Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2015) Volume 4, Issue 2
The Asian scorpion Buthus martensii Karsch has been used in Chinese Traditional Medicine for thousands of years. The scorpion, venom of scorpion and their extracts are effective in treating a variety of nervous system diseases such as epilepsy, apoplexy pains and facial paralysis. Apart from these therapeutic effects, the peptides purified from scorpion also showed antitumor activities, antimicrobial effects and protective effects on vascular. In this review, we reported various pharmacological effects of bioactive peptides purified from scorpion Buthus martensii Karsch so that we can use different effects of these bioactive peptides to treat different diseases and provide reference for the study of scorpion Buthus martensii Karsch in future.
<Keywords: Buthus martensii Karsch; Bioactive peptides; Pharmacological activities
Scorpion Buthus martensii Karsch (BmK) is one of the most ancient animals widely distributed in Mongolia, Korea and China. In China, it has been one of the indispensable and essential materials traditionally used as Chinese medicine for treating the symptoms associated with convulsive spasm, cardiovascular and cerebrovascular diseases, inflammation, hepatopathy and tumours for 1000 years since the Sung Dynasty [1,2]. Over 400 million-years evolution, a great diversity of bioactive peptides has been conserved in the venom glands of scorpion for defense or prey [3].
To date, there have been an increasing number of researches identifying the effective components and pharmacological properties of scorpion BmK. Scorpions and their venoms have been extensively studied and more than a hundred peptides have been isolated and characterized successively. Some of these identified scorpion peptides have been demonstrated to have considerable therapeutic effects such as antimicrobial activity, antitumor, analgesic activity, anti-epilepsy, antivascular disease and so on (Table 1). Although the clinical researches about pharmacological effects of BmK is less, BmK as a alternative and complementary medicine has been widely used.
| Pharmacological property | Active peptides | References |
|---|---|---|
| Antimicrobial activity in vitro and in vivo | BmKn2, BmKb1 BmKn2-7 compounds 1,2 |
Zeng et al.[7] Cao et al.[8] Gao et al.[9] |
| Inhibiting the glioma cell migration in vitro | BmK CT | Fu et al.[13-16] |
| Antitumor against Ehrlich ascites and S-180 fibrosarcoma in vitro | BmK AGAP-SYPU2 | Shao et al.[17] |
| Antinociceptive activity in vivo | BmK IT2 | Wang et al.[22] |
| BmK dITAP3 | Guan et al.[23] | |
| AngP1 | Guan et al.[24] | |
| BmK AGP-SYPU1 | Wang et al.[25,26] Deng et al.[27] |
|
| BmK AGP-SYPU2 | Zhang et al. [28,29] Zhao et al.[30] |
|
| BmK AGAP | Liu et al.[31] Ma et al.[32] Mao et al.[33] Cui et al.[34] |
|
| BmKAS | Cui et al.[35] | |
| Anti-epilepsy activityin vivo | AEP | Zhou et al.[37] Wang et al.[38] |
| Anti-vascular disease in vivo | Martentoxin SVAP |
Wang et al.[39] Song et al.[42] |
Table 1: Main pharmacological properties of Buthus martensii Karsch.
Antimicrobial activity
Antimicrobial peptides (AMPs) are widely distributed in various species including plants, insects, humans, as well as single-celled organisms [4]. The structure of AMPs is usually amphiphilic topologies [5,6]. Recently, researchers have reported and identified some venom peptides from BmK, which are found to show antimicrobial activity in vivo and in vitro. Some of these peptides present highly amphiphilic structure. Xian et al. [7] identified two amphiphilic venom peptides BmKb1 and BmKn2 from Buthus martensii Karsch. The functional tests showed that BmKn2 exhibited strong inhibitory activities against both Gram-positive and Gram-negative bacteria while BmKb1 had weaker activity than BmKn2 in suppressing the growth of these bacteria [7]. Furthermore, BmKn2-7 (Figure 1), a derivative of BmKn2 designed by Cao et al. [8] showed higher antibacterial activity against clinical antibiotic-resistant strains and increased inhibitory activity against both Gram-positive and Gram-negative bacteria. Additionally, the hemolytic activity of BmKn2-7 was distinctly decreased compared with BmKn2. Animal studies showed that the topical use of BmK2n-7 protected the skin of mice infected by Staphylococcus aureus.
