Journal of Pharmaceutical Care & Health Systems
Open Access
ISSN: 2376-0419
ISSN: 2376-0419
Research Article - (2014) Volume 1, Issue 4
This study evaluated the effects of one or two injection of differents concentrations (0.2 and 0.04μg/μL) of extracts obtained from the salivary glands of female ticks Rhipicephalus sanguineus (Acari: Ixodidae) (Latreille, 1806) also called “brown dog tick” fed for 2 days on the rabbit hosts on the morpho-physiology of the leg musculature of female Wistar rats inoculated with Walker 256 tumor cells. The number of leukocytes and the creatinine levels were quantified for individuals from all groups. The results of both histology and electron microscopy revealed that a single, low-concentration injection of the extract (0.04μg/μL) was more effective in containing tumor invasion and caused less ‘collateral damage’ to the muscle tissue, which was the object of this study. The results also revealed that creatinine levels were higher in rats subjected to both one and two injections of the extract at a concentration of 0.04μg/μL than in those subjected to one and two injections of the extract at the higher concentration (0.2μg/μL), suggesting that in the first group, injection of the extract contributed to maintaining the integrity of the muscle tissue. With regard to the number of leukocytes, the results suggested that in all the inoculated rats (Walker 256 cells), there was a significant increase in the total number of leukocytes. The inoculated rats that received both one and two injections of the extract at a concentration of 0.2μg/μL experienced a significant increase in the number of leukocytes compared with those inoculated but not exposed to the extract; this result can be explained by the fact that beyond the tumor cells, the extract itself acted to boost the defense response at this concentration. However, inoculated rats subjected to injections (one and two) of the extract at a concentration of 0.04μg/μL showed a significant decrease in the total number of leukocytes compared with rats that were only inoculated and those inoculated and injected with the extract at a concentration of 0.2μg/μL. These results reinforced that the extract at the 0.04μg/μL concentration not only acted more effectively to inhibit Walker 256 tumor cells but also did not act as a stressor, because the number of leukocytes was lower. Therefore, the data obtained here indicated that the same molecules or a pool of molecules produced by the salivary glands of ticks belonging to this species have the ability to inhibit tumor growth, while minimizing ‘collateral damage’ to the body.
Keywords: Ticks; Salivary gland extracts; Wistar rats; Muscle; Morphophysiology
The saliva of ticks is a complex mixture and is considered to be a large and effective pharmacological arsenal that guarantees the nutrition and survival of these ectoparasites [1,2] through the production of substances with diverse actions. Within this secretion are found bioactive agents that can operate mainly by modulating the immune, inflammatory and hemostatic systems of the host [1,3]. Among these, it is possible to highlight the presence of components of glycoproteins and lipoproteins origin, in addition to lipid components [4], calreticulin, acid phosphatase, esterases, amino peptidases, metalloproteinase, lipocalins and prostaglandins [1,4-7]. For this reason, the saliva of ticks in general has been the subject of various studies, mainly because of the great interest in the composition and mode of action of these bioactive agents, some of which have been previously described as vasodilators, anticoagulants, immunosuppressant [8] and, more recently, antiangiogenic [2,9] and anti-tumor agents [10-13].
With specific regard to the dog brown tick belonging to the species Rhipicephalus sanguineus, studies have demonstrated that the biochemical complexity of the saliva undergoes changes as the blood feeding progresses because of the need of ectoparasites to modulate the homeostatic system of the host, allowing the ectoparasites to be biologically successful. This mixture that makes up the saliva of R. sanguineus ticks can undergo both quantitative and qualitative variations considering specific periods of their glandular cycle, such as its beginning at 2 days of feeding, the middle at 4 days, and the end at 6 days [14,15].
