Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Review Article - (2014) Volume 2, Issue 1
Salinity is becoming a major threat to plant productivity loss in agricultural system. Plants respond to saline
environment by modulating the inherent mechanisms to adjust to the changing environment. The understanding
of the mechanisms that plants operate under saline environment is essential beginning in efforts to reduce the
adverse effects of salinity stress. The agricultural system is tightly linked with the fertilizer input and thus the
judicious application of fertilizers is expected to lead positive effects in reversing the salinity effects. Sulfur is a
macronutrient with essential roles in plant development under optimal and stressful environment. Several compounds
are synthesized from sulfur metabolism useful in reversing the adverse effects of abiotic stress because of their
free radicals scavenging property. Sulfur-containing metabolites, amino acids (cysteine and methionine), vitamins
(biotin and thiamine), thioredoxin system, glutathione lipoic acid and glucosinolats have potential to promote or
modify physiological and molecular processes under salinity stress in plants. Thus, modulation of sulfur metabolites
production could alter physiological and molecular mechanisms to provide tolerance against salinity. The present
review discusses the role of sulfur-containing compounds in modifying various physiological and molecular processes
in plants to confer salinity tolerance in plants.
Salinity is a major abiotic stress which adversely affects plant processes at physiological, biochemical and molecular level and reduces plant productivity [1,2]. The fertility status of soil is deteriorating due to increasing salinity levels and has been a subject of concern to agricultural scientists for sustainable developments of crops [3]. It is expected to result in the loss of up to 50% fertile land by the middle of the 21st century [4]. The loss in plant productivity due to salinity has been associated to imbalance in ionic and nutrients status of plants [5,6], overproduction of Reactive Oxygen Species (ROS) and redox state of cell. The consequence of ROS overproduction in chloroplast, mitochondria and peroxisomes is damage to lipids, proteins, nucleic acids and integrity of cell membrane properties [7,8]. Among various mechanisms adopted by plants to cope with the increasing salinityinduced oxidative stress, maintaining redox state of cell through increased sulfur (S) metabolism and production of S-containing compounds are of paramount importance. It is, therefore, necessary to consolidate our understanding on the S metabolism for improving salinity tolerance in plants. As S is an essential element for all living organisms and occupies fourth place in importance after nitrogen (N), phosphorus (P) and potassium (K) in agricultural system, it could emerge as an element necessarily required for sustainability of crop plants under abiotic stress. Khan et al. [9] and Nocito [10] have shown that S is necessary for abiotic stress tolerance of plants being an integral part of major metabolic compounds, such as amino acids (methionine; Met and cysteine; Cys), antioxidant (GSH), proteins, and sulfolipids. In addition, S is also a component of iron-S- clusters, polysaccharides and lipids and a broad variety of biomolecules including vitamins (biotin and thiamine), cofactors (CoA and S-adenosyl-Met), peptides (GSH and phytochelatins) and secondary products (allyl-cysteine sulphoxides and glucosinolates).
It is expected that modulation of S metabolism in plants would help in alleviating adverse effects of salinity as its metabolites control wide range of plant processes. The production of S-containing compounds through S metabolism is linked to antioxidant system in plants under salinity stress (Figure 1). Studies have revealed various aspects of salinity tolerance mechanisms [5,11-13], but a consolidated approach on salinity tolerance through S metabolism and S-containing compounds is not available. The present review focuses on improving our understanding towards the potentiality of S metabolites in modulation of biochemical, physiological and molecular processes to confer tolerance in plants under salt stress.
