and 7b) showed higher values during June, February and October; lower values are observed in December, January and September.
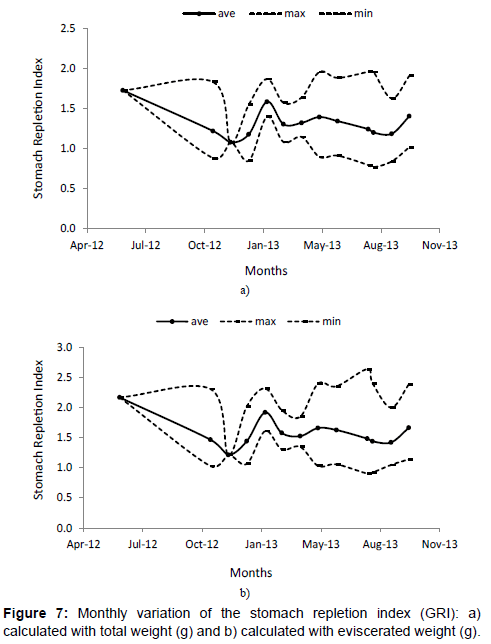
Figure 7: Monthly variation of the stomach repletion index (GRI): a) calculated with total weight (g) and b) calculated with eviscerated weight (g).
Figure 8 shows data of the condition factor; the highest values are obtained in January, August and December for Clark index and Safran EW. For Fulton index and Safran TW the highest values correspond to July, September, October and November.
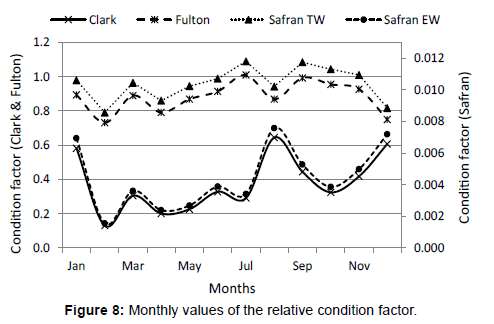
Figure 8: Monthly values of the relative condition factor.
The highest length growth rates of M. cephalus calculated by Espino- Barr et al. are in groups zero and three years of age, a second period corresponds between ages 4 and 7 years, and a third period between ages 8 and 12 years, which show the lowest length growth rate [19]. As length growth rate starts to decrease, total weight, gonad weight and fatty reserve index start to rise. In this way, two main seasons were registered in the life cycle of M. cephalus: first, from ages zero and three when most of the energy obtained through food is used to increase its length (reducing depredation and interspecific competence), and second from ages four to twelve, when this energy is oriented to form the sexual products and fatty reserves (Figure 9) [33,38,39].
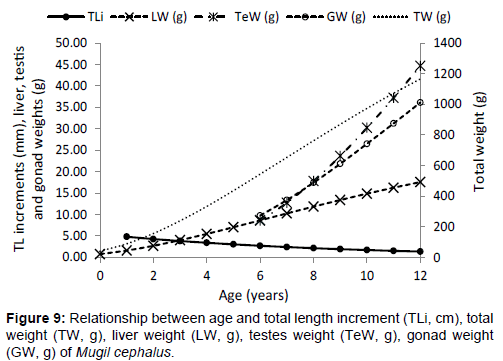
Figure 9: Relationship between age and total length increment (TLi, cm), total weight (TW, g), liver weight (LW, g), testes weight (TeW, g), gonad weight (GW, g) of Mugil cephalus.
Sexual proportion was 0.88:1 male:female, values slightly higher of 1:1.1 male:female were found for M. cephalus in Tamiahua lagoon in the Atlantic sea [40].
During all year round M. cephalus specimens were observed in post spawning phase (Figure 3), which indicates that during every months mature organisms are present and that the reproduction carries out during every month of the year. However, most of the organisms in spawning phase occur during November, December and January. In the case of M. cephalus of Tamiahua, Veracruz, spawning occurs during autumn and winter, coinciding with the northern winds [40].