The literature brings previous studies that found a protein participating in coagulation processes that has cytotoxic activity on tumor cell lines in the saliva of Amblyomma cajennense ticks, which prefer horses as their hosts [11]. In addition, studies by [10] have demonstrated that different molecules present in extracts from the salivary glands of other tick species have the capacity to inhibit the growth of HeLa lineage cells either by inducing apoptosis (R. appendiculatus and A. variegatum) or because of their anti-proliferative action (Ixodes Ricinus, R. appendiculatus, Dermacentor reticulatus and A. variegatum). Furthermore, [12] also reported that the final saliva of some species of ticks can act as an in vitro control for tumor cells, which begin to die as a result of exposure. Finally, [16] reported the inhibitory effects of tick saliva on the migratory and invasive capacity of MDA-MB-231 cells.
The above information is the product of a new perspective on the study of ticks, which no longer considers them pests and harmful organisms but an important source of bioactive agents with the potential to impact the control of tumor cell line development and, consequently, tumor growth [17,18]. However, many studies on this topic have yet to be undertaken using the diverse technical tools that are now available.
Therefore, taking this into consideration, the present study aimed to analyze the effects of the extract obtained from the salivary glands of female R. sanguineus ticks fed for 2 days on rabbit hosts on the morphophysiology of the hind leg musculature of Wistar rats previously inoculated with Walker 256 tumor cells in vivo.
In vivo tumor model study
This in vivo study used 35 female Wistar rats with an average weight of 450 g–500 g and aged 90–120 days old. The rats were distributed into the following study groups: a) Control 1 (CG1; 5 healthy individuals), b) Control 2 (CG2; 5 individuals with a tumor treated with sodium phosphate buffer, pH 7.4), c) Control 3 (CG3; 5 individuals with a tumor only), d) Test 1 (TG1; 5 individuals with a tumor, treated with an injection of the extract SGE2 at a concentration of 0.04μg/μL), e) Test 2 (TG2; 5 individuals with a tumor, treated with two injections of the extract SGE2 at a concentration of 0.04μg/μL), f) Test 3 (TG3; 5 individuals with a tumor, treated with an injection of the extract SGE2 at a concentration of 0.2μg/μL) and g) Test 4 (TG4; 5 individuals with a tumor, treated with two injections of the extract SGE2 at a concentration of 0.2μg/μL).
Collection and maintenance of walker 256 tumor cells
The Walker 256 tumor cells were kindly provided by Professor Maria Cristina Cintra Gomes Marcondes, PhD, Supervisor of the Laboratory of Nutrition and Cancer, Department of Physiology IB/ UNICAMP, Campinas, SP, Brazil. The rats inoculated with the cells were euthanized, the tumor tissue was fragmented and filtered in gauze and the collected liquid was transferred into 50 mL Falcon tubes, which were centrifuged at 4°C for 10 min at 3000 rpm. All procedures were performed at 4°C. After centrifugation, the supernatant was discarded and the precipitate resuspended in 0.9% saline solution, thereby forming a concentrated cell suspension.
To count the cells, a 1:100 dilution was made using 0.9% saline solutions. Next, 200μL of the diluted suspension was mixed with 200μL of 1% trypan blue dye. The cells were counted using a light microscope in a Neubauer chamber. Cell viability was determined considering the fact that viable cells would be refractive to light. The number of cells in the working suspension was 1×107 cells/mL.
After counting the cells, 1 mL of the working suspension was inoculated intraperitoneally into the abdominal region of three adult Wistar rats to obtain the tumor liquid (ascites). After approximately 15 days of inoculation, the ascites was collected from the abdomens of these rats.
Implantation of walker 256 tumor cells
The cancer model used in this study was described by [19]. In this model, the rats with ascites were euthanized using a mixture of ketamine and xylazine. After the ascitic fluid was collected, it was placed in tubes containing EDTA and diluted to a ratio of 1:100 in sodium phosphate buffer, pH 7.4. The cells were then counted using trypan blue dye, as described previously. The suspension, with 1×106 cells/mL, was then inoculated in the right intraplantar posterior leg (300μL suspension/paw).
Obtaining salivary glands and SGE2 glandular extracts
To obtain female ticks after 2 days of feeding, male and female fasting ticks were used; these were obtained from R. sanguineus colonies that were kept especially for the purpose of this study in a BOD chamber under controlled conditions (29°C, 80% humidity and a 12 h photoperiod) in the Animal Facility in the Department of Biology, UNESP, Rio Claro, SP- Brazil.