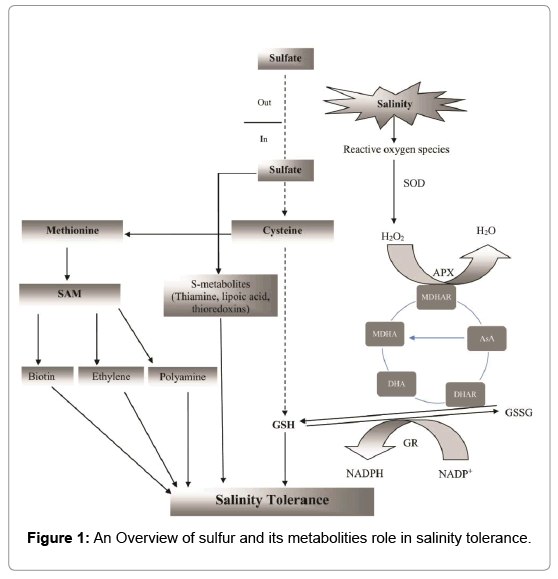
Figure 1: An Overview of sulfur and its metabolities role in salinity tolerance.
Salinity tolerance is a complex phenomenon as salinity has its multigenic and quantitative nature that may occur at multiple stages of plant and affects plant development. Salinity induces ionic and osmotic effects by increasing water deficit. This causes generation of ROS and oxidative stress. The increase in salinity levels increases ion toxicity primarily due to the accumulation of Na+ and Cl-, restriction of mineral nutrients uptake, loss of osmotic potential and ROS production and alterations in various physiological processes [9,14,15]. The increased ROS production causes damage to cells, but in addition, they induce signals for enhancing some protective mechanisms to scavenge ROS as well as damaging agents contributing to stress injury in plants [16]. The scavenging of ROS in chloroplast has been shown to be essential for tolerance to salinity in salinity tolerant cultivars [17].
Regulation of ion transport system is primary step to plant salt tolerance. Salt Overly Sensitive (SOS) pathway has a regulatory role in ion homeostasis under salinity stress. In plants, SOS signaltransduction pathway has been extensively studied and is considered as important for the maintenance of ion homeostasis and salt tolerance [18]. It is believed that ROS activate mitogen-activated protein kinase (MAPK) signaling pathways in plants under salinity stress, and MAPK kinase signaling cascades activated by ROS help in mediating salinity tolerance [19]. In addition to salinity stress, MAPK signaling modules are involved in eliciting responses to various stresses in which H2O2 activates several MAPKs [20]. Mishra et al. [21] demonstrated the role of several plant MAP kinases in response to salinity. In Arabidopsis, H2O2 activates the MAPKs, MPK3, and MPK6 via MAPKKK ANP1, and the over expression of ANP1 in transgenic plants resulted in increased tolerance to heat shock, freezing, and salinity [22]. A new MAPK gene (CbMAPK3) from Chorispora bungeana plant and its transcript level was found up-regulated in response to stress [23].
Antioxidant system has been extensively studied in plants in relation to its significance in salt tolerance. The activity of antioxidant enzymes has been found associated with both salt tolerant and salt sensitive types of plants, but differences lie with the ability of plant types to detoxify ROS [24]. Both enzymatic and non-enzymatic antioxidant defense systems guard plants against oxidative stress, and the increased activity/content of antioxidants has been found related to the improved salt tolerance [6,25]. The increased activity of antioxidant enzymes, superoxide dismutase (SOD), ascorbate peroxidase (APX) and glutathione reductase (GR) has been shown under salinity stress [5]. Similarly, increased content of non-enzymatic antioxidants, proline [13] and glutathione (GSH) [14] and their association with salinity tolerance have been reported. The relationship between activation of antioxidant enzymes and salinity tolerance has been shown in a wide range of plants, including Triticum aestivum [13,14], Brassica juncea [26], Vigna radiata [5], Vicia faba [27], Plantago [28], Lycopersicum, Zea mays [29], Pisum sativum [30], Arabidopsis and Oryza sativa [31].