The highest first reproduction size of M. cephalus (L50) was 42.52 cm in males and females which corresponds to an age of three years, in the Gulf of México (Table 2), followed by Briones-Avila who obtained values of 38.00 cm in males and females of specimens from Nayarit and Sinaloa in the Mexican Pacific [7,41,42]. Render et al. found sizes of 36.95 cm in organisms of Louisiana in USA [43]. Hubbs reported lengths of 33.00 cm in males and 35.00 cm in females with two years of age in Florida, USA [44]. Organisms of smaller lengths have been reported by Arnold [45,46], Jacot et al. in the coasts of Florida, and by Oren in Texas and Florida, USA [47]. In the present study males reached first reproduction length at 34.0 cm (4.64 years old) and females at 35.0 cm of total length and 4.98 years of age
| Males | Females | Author | ||||
|---|---|---|---|---|---|---|
| Area | TL (cm) | Age (yr) | TL (cm) | Age (yr) | ||
| Florida (E ), USA | 30.36 | 2 | 30.36 | 2 | Jacot[45] | |
| Florida, USA | 33.00 | 2 | 356.00 | 2 | Hubbs[44] | |
| Gulf of Mexico | 24.00 | - | 25.80 | - | Arnoldand Thompson [46] | |
| Gulf of Mexico | 42.52 | 3 | 42.52 | 3 | Márquez-Millán [7] | |
| Texas, USA | 20.00-35.50 | 25.00-35.00 | Oren [47] | |||
| Florida (W), USA | 23.00-29.00 | 3 | 24.00-31.00 | 3 | Oren [47] | |
| Florida (E ), USA | 23.60 | 2 | 25.00 | 3 | Oren [47] | |
| Louisiana, USA | 36.95 | 36.95 | Render[43] | |||
| Gulf of Mexico | 28.00 | 3 | 29.90 | 3 | Ibáñez-Aguirre and Gallardo-Cabello [40] | |
| Nayarit, Mexico | 38.00 | - | 38.00 | - | Briones-Ávila [41] | |
| Sinaloa, Mexico | 38.00 | - | 38.00 | - | Briones-Ávila [42] | |
| Mexican Central Pacific | 30.00-34.00 | 3.4-4.64 | 30.00-35.00 | 3.4-4.98 | Thisstudy | |
Table 2: First maturity (L25) and reproduction (L50) length of Mugil cephalus in different places.
The gonadosomatic index (GSI) reached the highest values in November and January in this study (Figures 5a and 5b). Similar spawning seasons to the present study (November to February), were found in Tamiahua lagoon Veracruz, Mexico [40], in North Carolina and to lower Florida [48], in Hawaii [49], in southwestern Taiwan [50], west coast of Taiwan [51], northeastern coasts of Taiwan [52], south west of Bay of Bengal [53]; in all these areas the massive spawning period of M. cephalus is from the end of autumn and during winter.
However in other parts of the world, spawning of M. cephalus occurs mainly during summer, as is the case of Delaware Bay [48], Natal estuaries of South Africa [54] and Primorye, Russia [55]. In other areas the maximum reproduction of M. cephalus takes place during summer and early winter, as occurs in France [56] and the Balear Island of Menorca, Spain [57]. Also spawning of M. cephalus can occur during spring and summer as is the case of the east and west coast of Australia [58] (Table 3).
| Country | Locality | Period | J | F | M | A | M | J | J | A | S | O | N | D | Author |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Australia | eastcoast | March-July | - | - | x | x | x | x | x | - | - | - | - | - | Kailola[58] |
| Australia | west coast | March-September | - | - | x | x | x | x | x | x | x | - | - | - | Kailola[58] |
| South Africa | Natal estuaries | May-August | - | - | - | - | x | x | x | x | - | - | - | - | Marais[54] |
| Russia | Primorye | May-September | - | - | - | - | x | x | x | x | x | - | - | - | Novikov [55] |
| France | France | July-November | - | - | - | - | - | - | x | x | x | x | x | - | Keith andAllardi[56] |
| Spain | Minorca (Balearic Archipelago) | August-November | - | - | - | - | - | - | - | x | x | x | x | - | Cardona [57] |
| India | southwestBay of Bengal | October-December | - | - | - | - | - | - | - | - | - | x | x | x | Jeyaseelan[53] |
| Taiwan | northeasterncoasts | December-January | x | - | - | - | - | - | - | - | - | - | - | x | Hsu [52] |
| Taiwan | West coast | december | - | - | - | - | - | - | - | - | - | - | - | x | Shung[51] |
| Taiwan | southwestern Taiwan | October-February | x | x | - | - | - | - | - | - | - | x | x | x | Chang [57] |
| Hawaii | Hawaii | December-February | x | x | - | - | - | - | - | - | - | - | - | x | Honebrink [49] |
| USA | North Carolina to lower Florida | October-February | x | x | - | - | - | - | - | - | - | x | x | x | Scotton[48] |
| USA | Delaware Bay | June-August | - | - | - | x | x | x | - | - | - | - | Scotton[48] | ||
| Mexico | TamiahuaLagoon, Veracruz | December-February | x | x | - | - | - | - | - | - | - | - | x | x | Ibáñez-Aguirre and Gallardo-Cabello [40] |
| Mexico | Central Pacific (Jalisco and Colima) | - | x | - | - | - | - | - | - | - | - | - | x | x | Presentstudy |
Table 3: Spawning seasons of Mugil cephalus in different places.
