Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2013) Volume 2, Issue 2
Keywords: Magnetite, Magnetic separation, Cyclodextrin glycosyltransferase, Bacillus lehensis, Immobilization
Cyclodextrins (CDs) are compounds formed from five or more units of glucose linked by α-1,4 glycosidic bonds. The most common CDs have six (α-CD), seven (β-CD) and eight (γ-CD) glucose units forming the cone shaped structure, that is able to accommodate hydrophobic molecules within is interior, the diameter of which is approximately 5.2 A° (α-CD), 6.6 A° (β-CD), and 8.4 A° (γ-CD) [1]. Externally, the structure is sufficiently hydrophilic to make CDs water soluble [2]. These cyclic glucose polymers are used in the pharmaceutical, cosmetic and food industries [3].
CD production involves starch catalysis by the enzyme cyclodextrin glycosyltransferase (CGTase), produced by bacteria of the genera Bacillus [4], Paenibacillus [5], Klebsiella [6] and Thermoanaerobacterium [7]. Enzyme stability during catalytic activity and the separation of the product from the reaction medium are important factors to address, when using CGTase for CD production. A viable alternative may be the use of immobilized CGTase. Immobilized enzymes preserve function during repeated use through an enhancement of functional stability [8]. Immobilization confines the physical enzyme to a defined region of space, allowing the catalytic activity to be used continuously [9]. Immobilized enzymes exhibit resistance to proteolysis [10], and can be stored at high temperatures [11]. Immobilized proteins retain their enzyme activity due to the non-modification of the structure of immobilized enzyme [10]. Moreover, immobilized enzymes have demonstrated the capacity for cyclic reuse [12], and are widely employed in different industries due to their desirable properties such as recovery, stability, selectivity and reduced inhibition by different substances [13].
Enzymes can be immobilized on an insoluble support for subsequent separation from the reaction medium. Magnetite (iron oxide) has been employed in biotechnology and biomedicine for the separation of biological molecules, as well as use in magnetic resonance, the controlled release of drugs and clinical diagnoses [14]. The encapsulation on porous support consists of physical entrapment of the enzyme in the internal cavities of a porous matrix. On porous substrates can be adhered substances that are not the enzyme of interest. Other limitation in the use of agarose, and of porous supports is the high cost. The method of covalent immobilization of the enzyme on the support present the advantage of stable nature of the bonds formed between enzyme and matrix, and the advantage of the enzyme not be released into the solution upon use.
It is important to characterize the physicochemical and kinetics properties of immobilized CGTase, which differ from those of the free enzyme. Magnetite offers an advantage over conventional supports due to the ease of recovery of the immobilized enzyme from the medium with the application of a magnetic field, thereby eliminating the need for expensive separation systems [15]. Nanoparticles can be used as supports for the immobilization of catalysts that are more accessible to reactants [16].
The main advantages of enzymes encapsulation in nanoparticles refer to the high surface-to-volume rate, which facilitates contact enzyme support. The method of hydrothermal synthesis of magnetic nanoparticles used in this work is cheap and easy realization, in comparison to methods of chemical synthesis.
The aim of the present study was to describe the immobilization of CGTase from B. lehensis on magnetite nanoparticles and determine the proprieties of the immobilized enzyme, which demonstrates greater stability in comparison to the soluble form.
Strain and enzyme
Bacillus lehensis was isolated from wastewater from a cassava flour mill in Brazil. The 16S rRNA sequence was deposited with the Genetic sequence database at the National Center for Biotechnology Information (NCBI) (GenBank: HQ399547) [4]. The strain was cultivated to produce CGTase in an optimized medium containing 6.96 g.L-1 of cassava starch, 8.07 g.L-1 of nitrogen sources, 9.45 g.L-1 of sodium carbonate, 1.00 g.L-1 of K2HPO4, 0.20 g.L-1 of MgSO4.7H2O and 10.00 g.L-1 of Na2CO3. Incubation was performed at 35 ± 1°C for 72 h on a rotary shaker (New Brunswick Scientific Co, NJ, USA), at 150 rpm with an initial pH of 9.2. The cell-free supernatant was used to obtain the partially purified enzyme through precipitation with ammonium sulfate at a concentration of 80%. The precipitate was re-suspended in HCl-Tris buffer, pH 8.0, and dialyzed using a 12 kDa cellulosic membrane against the same buffer. The buffer was changed every two hours.