Sulfur Metabolism: Regulation and Role in Salinity Tolerance
Most of the S in higher plants is taken up in the form of sulfate through sulfate transporter from soil. Sulfate can also be taken up through leaves in the form of S gases like H2S or SO2. Sulfur assimilation pathway and synthesis of reduced S compounds are well documented. Once S is reduced in the cell, it either remains in cytosol or transported into the plastid or stored in the vacuole for further metabolic reactions. The assimilation of S is highly regulated in a demand-driven manner [32]. First step in S assimilation involves the activation of sulfate to adenosine 5’-phosphosulfate (APS) catalyzed by ATP sulfurylase (ATP-S; EC 2.7.7.4). A relatively minor extension of this pathway is the phosphorylation of APS to 3’-phosphoadenosine-5’-phosphosulfate (PAPS), catalyzed by APS kinase (APK; EC: 2.7.1.25). Subsequently, APS is reduced to sulfite by APS reductase (APR; EC: 1.8.99.2) and the sulfite is further reduced to sulfide by a ferredoxin-dependent sulfite reductase (SiR; EC: 1.8.7.1) and the sulfide is incorporated into the amino acid skeleton of O-acetyl-L-Serine (OAS) by O-acetyl-L-Serine (thiol) lyase (OAS-TL), forming Cys [33]. Serine acetyl transferase (SAT; E.C 2.3.1.30), catalyzes the formation of OAS from L-Serine and acetyl-CoA links the serine metabolism to Cys biosynthesis [34]. Subsequently, Cys is formed by the condensation of sulfide and OAS, catalyzed by cysteine synthase (CSase). Synthesis of Cys and the activity of SAT and OAS-TL are strongly regulated by protein-protein interactions in the multi-enzyme complex of CSase [35,36]. Activity of SAT is strongly activated by association with OAS-TL, which is inactive and has only a regulatory role in the complex. The bound SAT exhibited higher apparent affinity for its substrate, acetyl-CoA and L-serine which might offer the possibility of regulation of synthesis of Cys in plants [37]. Consequently, the formation of Cys is the crucial step for assimilation of reduced S into S-containing organic compounds.
Plants have potential to develop several mechanisms to overcome the adverse effects of salinity stress. Among all strategies adopted by plants to cope salinity stress, availability of S along with S-containing compounds is of paramount importance. ATP-S is considered as the first rate limiting enzyme of the S assimilation pathway and is up-regulated under salinity stress [38]. It has been shown that salinity regulates key enzyme of sulfate assimilation, APR activity; mRNA levels of three APR isoforms increased by 3-folds with 150 mM NaCl treatment. Moreover, the increase in APR activity can be correlated with a higher rate of Cys synthesis to adjust the increased demand for GSH [39]. The activity of other enzymes of sulfate assimilation, SAT and a cytosolic isoform of OAS-TL are also induced by salinity [38]. It has been reported that salinity induces transcription and translation of OAS-TL genes probably due to higher demand of Cys or other S-containing compounds required by the plant as an adaptation/protection against higher level of salinity [40]. Most abundant cytosolic OAS-TL isoform, OAS-A1, is known to be involved in the defense responses of Arabidopsis against salinity [41,42]. The activity of OAS-TL was found increased under salinity in Typha and Phragmites contributing substantially to satisfy the higher demand of Cys for adaptation and protection. Higher Cys synthesis in both the plants supports the efficiency of the thiol-metabolism based tolerance [43]. The over-expression of CSase, or SAT, key enzymes of Cys biosynthesis was related to higher tolerance to oxidative stress and the overexpression of these enzymes is considered promising tool for engineering S assimilation for higher Cys and GSH synthesis [44]. Gene expression of Atcys-3A, coding for cytosolic OAS-TL, was induced as a tolerance mechanism under salinity [41]. In salt-treated plants, sufficient S supply allows GSH synthesis necessary to prevent the adverse effects of ROS on photosynthesis. Plants with higher levels of thiol compounds in S-sufficient plants were more able to remove the toxic effects of salinity and were more salinity tolerant [45]. Overexpression of BrECS1 and BrECS2 in transgenic rice plants tolerated high salinity by maintaining a cellular redox state, which prevented unnecessary membrane oxidation. These rice plants also showed lower relative ion leakage and higher chlorophyll-fluorescence than wildtype rice plants on exposure to salt, resulting in enhanced tolerance to abiotic stresses [46].