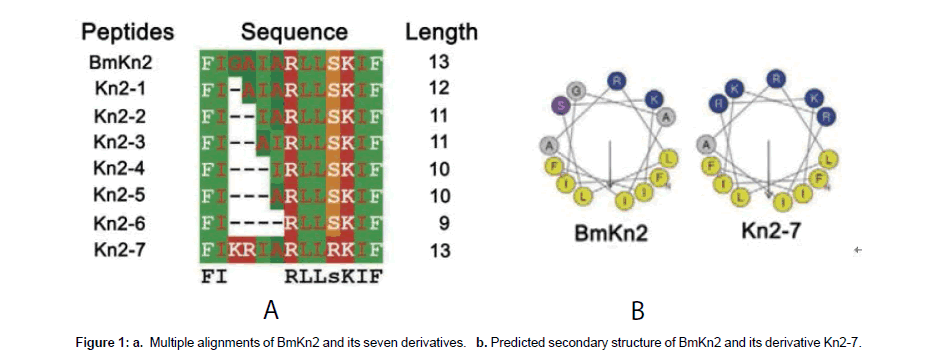
Figure 1: a. Multiple alignments of BmKn2 and its seven derivatives. b. Predicted secondary structure of BmKn2 and its derivative Kn2-7.
Gao et al. [9] isolated two compounds, 3β-acetoxyl,2,14,22- trihydroxy, 19-hydroxymethyl,9α,5β,14β-card-20(22)enoli (1; Figure 2) and 1,2,3,4-tetrahydro-6-hydroxy1-5-pyrimidinecarbox-aldehyde (2; Figure 2), from 70% extraction of Chinese scorpion BmK with medium pressure liquid chromatography and RP-HPLC analysis technology. Both compounds exerted inhibitory activities against the Gram-positive bacteria Bacillus subtilis.
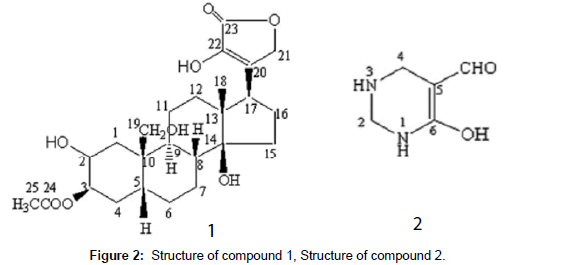
Figure 2: Structure of compound 1, Structure of compound 2.
Antitumor activity
Gliomas cell can diffuse into the normal brain tissue. And Matrix metalloproteinase-2(MMP-2) and chloride ion channel play an important role in invasion of glioma cell [10,11]. Recently, researchers found that Cltx, as a specific chloride channel blocker, can bind specifically to glioma cell surface and inhibit the glioma cell migration. The first chlorotoxin (Cltx)-like peptide, BmK CT, isolated from the venom gland of BmK [12], is a novel blocker of the chloride ion channel and MMP-2 [13]. Yuejun Fu et al. [14] studied the influence of basic residues in BmK CT on the inhibitory activity to MMP-2 and proposed a model of structural mechanism in inhibition of glioma cell migration by BmK CT. Subsequently, Yuejun Fu et al. [15] found pEGFP-N1 mediated BmK CT expression displayed a significant suppression in migration of rat glioma C6 cells by in vitro studies. Moreover, Yuejun Fu et al. [16] also performed a study to observe the combination of gene therapy pEGFP-N1-BmK CT with lithium chloride (LiCl) on C6 glioma cells. They reported that the combination therapy could inhibit the secretion and proliferation of pro-MMP2 in a synergistic manner, which might provide a new therapeutic strategy for glioma cancer. Besides, JianHua Shao et al. [17] isolated a dual-function peptide with analegestic and antitumor activities from scorpion venom. The peptide, BmK AGAP-SYPU2, showed antitumor effect in the Ehrlich ascites model and S-180 fibrosarcoma models in vivo.
Antinociceptive activity
Scorpion and its extracts can be used for soothing nerves and relieving pains caused by meningitis, cerebral palsy and rheumatism [18-21]. To date, more and more analgesic peptides from venom of the scorpion BmK have been found such as BmK IT2 [22], BmK dITAP3 [23], AngP1 [24], BmK AGP-SYPU1 [25-27], BmK AGP-SYPU2 [28-30], BmK AGAP [31-34] and BmKAS [35]. Moreover, Li et al. [19] reported that the venom of scorpion did not elicit dependence, while the commonly used analgesic substances such as morphine, heroin, and aspirin present the addictive nature. Therefore, the analgesic peptides from scorpion can be used as potential analgesic ingredients.
Depressant and excitatory insect neurotoxins: Scorpion neurotoxin can be divided into α- or β- mammal neurotoxins and depressant or excitatory insect-selective neurotoxins [20,21]. BmK IT2 and BmK dITAP3 are depressant neurotoxins, which had been previously purified from venom of the scorpion BmK. The bioassay showed that both peptides could produce antinociception in rats [22,23].