The hepatosomatic index obtained in this study was
b=2.771 (r2=0.849),
which indicates a negative allometric growth; since fish decrease their fatty reserves as they grow older. In the case of M. cephalus in the Tamiahua lagoon, Veracruz, the relationship obtained for the hepatosomatic index showed an isometric growth b=3.0, which shows that the increments of the liver weight are directly proportional to the cubic length [40].
Monthly values of hepatosomatic index (Figures 6a and 6b) showed that the liver accelerates its activity of reserving fatty acids during the periods before spawning; therefore, their weight increases considerably. The highest activity of fatty acid reserves is in June, July and August and starts to decrease in February and March after the spawning period of November, December and January. A similar phenomenon was observed in Tamiahua, where the fatty acids accumulation occurred in previous spawning periods [40]. However the largest accumulation of fatty acids was in the abdominal cavity instead of the liver. Ibáñez and Gallardo-Cabello reported this same observation in M. cephalus in Tamiahua, where the accumulation of glycogen is so intense in the periods previous spawning, that the stored reserve of hepatic acids is insufficient, which shows a sub estimated hepatosomatic index of the fatty acids reserve in the animal’s body [40].
Variations in the stomach repletion index (Figures 7a and 7b) showed higher values during June, February and October, which are the months previous to spawning and higher accumulation of fatty acids reserve in the liver. Once the spawning has occurred, in the months of November, December and January, values of the gastric repletion values decline significantly.
Figure 8 shows the values of the condition factor; the highest values are obtained in January, August and December for the Clark index and Safran with eviscerated weight values (EW). For the Fulton index and Safran with total weight (TW) the highest values correspond to July, September, October and November. In all cases the highest were in the months previous to spawning or at its beginning. Similar values were obtained in Tamiahua where the highest value of the condition factor increments before the spawning period and decreases at its end; in May is when the higher recovery of the condition factor occurs [40].
Table 4 shows the fecundity values of M. cephalus in different countries; the highest values correspond to the Black Sea, where it can reach values over 7 million of oocytes [47]. Also Berg reports in the same area 7 million of oocytes in organisms of 52.00 cm of total length and 13 years of age [47]. In the Hawaiian Islands, Keith et al. found a maximum fecundity of 7 million of eggs; in Mauritania [59], Brulhet found a maximum of 6 million, and Popescu of 5 million in organisms from the Danuvian delta [47]. Grant and Spain reported 4 million 800 thousand in Australia [60]. Values of fecundity of 3 million 790 thousand eggs were reported in SW Korea by Yang and Kim, and of 3 million in Taiwan by Tung and Hsu [52,61,62]. Solís found a maximum value of fecundity for M. cephalus in Tamaulipas, Mexico in the Atlantic Ocean of 2 million 919 thousand oocytes in females of 48.00 to 56.00 cm of total length [63]. In Australia, Thomson and Kesteven reported 2’781,000 and Tosh 2 million and a half in females of M. cephalus [47,64,65]. In Mauritania and Senegal, Landret found a value of 2’322,400 oocytes [47]. In our study values of 1’582,684 to 1’747,736 oocytes in females of the coast of the Central Mexican Pacific (Jalisco and Colima) were found in organisms of 37.70 to 48.80 cm of total length and 6 to 12 years of age. The lowest values were reported by Ibáñez and Gallardo-Cabello of 898,512 oocytes in females of 38.00 cm total length and 6 years of age, from the Tamiahua lagoon, Veracruz, Mexico, although these authors mention to have found fecundity values of 1’483,056 oocytes in females older than 6 years of age [40]. At last, Shehadeh et al. found the lowest value reported in the Table 4, of 795,000 oocytes in M. cephalus females from Hawaii [47].