S is a ubiquitous and essential element for all living organisms from bacteria to animals and plants. S played many important roles in plants under optimal and stressful environments, but cannot involve directly in any specific metabolic role in plants. The presence of S in biomolecules is responsible for their catalytic or electro-chemical properties and thus for their involvement in specific biochemical mechanisms. In plants, S is an essential and integral part of amino acids i.e., Cys and Met, vitamins (biotin and thiamine), antioxidant (GSH), fatty acid (lipoic acid) and thioredoxin systems (Table 1). These biomolecules have structural or redox control in proteins, especially S-donation in iron-S cluster and vitamin biosynthesis and detoxification of ROS and xenobiotics [33,36,47-49]. In addition to these roles, S application improved photosynthetic efficiency and growth in two cultivars of mustard under salinity [50]. Cys synthesis in plants represents the final step of assimilatory sulfate reduction and the almost exclusive entry reaction of reduced S into metabolism of plants in a demanddriven manner [51,52]. In the following section, the importance of S metabolites is discussed individually.
| Sulfur-compounds | Plants | Response | NaCl concentration | Reference |
|---|---|---|---|---|
| Cysteine | Typha latifolia, Phragmites australis | + | 100 mM | [43] |
| Cysteine | Arabidopsis thaliana | + | 0.17 M | [40] |
| Cysteine | Brassica napus | + | 75mM 100mM | [38] |
| GSH | Hordeum vulgare | + | 100 mM | [45] |
| GSH | Catharanthus roseus | - | 80 mM | [93] |
| GSH | Triticum aestivum | + | 100 mM | [14] |
| GSH | Vigna radiata | + | 50mM | [5] |
| GSH | Lycopersicon esculentum | + | 100 mM | [93] |
| GSH | Brassica napus | + | 75 mM 150 mM | [38] |
| SAM | Lycopersicon esculentum | + | 5 g L-1 | [63] |
| Thiamine | Arabidopsis thaliana | + | 200 mM | [80] |
| Thiamine | Arabidopsis thaliana | + | 50 mM | [77] |
| Thiamine diphosphate | Zea mays | + | 0.1M, 0.2M,0.4M | [79] |
| Thiamine | Triticum aestivum | + | 100 mM | [81] |
| Thioredoxin (PsTrxo1) | Pisum sativum | + | 150 mM | [85] |
| Lipoic acid (DHLA) | Hordeum vulgare | - | 80 mM,160 mM,240 mM | [97] |
| Lipoic acid (DHLA) | Ocimum basilicum | + | 50 mM | [98] |
| Glucosinolates | Brassica oleracea | - | 40 mM 100 mM | [108] |
| Glucosinolates | Brassica oleracea | + | 40 mM 80 mM | [105] |
| Glucosinolates | Thellungiella salsuginea | + | 200 mM 300 mM | [106] |
| Glucosinolates | Thellungiella salsuginea | - | 400 mM 500 mM | [106] |
Table 1: Response of sulfur metabolites in salinity tolerance.
Cysteine is the metabolic precursor of essential biomolecules such as vitamins, cofactors, antioxidants and many defense compounds. It is synthesized in plants in the cytosol, plastids and mitochondria by the sequential action of enzymes in S assimilation. SAT synthesizes the intermediary product OAS, and OAS-TL combines a sulfide with an OAS to produce Cys. Most of the Cys is formed and accumulated in the cytosol [53] by the action of the major cytosolic OAS-TL, encoded by OAS-A1 [54,55]. It has been demonstrated that OAS-A1 is involved in the defense responses against abiotic stresses [41,42,56] and is essential for maintaining the antioxidant capacity of the cytosol [55,57]. In knock-out oas-a1 plants, intracellular Cys and GSH levels are significantly reduced, and the GSH redox state is shifted towards its oxidized form. Moreover, oas-a1 mutant accumulates ROS in the absence of external stress, and show spontaneous cell death lesions in the leaves. Besides, Cys in conserved form has paramount importance in the function and signaling of enzymatic processes under environmental stress [58]. Studies have shown that salinity induced higher rates of Cys synthesis with the increased expression of the cytosolic form of OASTL, and OAS-TL was related to salt tolerance [40,41,43]. Ruiz and Blumwald [38] showed that the increase in NaCl in the growth medium led to an increase in SAT in wild-type and transgenic canola plants. The increase in SAT was limited to only 10% in transgenic plants growing in the presence of 150 mM NaCl, whereas wild type plants showed a marked increase in SAT activity of about 2.5 times. The salinity induced differences in SAT activity between wild-type and transgenic