In addition, an excitatory insect neurotoxin, AngP1, purified from scorpion BmK was reported to have 43% inhibition efficiency for analgesia at a dose of 5 mg/kg on mice. And AngP1 was definitely devoid of mammalian toxicity [24].
BmK AGP-SYPU1: Yu Wang et al. [25] initially purified the analgesic peptides named BmK AGP-SYPU1 from the venom of Chinese scorpion BmK through a four-step chromatographic process which was composed of amino acid residues. The recombinant BmK AGP-SYPU1 was also purified by Yu Wang et al. with affinity column and cation exchange chromatography. The mouse twisting assay indicated that the native and recombinant BmK AGP-SYPU1 showed similar analgesic effects on mice. Subsequently, Yu Wang et al. [26] further investigated the relationship between the arginines in the C-terminal (65-67) of BmK AGP-SYPU1 and the analgesic activity. They found the arginines in the C-terminal were crucial for the analgesic activity and might be located at analgesic functional sites.
Recently, the study by Li Deng et al. [27] identified a new relationship between the structure and analgesic activity of the scorpion BmK AGP-SYPU1. They performed the study to investigate the role of wo conversed tyrosines (Tyr5 and Tyr42) from BmK AGP-SYPU1. The mouse-twisting test showed that Tyr5 and Tyr42 were involved in the analgesic activity. And the results of molecular simulation revealed that the core domain of BmK AGP-SYPU1 (Figure 3) was the key to analgesic activity and Tyr42 might be associated with pharmacological function when the core domain conformation was altered.
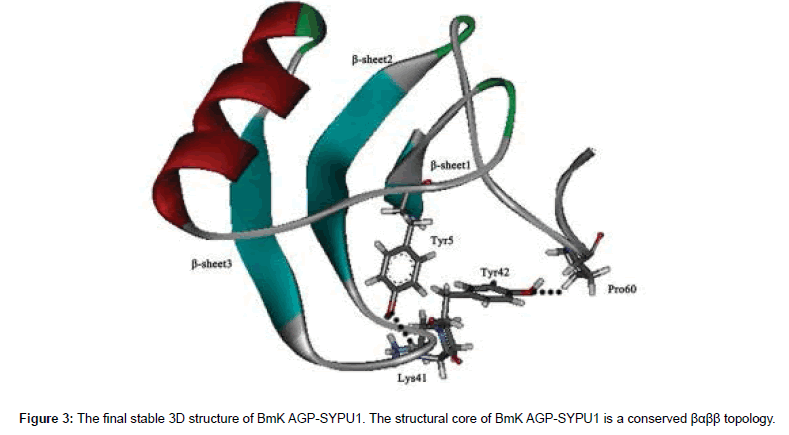
Figure 3: The final stable 3D structure of BmK AGP-SYPU1. The structural core of BmK AGP-SYPU1 is a conserved βαββ topology.
BmK AGP-SYPU2: Rong Zhang et al. [28] purified an analgestic peptides, BmK AGP-SYPU2, from the Chinese scorpion BmK through five continuous chromatographic steps. They identified the target peptides in every separation step by the mouse twisting model test and determined the molecular weight, isoelectric point and N-terminal sequence of the peptide. Animal studies demonstrated that BmK AGPSYPU2 exhibited obvious analgesic effects compared with morphine. Rong Zhang et al. [29] further investigated the functional role of glycines in the C-terminal end of BmK AGP-SYPU2. They detected the analgesic activity of nature BmK AGP-SYPU2 and its mutants which were obtained by site-directed mutagenesis. The study indicated that glycines at the C-terminal end played an important role for the analgesic activity of BmK AGP-SYPU2.
In addition, Yongshan Zhao et al. [30] performed three molecular dynamics simulations (BmK AGP-SYPU2 structure and its two mutants of its structure) to investigate the relationship between the C-terminal segment of BmK AGP-SYPU2 and its analgesic activity. The data of these calculations suggested that the glycine residues at the C-terminal segment stabilized the protruding topology of the NC domain, which was an important decisive factor for the analgesic activity of BmK AGPSYPU2.
AGAP: Antitumor–analgesic peptide (AGAP) from the venom of the Chinese scorpion BmK has both analgesic and antitumor activities. YanFeng Liu et al. [31] reported the expression and purification of AGAP in Escherichia coli and demonstrated the analgesic and antitumor activities of AGAP on mice. Rui Ma et al. [32] looked for the analgesic domains by using the conformational stability, catalytic activity and folding, and site-directed mutagenesis in disulfide bridges and examined the analgesic activity of 12 mutants through the mousetwisting assay. They found the key to the analgesic activity, which was called “core domain”. Qinghong Mao et al. [33] investigated the antinociceptive effect and underlying mechanism of AGAP from scorpion BmK. The study showed that formalin-induced inflammatory pain could be suppressed by pre-intraplantar injection of AGAP in a dosedependent manner through a MAPKs-mediated mechanism in mice. Furthermore, AGAP could potentiate the effects of the inhibitors of MAPKs on the inflammatory pain. In addition, Yong Cui et al. [34] studied the role of residues in AGAP by using an effective Escherichia coli expression system and evaluated the extent to which residues of AGAP contributed to its analgesic activity. And they pinpointed the important residue for the analgesic activity successfully.