Females feeding since 2 days after prior infestation were removed from the hosts using circular movements with surgical forceps in the hypostome region. Next, these individuals were placed in Petri dishes containing saline for dissection (7.5 g NaCl, 2.38 g Na2HPO4, 2.72 g KH2PO4 and 1000 mL of distilled water) to remove the salivary glands.
The salivary glands were placed into Eppendorf tubes containing 50μL of phosphate buffer, pH 7.4, and macerated. The tubes were then centrifuged for 30 min at 10,000×g, and the collected supernatants were placed into previously sterilized Eppendorf tubes.
Within the pre-sterile vertical laminar flow chamber, the extract was filtered with the aid of 0.22μm and 13 mm diameter sterile filter units (JBR610303 disposable filter unit, Millex GV Durapore PVDF membrane, Millipore, MilliUni) attached to hypodermic syringes, following which proteins were dosed (Bradford method).
Next, the concentrated extract was stored at −20°C until further use.
The SGE2 extracts were diluted with sodium phosphate buffer, pH 7.4, to obtain concentrations of 0.04μg/μL and 0.2mg/μL.
The procedure for injecting different concentrations of the SGE2 extract was conducted in a closed and sterile area in the Animal Facility of the Department of Biology, Institute of Biosciences at UNESP campus, Rio Claro, SP- Brazil.
The first injection of different concentrations of the SGE2 extract was administered 15 days after tumor cell inoculation, with the 0.04μg/ μL extract injected into Test groups 1 (TG1) and 2 (TG2) and the 0.2μg/ μL extract injected into Test groups 3 (TG3) and 4 (TG4).
On the same day, a single application of sodium phosphate buffer, pH 7.4, was injected into Control group 3 (CG3).
The second injection of extracts was administered 4 days after the first, only in Test group 2 (TG2), with the SGE2 0.04μg/μL extract, and Test group 4 (TG4), with the SGE2 0.2μg/μL extract.
On day 21 after inoculation of the tumor cells and injection of the extract, the animals were euthanized using an overdose of ketamine and xylazine.
Next, in a sterile environment, the muscle was removed and prepared for analysis.
Conventional light microscopy
Harris haematoxylin and aqueous eosin technique: The materials were fixed with 4% paraformaldehyde in 1.5 mL plastic micro tubes for a period of 24 h. The samples were then washed three times with 0.1M sodium phosphate buffers, pH 7.4. Dehydration occurred in an ascending ethanol series of 70%, 80%, 90% and 95% (baths, 30 min each). Subsequently, the material was soaked in resin for 24 h and embedded in plastic molds containing polymerizing resin until the blocks hardened.
After this process, the blocks were sectioned on a microtome to obtain 3-μm-thick sections, which were then collected on glass slides.
The slides, which contained approximately six sections each, were stored in an oven and subsequently stained with haematoxylin for 7 min. After washing for 4 min in running water, they were then stained with aqueous eosin for 5 min and washed in running water again. After air-drying, the slides were quickly dipped in xylene, covered with Canada balsam and mounted with a cover slip.
The slides obtained were examined and photographed under a light microscope.
Transmission Electron Microscopy (TEM)
To perform routine TEM analysis, muscle fragments were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate (pH 7.2) buffer for 2 h at room temperature. Then, two washes were performed in 0.1 M sodium cacodylate buffer for 15 min each, and the fragments were immediately post-fixed in a solution of 1% osmium tetroxide (OsO4) for 2 h. The samples were then washed twice for 15 min in 0.1 M sodium cacodylate buffer. The material was washed in 10% ethyl alcohol for 15 min, contrasted in 1% uranyl acetate solution and dissolved in 10% alcohol for 2 h in the dark. This was followed by dehydration in a graded series of ethanol (70%–100%, lasting 15 min each) and pure acetone (lasting 15 min). The material was embedded in resin + acetone at a 1:1 ratio for 24 h and embedded in pure resin. It was then placed in a 60°C oven for 72 h for polymerization.