plants were correlated with the leaf Cys concentrations because Cys content was found 1.5 times more in wild-type plants than transgenic plants. Cys can be converted into Met and further to S-adenosylmethionine (SAM) through reaction with ATP by SAM synthetase. In plants with S deficiency, the decline in chlorophyll together with decrease in SAM by many folds has been observed [59]. S-containing amino acid Met is an essential amino acid and executes central role in the initiation of mRNA translation of plants. Met is also a fundamental metabolite in plant cells as it directly or indirectly regulates a variety of cellular processes such as the precursor of SAM, which is the primary biological methyl group donor in cell wall synthesis, secondary metabolites, chlorophyll synthesis, DNA replication [60], ethylene, glycine betaine [14] nicotianamine and polyamine [61]. In addition, SAM is required for other methylation reaction of chlorophyll biosynthesis [59]. In addition, Met has nutritional importance in crop plants [62]. Sanchez-Aguayo et al. [63] have established a relationship between the increased levels of SAM synthetase (enzyme in SAM synthesis from Met) content and salinity tolerance in tomato plants. Recently, Ogawa and Mitsuya [64] showed that utilization of S-methyl methionine was involved in salinity tolerance at the germination and early growth stages of Arabidopsis. Thus, plant development under stress conditions could be augmented with enhanced Met metabolism. The derivatives of methionine, ethylene and polyamines (PAs) have also been shown to induce salinity tolerance in plants. Cao et al. [65] have given an insight into the link between ethylene signaling pathway and salinity tolerance. They also showed that ethylene signaling modulates salt response at different levels, including membrane receptors, components in cytoplasm, and nuclear transcription factors in the pathway. Recently, Abbas and Morris [66] showed that MAPK signal transduction pathways are important to salt tolerance. The over expressed transgenic barley lines had constitutively higher levels of ethylene with jasmonic acid and showed tolerance to salinity; and after two weeks of salt treatment barley transgenic plants showed less reduction in growth. Polyamines are Met derivative plant growth regulators which include putrescine, cadaverine, spermidine and spermine. The tolerance of plants induced by PAs has been correlated with elevated levels of putrescine, spermidine or spermine [67]. In a study, Yamaguchi et al. [68] have found that absence of spermine causes an imbalance in calcium homeostasis in the mutant plant in high salinity responses. In several other studies, putrescine, spermidine and spermine have shown potential in enhancing salt tolerance in plants [69-72].
Word “vitamin” generally links with the necessary dietary factors for animals. However, in general, plants are the only sources of vitamins. S is a core substance in nutritional vitamins i.e., biotin or vitamin H and thiamine or vitamin B1.
Biotin is a water-soluble vitamin biosynthesized by plants and is required by all living organisms for normal cellular functions and growth [73]. In plants, biotin plays a role as coenzyme which binds covalently to lysine residue of a group of enzymes and catalyzes many of reactions including carboxylation and decarboxylation [74]. Besides acting as a catalytic cofactor, biotin has a critical role in the enzymological mechanism of a number of enzymes that are essential in both catabolic and anabolic metabolic processes. Che et al. [73] demonstrated that biotin has additional non-catalytic functions in regulating gene expression in Arabidopsis plants. Biotin controls expression of the biotin-containing enzyme, methyl crotonyl-coenzyme A (CoA) carboxylase by modulating the transcriptional, translational and/or post-translational regulation of this enzyme. Similarly, Li et al. [75] suggested that biotin deficiency resulted in spontaneous cell death and modulated defense gene expression. However, role of biotin under salinity is not well studied till date, but Hamdia [76] have studied the influence of biotin in ameliorating the effects of salinity on growth and metabolism in lupine plants. More research is needed to evaluate the role of biotin in salinity tolerance.