Supports for immobilization
Hydrothermal synthesis: The support synthesis (magnetite nanoparticles) was performed using 10 M of FeCl3.H2O (97% pure) and 20 M β-CD. The reaction mixture was heated to 65°C for 30 min and the pH was adjusted to 10 by the addition of NH4OH 35% (w/v). Hydrothermal synthesis occurred in a steel reactor internally coated with Teflon, with total volume of 100 mL. The following were the reaction conditions: temperature of 150°C, pressure of 1907 atm and reaction time of 6 hours. The synthesized particles were washed six times with deionized water and dried at 100°C.
Support silanization and activationc
To silanize the support, the synthesized particles were treated with 3-aminopropyltriethoxysilane (APTES, Aldrich). The silanization process was as follows: 50 mg of synthesized particles were treated with 1 ml of APTES ( = 98%) at 80°C under 1500 rpm for 6 h, followed by a washing step with 0.05 M Tris-HCl, pH 8.0. To activate the support, the magnetite particles were re-suspended in 5% glutaraldehyde (w/v) in Tris-HCl buffer, pH 8.0, at room temperature under 1500 rpm for 4 h. The activated support was washed six times with deionized water at room temperature for 30 min. The particles were re-suspended in Tris- HCl buffer 0.05 M, pH 8.0, and stored at 4°C.
Immobilization
One ml of 0.05 M Tris-HCl, pH 8.0, containing 0.1 mg CGTase as the protein, was added to 100 mg of the synthesized magnetite support, as described in section 4.3. The reaction product was stirred in a thermo mixer at 200 rpm for 6 h. After chemicallythe coupling step, the supernatant was removed and the magnetite support was washed abundantly with deionized water.
Enzyme activity, partial purification and protein determination
Enzyme activity was measured based on the discoloration of phenolphthalein solutions, following the method described by Makela et al. [17]. The enzyme was incubated in 1% (w/v) potato starch in 50 mM Tris-HCl buffer, pH 8.0, at 55°C. Reaction samples were withdrawn at 0, 3, 6, 9 and 12 min, and the enzymes were inactivated at 100°C for 5 min. Total CD content was determined by absorption at 550 nm in a solution containing phenolphthalein (3 mM) and carbonate sodium buffer (0.6 M). One unit of CGTase activity was defined as the amount of enzyme that produces 1 µmol of β-CD.min-1. For partial purification, the enzyme was precipitated using 80% ammonium sulfate, resuspended in 0.05 M HCl-Tris buffer, pH 8.0, and dialyzed using a 12 kDa cellulosic membrane against 0.05 M HCl-Tris buffer, pH 8.0 for 10 h. Protein determination of CGTase was made using bovine serum albumin, as described in Lowry’s method [18].
Enzyme stability in reuse of immobilized CGTase
The CGTase immobilized on magnetite was incubated under the optimum conditions. Reuse was determined by measuring the absorbance (550 nm) of the phenolphthalein/β-CD complex, after its production in the enzyme reaction. At the end of each cycle, the immobilized CGTase was submitted to magnetic separation and washing with Tris-HCl 0.05 M, pH 8.0, followed by re-suspension in the same buffer to initiate a new cycle.
Characterization of immobilized CGTase on support
Optimum pH and temperature: Optimum pH was determined using 0.05 M buffers in the 3.0 to 10.0 range. The following buffers were used: glycine-HCL (pH 3.0), sodium acetate (pH 4.0 to 5.0), phosphate (pH 6.0), Tris-HCl (pH 7.0, 8.0 and 9.0) and glycine-NaOH (pH 10.0) at 55°C. The enzyme activity of immobilized CGTase was measured in accordance with that described in item 4.6. For the determination of optimum temperature, the purified enzyme was analyzed in the range from 35 to 85°C in Tris-HCl buffer, pH 8.
Thermal and pH stability of CGTase: The CGTase stability at pH 3.0 to 10.0 was determined by performing the reaction at 70°C (optimum temperature), with incubation for 12 h and 24 h. CGTase thermal stability assays were performed by incubating the enzyme at 70°C for 110 min.
Substances effect on immobilized CGTase: The effect of different compounds on the enzyme activity of immobilized CGTase was investigated using concentrations of 2 mM of each substance for 12 min at 70°C in 0.05 M Tris-HCl buffer, pH 8. In all experiments, CGTase activity without the addition of inhibitors (control) was considered to be 100%.
Kinetic parameters of CGTase: For the determination of kinetic parameters, 5 mL of immobilized enzyme were mixed with 5 mL of starch solution (3, 3.5, 4, 5, 6, 7, 8, 9 and 10 mM) in 0.05 M Tris-HCl buffer, pH 8.0, at 70°C for 30 min. The Michaelis-Menten constant (Km) and maximum velocity (Vmax) were defined by the Lineweaver-Burk plot.