Anti-epilepsy activity
In China, scorpions have been used to treat epilepsy and convulsion by Chinese traditional doctors since the Sung Dynasty (A.D. 960-1279) [36].
The first anti-epilepsy peptide (AEP) was isolated and purified from venom of scorpion BmK by Zhou et al. [37]. The peptide is composed of 66 residues from 18 amino acids and has molecular weight of 8290. The pharmacological test suggested that AEP could suppress epilepsy induced by coriaria lactone and cephaloridine and had no side effect on heart rate and blood pressure in rats. Chun-Guang Wang et al. [38] further studied BmK AEP and confirmed its anti-epilepsy effect on rat. The cDNA sequence of BmK AEP showed that it shared homology with other depressant insect toxins, but there was a remarkable difference between them mainly focused at residues 6, 7 and 39, which might relate to the unique action of BmK AEP.
Anti-vascular disease
In certain vascular disease, nitric oxide (NO) can be released from the endothelium, which is one of the most important vasoactive substances [39]. NO is produced by nitric oxide synthase (NOS) which includes neuronal NOS (nNOS), endothelial NOS (eNOS) and inducible NOS (iNOS) in mammals [40,41].
NO release in endothelium is associated with Hyperpolarization which caused by the activation of Ca2+-activated K+ channels (BKCa channels) Jun Wang et al. [39] purified a large-conductance Ca2+-activated K+ channel inhibitor, martentoxin, from BmK and demonstrated its protective effects in inflammatory vascular diseases. They found martentoxin decreased NO production and iNOS activation induced by TNF-α. They also demonstrated that martentoxin retarded the down-regulation of eNOS mRNA induced by TNF-α in human umbilical vein endothelial cells (HUVECs).
Yimin Song et al. [42] performed studies to investigate effects of scorpion venom active polypeptide (SVAP) from scorpion BmK on platelet aggregation in ex vivo and vitro in rabbits, thrombosis in carotid artery of rats as well as plasma 6-keto-PG F1α and TXB2 in rats. The results showed that SVAP significantly inhibited the rabbit platelet aggregation triggered by thrombin and ADP in vitro and prolonged the occlusion time of thrombosis induced by electrical stimulation. Additionally, SVAP could distinctly increase the plasma concentration of 6-keto-PG F1α and value of PG I2/TXA22, but slightly effect rat’s plasma concentration of TXB2 in vitro and in ex vivo, which indicated the mechanism of the antithrombotic action of SVAP.
Other beneficial effects
There are not only these pharmacological effects of BmK described sabove, but including other beneficial effects. As is well known, scorpion and its venom have been used in Traditional Chinese Medicine to treat chronic neurological disorders. However, the mechanism about that remains unknown. Tao Wang et al. [43] purified scorpion venom heatresistant peptide (SVHRP) from scorpion BmK and applied it in vitro and in vivo to study its effects on the neurogenesis and maturation of adult immature neurons and explored associated molecular mechanisms. The study by Tao Wang et al. [43] also suggested the novel therapeutic values of SVHRP. Furthermore, one study suggested that BmKbpp, a 47 amino acid peptide, displayed bradykinin potentiating and immunemodulatory activities [44]. The peptides SVAP purified from scorpion BmK have effects of inhibiting the rabbit platelet aggregation and thrombosis described above. Besides, one study indicated that SVAP could improve mesenteric microcirculation [45].
Scorpion Buthus martensii Karsch are rich in peptides which represent a tremendous unexplored resource for use in drug design and development. For now, more and more researchers study the role of amino acids in pharmacological activities and characterize the structurefunction relationships of bioactive peptides in order to design the peptides with higher bioactivities [8]. Based on the identified peptides purified from scorpion Buthus martensii Karsch, some researchers have tried to establish a new, economical and effective system for industrial peptides production with the use of molecular cloning technique [46]. However, some bioactivity peptides have not been identified and the mechanism of some pharmacological effects is not clarified. Therefore, the study about peptides should be further developed. Furthermore, though the scorpion has been traditionally used as a source of drugs, the clinical studies about single peptide are few. The careful clinical studies data still need to determine whether the peptides of scorpion Buthus martensii Karsch provide real clinical benefits.