Due to the abundance of thiamine in whole grains and green vegetables, plants represent the primary dietary source of thiamine in human and animal diets. Thiamine in the form of thiamine pyrophosphate (TPP), acts as a cofactor for several enzymes in key cellular metabolic pathways such as glycolysis, the pentose phosphate pathway and the tricarboxylic acid cycle (TCA); and in amino acid and isoprenoid biosynthesis [77]. Another form of thiamine is thiamine diphosphate (TDP), a coenzyme in a number of metabolic reactions including acetyl-CoA synthesis, TCA cycle, anaerobic fermentation, oxidative pentose phosphate pathway, the Calvin cycle and plant pigment biosynthesis [78,79]. Rapala-Kozik et al. [79] observed responses of Zea mays seedlings to abiotic stress including salinity and suggested the involvement of TDP-dependent enzymes metabolism under stress conditions. Total thiamine content in maize seedling leaves increased under salinity stress and the increase was found associated with changes in the relative distribution of free thiamine, thiamine monophosphate (TMP) and TDP suggesting a role of thiamine metabolism in the plants’ response to salinity. In another study, Rapala-Kozik et al. [80] has shown the involvement of biosynthesis of thiamine compounds and thiamine diphosphate-dependent enzymes in salinity and osmotic stress sensing and adaptation processes in Arabidopsis thaliana. El-Shintinawy and El-Shourbagy [81] observed that the addition of thiamine in 100 mM NaCl concentration alleviated the reduction of growth in plants and further they described that alleviation was correlated to the induction of 20 kDa and 24 kDa low molecular proteins in total protein content and increased contents of Cys and Met.
Thioredoxins are small (12-13 KDa), ubiquitous and heat stable proteins found in all types of organisms. Thioredoxins are believed to regulate the cell redox with active disulfide bridge [82] with a conserved pair of vicinal Cys (-Trp-Cys-Gly-Pro-Cys-Lys-) [83]. There are two distinct families of thioredoxins based on sequence of amino acids. Family I: one distinct thioredoxin domain whereas family II: fusion of proteins with one or more thioredoxin domains with additional domains [84]. Furthermore, family I is classified into six major groups: the thioredoxins f, h, m, o, x and y in higher plants. Thioredoxins f, m, x and y belong to chloroplasts, whereas the thioredoxin o is found in plant mitochondria. Marti et al. [85] showed a role of mitochondrial thioredoxin PsTrxo1 as a component of the defense system induced by 150 mM salt in pea mitochondria. The increase in mitochondrial Trx activity was observed in response to NaCl treatment and was correlated to mechanism by which plants respond to salinity and protects mitochondria from oxidative stress together with antioxidant enzyme. Similarly, Fernandez Trijueque [86] by using pea seedlings mRNA expression profile of the plastic PsTRX m1showed that TRX is expressed in cotyledons. Furthermore, the response of plastid TRXs to NaCl and its potential in restoring growth of TRX-deficient yeast under saline conditions was studied. It was suggested that there was reserve mobilization in seedling cotyledons and physiological functions of PsTRX m1 in the salinity response during germination. Zhang et al. [87] have reported that OsTRXh1 regulates the redox state of the apoplast and influences plant development and salinity responses. In addition, Serrato et al. [88] reported a novel NADPH thioredoxin reductase (NTR) in the chloroplast. Deficiency of NTR caused hypersensitivity to salinity in Arabidopsis thaliana as Arabidopsis NTRC knock-out mutant showed growth inhibition and hypersensitivity to salinity and suggested the role of NTRC gene in plant protection against oxidative stress.