The blocks were sectioned on an ultra-microtome and the sections were placed on copper grids. They were subsequently contrasted in uranyl acetate solution for 45 min, rinsed in water and contrasted in lead citrate for 15 min and washed again in water and 0.02 M sodium hydroxide (NaOH) (adapted from (20)).
After the procedure, the material was examined under the Philips CM 100 Transmission Electron Microscope at the Laboratory of Electron Microscopy, Department of Biology, IB-UNESP, Rio Claro, SP- Brazil.
Clinical examinations
For the clinical examination, blood was collected from animals at the time they were euthanized. This blood was stored in heparinized tubes and later sent for laboratory analysis of the following parameters: quantification of leukocytes and creatinine levels.
The values obtained for these parameters were statistically analyzed by ANOVA with Tukey’s post hoc test, and differences with p < 0.05 were considered statistically significant.
This study was submitted to the Institute of Biosciences, UNESP, Rio Claro, SP- Brazil, Ethics Committee on Animal Use (CEUA) and was duly approved with process number 3537.
Microscopic evaluation of the in vivo action of the extracts on the tumor model
Haematoxylin-Eosin staining technique control group 1: To compare the results obtained herein, the control subjects, i.e., those not inoculated with tumor cells (Figure 1A, C), served as a model for the description of the histological organization of skeletal muscle under normal conditions. Descriptions by [21] were also considered.
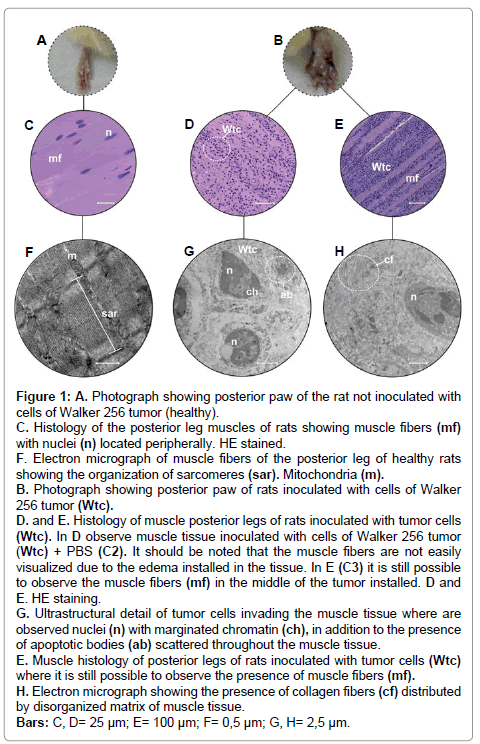
Figure 1: A. Photograph showing posterior paw of the rat not inoculated with cells of Walker 256 tumor (healthy). C. Histology of the posterior leg muscles of rats showing muscle fibers (mf) with nuclei (n) located peripherally. HE stained. F. Electron micrograph of muscle fibers of the posterior leg of healthy rats showing the organization of sarcomeres (sar). Mitochondria (m). B. Photograph showing posterior paw of rats inoculated with cells of Walker 256 tumor (Wtc). D. and E. Histology of muscle posterior legs of rats inoculated with tumor cells (Wtc). In D observe muscle tissue inoculated with cells of Walker 256 tumor (Wtc) + PBS (C2). It should be noted that the muscle fibers are not easily visualized due to the edema installed in the tissue. In E (C3) it is still possible to observe the muscle fibers (mf) in the middle of the tumor installed. D and E. HE staining. G. Ultrastructural detail of tumor cells invading the muscle tissue where are observed nuclei (n) with marginated chromatin (ch), in addition to the presence of apoptotic bodies (ab) scattered throughout the muscle tissue. E. Muscle histology of posterior legs of rats inoculated with tumor cells (Wtc) where it is still possible to observe the presence of muscle fibers (mf). H. Electron micrograph showing the presence of collagen fibers (cf) distributed by disorganized matrix of muscle tissue. Bars: C, D= 25 μm; E= 100 μm; F= 0,5 μm; G, H= 2,5 μm.