| Author | Area | Fecundity (oocytes) | Organism size |
|---|---|---|---|
| Thomson [65] | Australia | 1’275,000-2’781,000 | - |
| Kesteven[64] | Australia | 1’275,000-2’781,000 | - |
| Grant & Spain [60] | Australia | 1’600,000-4’800,000 | - |
| Tosh[47] | Australia | 2’000,000-2’500,000 | - |
| Jacob &Krishnamurthi[47] | India | 1’320,000 | 50 cm (TL) |
| Tung[62] | Taiwan | 700,000-3’000,000 | - |
| Hsu [52] | Taiwan | 700,000-3’000,000 | |
| Yand& Kim [47] | SW Korea | 3’790,000 | 78.7 cm, 5 yearsold |
| KeithyAllardi [56] | Francia | 500-2,800/gram | - |
| Popescu[47] | Black Sea: Danubian delta | 5’065,800-5’085,440 | - |
| Nikolskii[47] | Black Sea | 3’089,000-7’206,000 | - |
| Berg[47] | Black Sea | 5’000,000-7’000,000 | 52 cm, 13 yearsold |
| Brulhet[47] | Mauritania | 4’000,000-6’000,000 | - |
| Landret[47] | Mauritania and Senegal | 2’322,400 | 50 cm FL |
| Shehadeh[48] | Hawaii | 340,000-795,000 | inducedspawning |
| Nash [47] | Hawaii | 1’000,000 (effectivelyreleased) | - |
| Keith [47] | Hawaii | 5’000,000-7’000,000 oocytes | - |
| Solís [63] | Tamaulipas, Mexico | 1’341,000-2’919,000 oocytes (6,510 oocytes/g) | 48 and 56 cm (TL) |
| Ibáñez-Aguirre and Gallardo-Cabello [40] | Veracruz, Mexico | 405,767-898,512 (1,680 oocytes/g, 680-4,776) | - |
| This study | Central Mexican Pacific | 1’422,076-1’747,736 oocytes (2,830 oocytes·g-1, 1,500 - 2,900 oocytes·g-1) | 28.5 cm to 48.8 cm (TL) |
Table 4: Fecundity values by different authors and countries.
This great variability of the values found for the fecundity of M. cephalus can be because of the difference in length and age of the studied organisms, as there is a positive relation between fecundity and bigger and older aged females, even in the same area.
Relative fecundity values obtained in this study were 50,272 oocytes per gram, ranged from 9,425 to 150,011, higher values than those obtained by Ibáñez and Gallardo-Cabello for M. cephalus in the Tamiahua lagoon, Veracruz of 1,680 oocytes per gram and a range of 680 to 4,776 oocytes [40].
It was observed that in the same study area as M. cephalus, M. curema showed a fecundity of 9,612 to 238,795 in females of 0 to 5 years of age and lengths of 10.54 cm to 27.79 cm, and an average relative fecundity of 1,120 oocytes per gram (850 to 1,176 oocytes per gram), far below the values of M. cephalus.
• Sex ratio was 0.88:1 males: females.
• Average length of sexual maturity (L50) was 34.0 cm in both males and females with 4.64 years (males) and 4.98 years (females); average length of first maturity (L25) for both was 30.0 cm corresponding to 3.4 years of age.
• The gonadosomatic index was at its maximum values in November, December and January. A second very small period occurs during September and October. Mature organisms occur throughout the year.
• The allometric relationship between the liver weight and the fish length is negative (b=2.771). Monthly values of the hepatosomatic index are higher in June, July and August
• The gastric repletion index shows its highest values in June, February and October.
• The condition factor reaches its highest values in June, August and December with Clark and Safran EW indexes and July, September, October and November with Fulton and Safran TW indexes.
• Total fecundity was 1’422,076 to 1’747,736 oocytes for females of 3 to 12 years old.
• Relative fecundity was average: 2,830 oocytes?g-1 from 1,500 to 2,900 oocytes per female from 3 to 12 years old.
• Average oocytes diameter was 0.38 mm (from 0.22 to 0.52 mm).
The studies of the reproduction of the Mugil cephalus must be continued and published, because they are an important base line to compare if significant variations occur in the average size of sexual maturity (L50) and at first maturity (L25), which may indicate overexploitation of this resource.
Development of models of maximum sustainable yield and simulation capture should be taken into account to reach a rational management of this fishery, by capturing mature organisms that have already reproduced at least once and that will provide through recruitment new organisms to the population, preventing overexploitation of the resource. Also, ban seasons should be imposed and the law obeyed as in the mesh size of the fishing gear, to assure that only adults will be fished.
We want to express our gratefulness to fishermen, and also EG Cabral-Solís, D Brambila-López and A Pérez-Muñoz that provided samples to complete the information of this study.