Studies have shown the up-regulation of Cys synthesis in plants in response to salinity, suggesting a definite possible role of thiol in salt stress tolerance. Salinity creates over production of ROS and causes adverse effects on plant growth and metabolism which may be associated with the disturbances in osmotic potential of soil leading to osmotic stress or specific ion toxicity [24]. In order to cope the salt-induced adverse effects, plants develop defense mechanisms that include the up-regulation of synthesis of GSH which has essential roles within the plant metabolism in reducing the adverse effects of salinity stress [6]. GSH is a low-molecular weight S metabolite (thiol), nonenzymic antioxidant found in most of the cells. Synthesis of GSH is well regulated by S in a demand driven manner. Sulfate withdrawal from the growing medium decreases the levels of sulfate, Cys and GSH in plants leading to the up-regulation of sulfate transport systems and key enzymes of S assimilatory pathway [32]. It has been observed that increases in GSH synthesis are associated with up-regulation of the Cys synthesis. Similarly, S assimilation pathway enzymes have been found to be involved in the regulation of GSH synthesis. Queval et al. [89] reported that GSH accumulation was triggered by stress which resulted in the up-regulation of APR and SAT. Regulation of the GSH synthesis is dependent on the cell compartmentation as mitochondrial SAT is able to make major contribution to Cys synthesis under optimal conditions [90,91], whereas, the SAT in chloroplast was strongly induced during H2O2 triggered accumulation of GSH [89]. Oxidation state of GSH homeostasis (GSH/GSSG) can maintain cellular redox of GSH in plant cells [92]. The capacity of GSH to participate in the redox regulation in plant cells is, to a great extent, dependent on its absolute concentration and the ratio of GSH/GSSG under salinity stress [5,14]. The cellular GSH redox buffer present in cells forms major basis of redox homeostasis by which thiols proteins can maintain their redox state and can be reverted to their reduced form. Moreover, GSH is not only a redox buffer for cell, it also acts as an electron donor for scavenging of ROS in the major metabolic pathways of plants like photosynthesis, respiration or sulfate assimilation. Sensitivity of plants to salt plays a major role in the GSHmediated tolerance. Recently, Nazar et al. [5] have shown that Vigna radiata cultivars differing in salt tolerance have different rate of GSH biosynthesis; the GSH content was higher in salt tolerant than salt sensitive cultivar. The salt sensitive cultivar showed greater oxidative damage than salt tolerant cultivar. A study on Lycopersicon esculentum has also shown that salt sensitive plants contained lesser GSH content and redox state than the salt tolerant plants [93]. Similar reports are also available in Oryza sativa [94] and Arachis hypogaea [95]. GSH biosynthesis and its turnover differed in salt tolerant Lycopersicon esculentum and its wild relative Lycopersicon pennellii [96], and the salttolerant plants were able to maintain favorable GSH/GSSG redox state within 15 days in response to salt stress. Wild type Brassica napus plants accumulated higher GSH upon salt stress suggesting its protective role against salt-induced oxidative damage, whereas transgenic plants did not show these antioxidative responses [38]. In Nicotiana tobacum transgenics overexpressing glyoxalase enzymes (NtglyI) and (NtglyII) either alone or together at 200 mM NaCl maintained higher GSH levels under salt stress. These plants showed enhanced basal activity of GSH and suffered minimal salinity stress induced oxidative damage measured as lipid peroxidation [97]. In contrast, Jaleel et al. [98] reported that treatment of salinity at 80 mM concentration to Catharanthus roseus resulted in decreased GSH content when compared with control plants. A number of reports are available on the response of GSH to salt stress, but its mode of action in coping salt stress needs elaborative studies. The integrated knowledge on physiological, biochemical and molecular approaches may provide deeper insights towards the precise roles and mechanism of GSH involved in salt stress tolerance.
Lipoic acid (LA) is derived from octanoic acid, contains two S atoms connected by a disulfide bond at C-6 and C-8 of its chemical structure and is soluble in both water and lipid phase. Its antioxidative property and protective nature, both in reduced form as dihydrolipoic acid (DHLA) and oxidised form as LA have been suggested [99]. Bast and Haenen [100] reported that DHLA can reduce GSH and can be a good tool for detoxification of ROS in a stressful environment. However, it can also have pro-oxidant properties by its iron-reducing ability and by its ability to generate S-containing radicals that can damage proteins [101]. Perez-Lopez et al. [102] have shown that the presence of LA in barley leaves occurred in the reduced as well as in oxidized forms under non-salinized and salinized conditions. They found that non-salinized conditions represent LA that was present mainly in reduced form i.e. DHLA and was partly involved in the tolerance of barley to elevated salinity. In the same report, the effects of salinity were mitigated and were correlated to a higher constitutive level of LA, which was believed to improve regeneration of DHA, and GSSG under salinity. Tarchoune et al. [103] explained the role of LA and DHLA in salinity tolerance in basil (Ocimum basilicum). They have shown that salt tolerance was dependent on cultivar and salt exposure time as adaptive response was associated with higher constitutive amounts of tocopherols with a maximum ability of roots to use DHLA to maintain growth making basil tolerant to salinity.