Histologically, this type of muscle was composed of elongated cells, also called muscle fibers, which were characteristically multinucleate with peripheral nuclei and a flattened and fusiform morphology (Figure 1C). In the cytoplasm of these cells, it was possible to observe striations in both the longitudinal and transverse directions, with the former corresponding to the arrangement of the myofibrils and the latter corresponding to the organization of the thick and thin myofilaments, thus forming the sarcomeres, which are considered the contractile units of muscle fibers (Figure 1C, F).
Still related to the histological organization of this type of muscle, each cell or muscle fiber was surrounded by thin connective tissue called the endomysium. This set of cells then formed the muscle bundle covered externally by the perimysium, a layer of connective tissue comprising collagen and elastin that grouped the muscle fibers, and finally, the grouped bundles formed the muscles, which were in turn coated externally with the epimysium, a layer of connective tissue containing interlaced collagen fiber bundles.
This same histological pattern was observed in the muscles of the right posterior leg of the individuals in the Control 1 group, i.e. animals not subjected to inoculation with tumor cells that showed the same histological type, i.e. elongated and multinucleate fibers presenting both longitudinal and transverse striations (Figure1C). The presence of conjunctive tissue was observed between the muscle fibers. The nuclei present here were also fusiform and were arranged in the periphery of the fibers (Figure 1C).
Confirming the histological results, Figure 1F shows the organization of the striated skeletal muscle at the electron microscope level, where the sarcomeres are evident, mainly by the presence of transverse striations (arrangement of thick and thin myofilaments). Furthermore, fusiform, flattened and peripheral nuclei show uncondensed chromatin.
Control group 2
The individuals in this group were inoculated with cells from the Walker 256 tumor lineage + PBS. In the right posterior leg muscle tissue, extensive invasion of cells was seen. These cells were histologically characterized by hypertrophy, irregular borders, pleomorphism (variation in size and shape) and the presence of more than one nucleus, which in turn were irregular in shape, were very large in size and contained condensed chromatin (Figure 1D). These tumor cells filled practically all the space formerly occupied by the connective tissue between the muscle fibers, leaving these indistinguishable in histological sections and characterizing total disruption of this tissue (Figure 1D). In the ultra structural level, the morphology of these tumor cells can be observed, primarily highlighting the nucleus/ cytoplasm ratio of these cells (Figure 1G). The injected PBS probably also caused changes in the extracellular matrix, swelling the tissue as can be seen in Figure 1D.
In general, after performing these procedures, it was observed that the muscle tissue exhibited complete histological disorganization and also showed the presence of eosinophilic masses as a result of the large number of tumor cells, the morphological characteristics of which were previously described above (Figure 1D). Another feature in this group was the histological observation (confirmed by electron microscopy) of tumor cells with strongly stained irregular nuclei and with nuclei in the process of chromatin marginalization (1G). Electron microscopy also showed the presence of nuclei in the process of fragmentation has as well as apoptotic bodies (Figures 1G).
Control group 3
The results obtained from the individuals in this study group, which underwent only inoculation with the Walker 256 tumor cell line, showed an intermediate histology between that observed in individuals from the Control 1 and 2. This means that there was extensive invasion of tumor cells in muscle tissue, mostly in the extracellular matrix, which led to extensive disruption of the tissue (Figure 1E). However, although this disruption is quite apparent, it is possible to observe muscle fibers, a situation contrary to that observed in the Control 2 group.
In the intercellular regions, cells with pleomorphic characteristics, pyknotic nuclei and irregular shapes were observed (Figure1H). In these same regions, collagen fibers could be clearly observed by electron microscopy to have not only disrupted but also undergone fragmentation, thus differing from the typical aspect of the intercellular matrix observed in skeletal muscle (Figure 1H ).
In addition, it was no longer possible to view the typical transverse striations of the muscle fibers in the histological sections, indicating disorganization as a result of neoplastic invasion (Figure 1E).