Immobilization efficiency
In the present study, the enzyme CGTase from Bacillus lehensis was immobilized on magnetite. The advantages to use of magnetite as a magnetic carrier of enzymes are their simpler fabrication processes, and ability to be manipulated under a magnetic field. From such properties can perform easily the enzymatic separation for its use more of one time. The properties this inorganic support includes physical and microbial resistance when compared to organic carrier as natural polymers (agarose, albumin, etc) and synthetic polymers (polyacrylamide). The availability of magnetite present low cost when compared with natural polymers, as proteins that require purification processes of high cost.
The magnetite was silanized with 3-aminopropyltrimethoxysilane and activated with glutaraldehyde. The activation processes are generally generating electrophilic groups on the support. In the signaling of the magnetite support, the residue 3-Aminopropyltrimethoxysilane (APTES) is dislocated to the hydroxyl group of the magnetite support surface, producing an alkylamino derivative. Glutaraldehyde activates the reaction for the formation of the Schiff base, which generates an intermediate–the active aldehyde group. The activated aldehyde from the support binds with CGTase in the presence of Tris-HCl buffer, pH 8.0. This pH prevents the polymerization of the aldehyde near the surface of the support with a positive charge. Binding occurs between the aldehyde groups of the enzyme and support. The reaction sequence is shown in Figure 1.
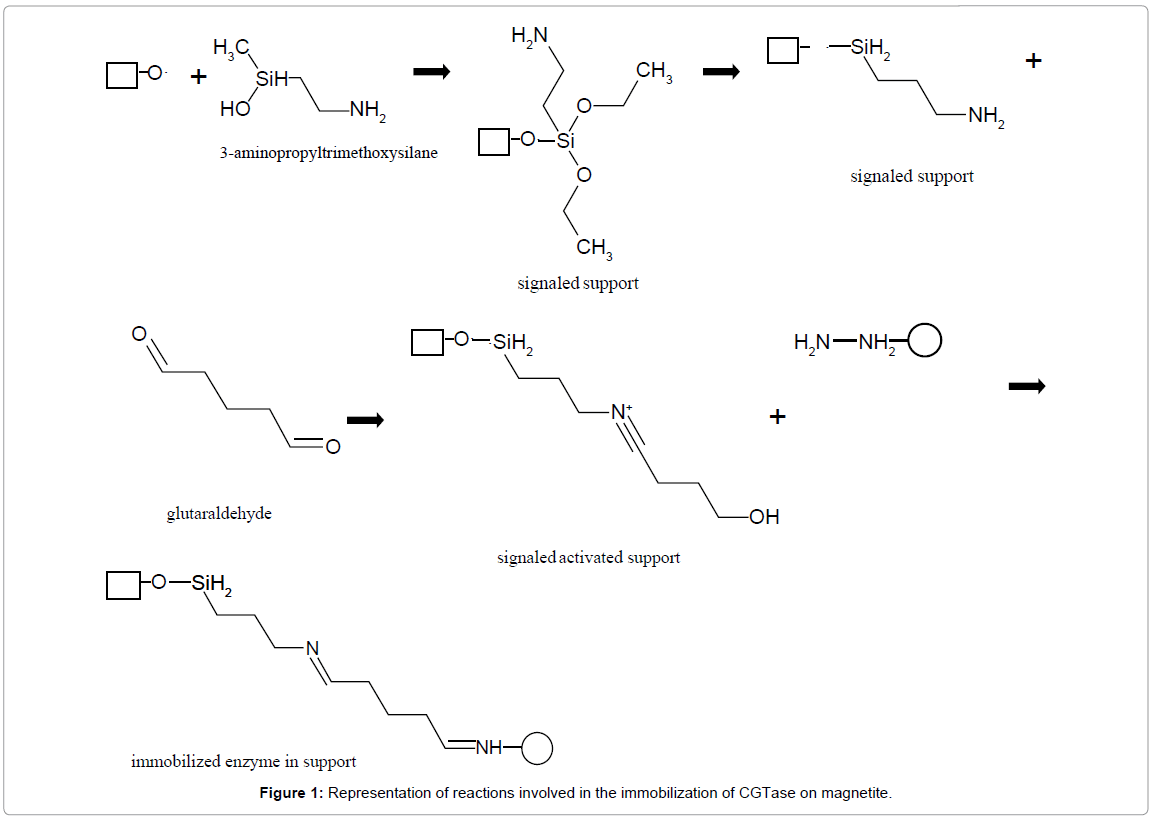
Figure 1: Representation of reactions involved in the immobilization of CGTase on magnetite.