Glucosinolates are a class of secondary metabolites having S as integral part of its structure and mainly found in Brassica crops like oilseed rape, broccoli and cabbage. Although, physiological significance of glucosinolates in plants is not clear but it is assumed that glucosinolates has a role in plant-pathogen interaction [104]. Besides pathogen interaction, glucosinolates metabolism has role in tolerance of environmental factors including saline condition. Glucosinolates accumulation in response to environmental stress has been studied showing that some environmental factors can change the glucosinolates content and changes in the glucosinolate profile under stress is variable for plant adaptation. López-Berenguer et al. [105] showed the influence of salinity level for glucosinolates content and found that salinity level at 40 mM increased total glucosinolates content while it decreased at 80 mM concentration of salt stress suggesting that glucosinolates have potential to versatile role under salt stress. This variation in the glucosinolates content may be related to the developmental stage of plants [106]. It has been suggested that increased glucosinolates was involved in osmotic adjustment and salt tolerance with low water potential in boroccoli plants [107]. The glucoerucin (a type of glucosinolates) content in broccoli sprouts was increased by NaCl treatment whereas total glucosinolates reduced under salt stress. Salt treatment at 60 mmol/L for 5 d maintained higher biomass and comparatively higher content of glucosinolates in sprouts of broccoli [108]. Allocation of glucosinolates could be one of the tolerance strategies to cope with stressful conditions with relative low energy cost. A direct allocation of glucosinolates production in Arabidopsis with an increase in photosynthetic energy has been reported by Bekaert et al. [109]. These all findings indicated that salt-induced increases in glucosinolates content may be involved in the salt stress response of plants but the effects of salinity on glucosinolates biosynthesis and metabolism need further attention at molecular and cellular levels. However, the biological function of a particular glucosinolate in specific tissues requires more investigation.
From the above discussion it is evident that S metabolism and S metabolites are essential to plants under optimal as well as under salt stress. Sulfur incorporated into methionine and cysteine in plants directly impacts the nutritional value of human food and livestock feeds. The goal of this review was to improve our understanding on the importance of S metabolites as a link between S assimilation and to the salt stressed plants. The review covered how S metabolites are useful in salt tolerance in plants by inducing physiological and molecular processes.
The increasing salinity is an imminent threat to soil fertility and agricultural productivity. In plants, S assimilates provide structural components of essential cellular molecules besides acting as signaling molecules for environmental communication. S-containing compounds can enhance salinity tolerance by modulating physiological and molecular processes and by up-regulating genes for stress tolerance. The use of S as fertilizer or soil supplementation with S-containing compounds in agriculture system can emerge as a good tool to combat salinity effects. It is important to carry out detailed studies on S assimilation pathway, its control through internal and external factors and to gain more physiologically relevant insight into plant responses to their environment. Study on sulfur metabolites (amino acids, vitamins, Thioredoxin system, glutathione, lipoic acid and glucosinolates) have been proved much effective in tolerance of salt stress by maintaining or modulating physiological and molecular processes. Currently, vitamins and lipoic acid have received attention in research for their role in abiotic stress tolerance, but the studies are still in its fancy. The study on biotin and lipoic acid seems to be important as they act as cofactor in many metabolic processes. Overall, strategies should be focused on the regulation of S assimilation pathway and production of thiols with further concentration on subcellular compartmentation of thiols in relation to control on salinity stress.
Authors are thankful to the University Grants Commission and Department of Science and Technology, Government of India, New Delhi of financial assistance for the work.