Test group 1
Individuals in this group, in addition to inoculation with tumor cells from the Walker 256 lineage, also received an injection of SGE2 extract at a concentration of 0.04μg/μL (Figure 2). The results showed that the fibers, although still histologically disorganized, show certain integrity. Unlike the previous situation, they can be observed in a parallel arrangement among themselves, with their nuclei located peripherally, and even feature the presence of transverse striations (Figure 3A–D). In the previously described group, i.e., Control 3, besides the histological disorganization, the muscle fibers themselves were internally disrupted, a fact confirmed by the absence of transverse striations as well as the presence of pyknotic nuclei and, in some cases, nuclei in the process of fragmentation (Figure 1E, H).
Figure 2: A. C. D. Illustrative diagrams showing: A. Location of salivary gland (sg) in tick. C and D. Injection of two days (SGE2) extracts at concentrations 0.04 and 0.2 ug/uL in rats. B. Total salivary gland (sg) where it can be observed the organization in acinus with rounded shape.
One detail observed very often in this study group was the presence of cells in the process of degranulation (Figure 3C, D), most likely because of the presence of the toxic element in the muscle. This degranulation was quite similar to that occurring with mast cells in cases of immediate hypersensitivity reactions.
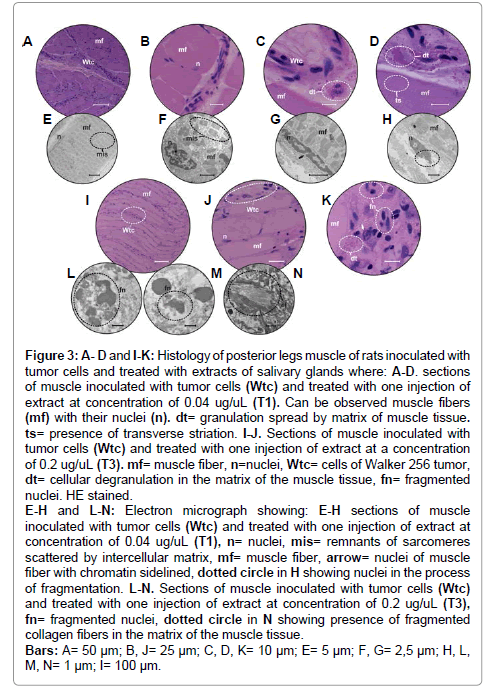
Figure 3: A- D and I-K: Histology of posterior legs muscle of rats inoculated with tumor cells and treated with extracts of salivary glands where: A-D. sections of muscle inoculated with tumor cells (Wtc) and treated with one injection of extract at concentration of 0.04 ug/uL (T1). Can be observed muscle fibers (mf) with their nuclei (n). dt= granulation spread by matrix of muscle tissue. ts= presence of transverse striation. I-J. Sections of muscle inoculated with tumor cells (Wtc) and treated with one injection of extract at a concentration of 0.2 ug/uL (T3). mf= muscle fiber, n=nuclei, Wtc= cells of Walker 256 tumor, dt= cellular degranulation in the matrix of the muscle tissue, fn= fragmented nuclei. HE stained. E-H and L-N: Electron micrograph showing: E-H sections of muscle inoculated with tumor cells (Wtc) and treated with one injection of extract at concentration of 0.04 ug/uL (T1), n= nuclei, mis= remnants of sarcomeres scattered by intercellular matrix, mf= muscle fiber, arrow= nuclei of muscle fiber with chromatin sidelined, dotted circle in H showing nuclei in the process of fragmentation. L-N. Sections of muscle inoculated with tumor cells (Wtc) and treated with one injection of extract at concentration of 0.2 ug/uL (T3), fn= fragmented nuclei, dotted circle in N showing presence of fragmented collagen fibers in the matrix of the muscle tissue. Bars: A= 50 μm; B, J= 25 μm; C, D, K= 10 μm; E= 5 μm; F, G= 2,5 μm; H, L, M, N= 1 μm; I= 100 μm.