The immobilization method using a magnetite support (silanized with 3-aminopropyltrimethoxysilane and activated with glutaraldehyde) was efficient for immobilizing CGTase from Bacillus lehensis. The covalent bond between the activated surface of magnetite and CGTase from B. lehensis was efficient for immobilization of the enzyme. The immobilization factor of 11.21 and 16.27% yield indicate that the enzyme was recovered 11 times in the immobilization process and 16% of the enzyme activity was maintained after immobilization on the magnetite support (Table 1). Thus, stable binding between the enzyme and matrix result on low yield of immobilized activity.
| Immobilization | Total activity (U) | Total protein (mg) | Specific activity (U.mg-1) | Yield (%) | Immobilization fold |
| Crude extract | 637.15 | 34.50 | 18.46 | 100.00 | 1.00 |
| Immobilized CGTase | 111.80 | 0.54 | 207.00 | 16.27 | 11.21 |
Table 1: Immobilization of CGTase from Bacillus lehensis CGII.
The specific activity of 207 U.mg-1 and immobilization fold of 11.21 demonstrates the high efficiency of this immobilization system for CGTase from B. lehensis. The specific activity of the lipase on porous polypropylene was 7.8-fold, due to greater instability as binding of the enzyme with the porous support this system [19].
CGTase immobilized on chitosan exhibited recovered enzyme activity of 5.2% [20]. Table 2 displays the results of the cycles of immobilized CGTase on magnetite. With each use of CGTase, enzyme activity was reduced. The covalent bond between the enzyme and the matrix decays the enzymatic activity, beyond this link (irreversivel) to occasion the discard of matrix together with the enzyme. However, CGTase function was maintained for four cycles. A similar finding is reported with CGTase immobilized on magnetic nanoparticles over seven cycles. However, Ivanova [21] using nanoparticles of magnetite as support for lipase immobilization obtained after 20 cycles [22]. According to Sheldona [23], the loss of activity in successive reuses reflects either the amount of linked CGTase on the carrier that is released into the assay solution or the loss enzyme function.
| N° of cycles | Enzyme activity (U.mL-1) | Retention of activity (%) |
| 1 | 112.00 | 100.00 |
| 2 | 105.65 | 94.33 |
| 3 | 53.91 | 51.03 |
| 4 | 10.44 | 19.37 |
Table 2: Cycles of immobilized CGTase on magnetite.
Optimal pH and temperature and thermal and pH stability
The analysis of the activity of an immobilized enzyme involves specific assays, as it exhibits a different structure from that of the soluble enzyme. Optimum temperature and pH for immobilized CGTase were evaluated in the range of 50 to 100°C and 3 to 10, respectively. Figure 2a and 2b displays the results of enzyme activity at an optimum temperature of 70°C and optimum pH of 8. A significant difference was found between soluble and immobilized CGTase regarding pH. However, the temperature of 70°C was 15°C higher than the optimum temperature for the soluble CGTase. This physical characteristic is important for applications in enzymatic industrial processes [22], due the enzymatic reaction generally be accelerated and reduce contamination. Conformational changes in the protein upon immobilization can cause changes in the properties of the immobilized enzyme such, as optimum temperature, pH, etc. Matte et al. [24] report a reduction in optimum temperature of immobilized CGTase from Thermoanaerobacter [23].
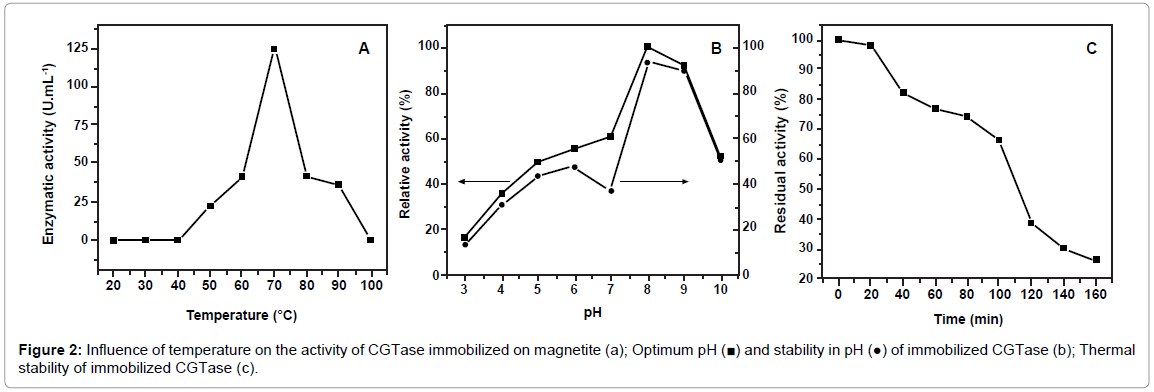
Figure 2: Influence of temperature on the activity of CGTase immobilized on magnetite (a); Optimum pH (■) and stability in pH (●) of immobilized CGTase (b); Thermal stability of immobilized CGTase (c).