Electron microscopy observations showed that when the frequency of tumor cell invasion decreased, it did not prevent the muscle fibers from damage (Figure 3E-H). In this sense, the damage was primarily related to misalignment of the sarcomeres (Figure 3E, F), changing the morpho-physiology of the tissue as a whole. Besides the fibers, the nuclei also underwent changes such as chromatin marginalization and, most likely, the beginning of fragmentation (Figure 3F-H).
Test group 2
In this group, individuals were inoculated with tumor cells and also received two injections of SGE2 extract at a concentration of 0.04μg/μL (Figure 2). The muscle histology of these individuals also shows an apparent disorganization of the muscle fibers, which can be observed parallel to each other, with peripheral nuclei that are fusiform and distinct (Figure 4A-D). However, unlike the previous group, the presence of transverse striations was rare, and longitudinal arrangements characterizing the parallel arrangements of the myofibrils also presented severe changes, because they seemed to be separated from each other by a space (not stained by the technique applied) as well as the consequent formation of vacuole-like structures (Figure 4C, D). Compared with that in the individuals from the previous group (Figure 3A-D), the intensity in the cells in the process of degranulation was less frequent; these cells were located between the spaces in the extracellular matrix (Figure 4D).
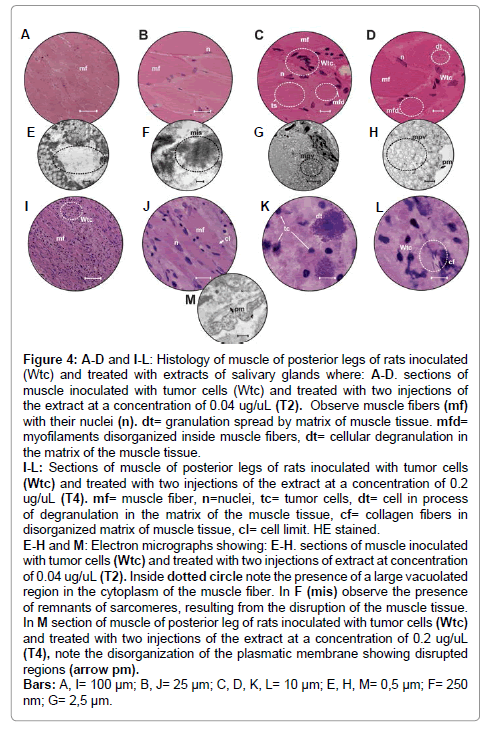
Figure 4: A-D and I-L: Histology of muscle of posterior legs of rats inoculated (Wtc) and treated with extracts of salivary glands where: A-D. sections of muscle inoculated with tumor cells (Wtc) and treated with two injections of the extract at a concentration of 0.04 ug/uL (T2). Observe muscle fibers (mf) with their nuclei (n). dt= granulation spread by matrix of muscle tissue. mfd= myofilaments disorganized inside muscle fibers, dt= cellular degranulation in the matrix of the muscle tissue. I-L: Sections of muscle of posterior legs of rats inoculated with tumor cells (Wtc) and treated with two injections of the extract at a concentration of 0.2 ug/uL (T4). mf= muscle fiber, n=nuclei, tc= tumor cells, dt= cell in process of degranulation in the matrix of the muscle tissue, cf= collagen fibers in disorganized matrix of muscle tissue, cl= cell limit. HE stained. E-H and M: Electron micrographs showing: E-H. sections of muscle inoculated with tumor cells (Wtc) and treated with two injections of extract at concentration of 0.04 ug/uL (T2). Inside dotted circle note the presence of a large vacuolated region in the cytoplasm of the muscle fiber. In F (mis) observe the presence of remnants of sarcomeres, resulting from the disruption of the muscle tissue. In M section of muscle of posterior leg of rats inoculated with tumor cells (Wtc) and treated with two injections of the extract at a concentration of 0.2 ug/uL (T4), note the disorganization of the plasmatic membrane showing disrupted regions (arrow pm). Bars: A, I= 100 μm; B, J= 25 μm; C, D, K, L= 10 μm; E, H, M= 0,5 μm; F= 250 nm; G= 2,5 μm.