The optimal temperature of enzyme free changed after of its immobilization. This indicates the possibility of a change in its structure. The conformational change of the immobilized enzyme increased its optimal temperature.
Thermostability is an important aspect in industrial applications of immobilized CGTase. Figure 2c displays the results of the thermal stability assays, demonstrating that 26.22% of the initial activity was retained after 160 min at the optimum temperature of 70°C. The thermal stability of immobilized enzymes is often not greater than that of soluble enzymes, as demonstrated by Matte et al. [24] for CGTase immobilized on chitosan.
Maximum activity of the immobilized CGTase from B. lehensis was achieved at pH 8.0 (Figure 2b), which was the same as that for soluble CGTase. The optimum pH of immobilized enzyme no changed of its free conformation. This indicate that the enzyme structure no was affected. Ivanova [21] found that the optimum pH for CGTase from Paenibacillus macerans immobilized on magnetic nanoparticles was 7, which was the same as that for the soluble enzyme. Bruno et al. [8] suggest that immobilization confers a kind of protection to enzymes, demonstrating different optimum pH for immobilized and soluble enzymes. Figure 2b shows that 50% of relative activity was maintained in the pH range of 3 to 10 after incubation for 24 h at 4°C. Stability in different pH values is due to the ability of the immobilized enzyme to exist in a stable conformation.
Effect of substances on immobilized CGTase
Enzyme stability was measured based on activity in the presence of 2 mM of different substances. The inhibitor/activator effects of the substances on immobilized CGTase are shown in Table 3. The ions, Mg2+, Mn2+ and Cu2+ and sodium dodecyl sulfate and ethylenediamine tetraacetic acid inhibited CGTase activity, indicating that the presence these ions interferes with the catalytic activity of CGTase on immobilized magnetite. These results are in agreement with findings reported for CGTase from the alkaliphilic Bacillus pseudalcaliphilus 20RF [25]. In contrast, Ca2+, Zn2+, Na and Fe did not exhibit an inhibitory potential on the enzyme, as the loss of enzyme activity in the presence of these substances was negligible (2.01%). The catalytic activity of enzymes depends on the integrity of its protein structure. With the structure of CGTase stabilized by immobilization, the catalytic center of the enzyme was not inhibited by these substances.
| Ions and reagents (2 mM) | Residual activity (%) |
| No reagent | 100.00 |
| CaCl2 | 99.80 |
| Fe(NH4)2(SO4)2.6H2O | 98.73 |
| FeCl3 | 100.34 |
| FeSO4.7H2O | 98.12 |
| Fe2(SO4)3.nH2O | 98.01 |
| Ca(CH3COO)2 | 99.72 |
| CaCO3 | 98.78 |
| ZnSO4.H2O | 98.59 |
| Na2SO4 | 99.04 |
| CuSO4.5H2O | 73.92 |
| MnSO4.H2O | 50.39 |
| Sodium dodecyl sulfate (SDS) | 74.48 |
| MgSO4.7H2O | 65.03 |
| Ethylenediamine tetraacetic acid (EDTA) | 64.24 |
Table 3: Effect of additives on immobilized CGTase from Bacillus lehensis.
Kinetic parameters
Enzymatic activity is defined with the CD formation per enzyme unit and time. The Michaelis-Menten model was used in this work to characterize a system of immobilized enzyme in magnetic support. The Michaelis-Menten model can be used to evaluate effects of immobilization on enzyme activity in the immobilized state. The Michaelis-Menten kinetics occurs when the rate of enzyme reaction for CD formation is constant, even with increased concentration of starch. The Vmax of product form of 45.45 mol.ml-1.min-1 was determined with of 0.826 mg.ml-1.
The present study developed magnetite nanoparticles to immobilize CGTase for use as a recyclable producer of CD. The enzyme was immobilized on a magnetite support, to assist in the separation of the enzyme from the reaction medium under the influence of a magnetic field. CGTase immobilization on magnetite altered enzyme properties, such as its optimum temperature, which was increased by 15°C, indicating that the support promoted the thermal protection of the enzyme. CGTase recovery by separation from the reaction mixture in the presence of a magnetic field proved efficient, allowing the reuse of the enzyme for four cycles.
The authors would like to express their gratitude to the National Council for Scientific and Technological Development (CNPq) for financial assistance in this research.