The areas invaded by the neoplastic cells were less frequent than those observed in Test Group 1 (Figure 4A-D). In these areas, the eosinophilic masses persisted, and it was impossible to observe the limits of the cells, only irregular nuclei and cells in the process of fragmentation (Figure 4C).
Although the neoplastic invasion was decreased, ultra structural analysis showed severe damage to the fibers and intercellular matrix. This damage could be translated into the presence of extensive areas of cytoplasmic degradation (Figure 4E), the presence of fragments of sarcomeres resulting from misalignment and fragmentation of these structures (Figure4F). With regard to the alterations, the membrane of the fibers was completely disorganized and the cytoplasm filled with micropynocitic vesicles (Figure 4G,H) namely, portions of the extracellular fluid that were encompassed and brought into the cell. Furthermore, granular and electron dense material was observed between the vesicles (Figure 4 E,H).
Test group 3
In this group, the individuals were inoculated with tumor cells and also received one injection of SGE2 extract at a concentration of 0.2μg/ μL (Figure 2). The histology of the muscle tissue of these individuals showed that, even in the presence of the extract, major invasion of tumor cells existed throughout the muscle fibers (Figure 3I-K), which was also confirmed by ultra-structural analyses (Figure 3LM). However, these fibers still maintained their parallel arrangement (Figure 3I-J). Transversal and longitudinal striations could not be observed (Figure3J), and the nuclei of the muscle fibers appeared with condensed chromatin (Figure 3I-J); furthermore, in some regions, these nuclei appeared to be undergoing fragmentation (Figure 3K).
The neoplastic cells were observed with greater frequency than that in the Test groups described above and occurred in the intercellular matrix (Figure 3L-M). Here, cells forming an eosinophilic mass could also be observed, where the cells were pleomorphic, featured pyknotic nuclei and chromatin marginalization, and were in the process of fragmentation (Figure 3K). Ultra structurally, it was possible to observe complete disorganization of the intercellular matrix, where beside this fragmented collagen fibers (Figure3N), the presence of very electron dense nuclear fragments could be observed (Figure3L-M).
In this group, the presence of granules distributed by the intercellular matrix could also be observed, probably the result of cell degranulation (Figure 3K).
Test group 4
The individuals in this group were inoculated with tumor cells and also received two injections of SGE2 extract at a concentration of 0.2μg/ μL (Figure 2). In general, the histological results showed muscular tissue completely invaded by the tumor cells (intercellular matrix), which disorganize the fibers (Figure 4I-J). This histology was similar to that observed in the individuals inoculated with the tumor cells but not subjected to extract injection. The muscle fibers did not present the typical arrangement; i.e., transverse striations were not seen (Figure 4J), and the histology suggested that the membrane (endomysium) of these fibers was altered, in the sense that in some regions, detachment from the peripheral cytoplasm (Figure4J) could be observed (results corroborated by ultra structural analysis- Figure 4M).
Furthermore, the nuclei, which should be arranged peripherally, now seemed to change their location on the basis of the disorganization exhibited by these cells (Figure 4J). Many of these muscle cells featured accentuated pyknosis, but some seemed to have dispersed chromatin (Figure 4J). Neoplastic areas were more frequent (Figure 4I-L) in comparison with those in specimens from Test Group 3. Within this mass, neoplastic cells with characteristics of mast cells undergoing degranulation were also observed (Figure 4K).
In the intercellular matrix, it was also possible to observe the presence of very small eosiniphilic structures resembling short filaments, which were probably collagen fibers, distributed among the tumor cells (Figure 4L).
Statistical analysis of blood parameters of the rats
The results obtained from clinical analysis of the blood of the rats used in this study showed that creatinine levels were significantly increased (1.15 ± 0.07) in rats subjected to one and two injections of the extract at a concentration of 0.04 μg/uL compared with those in rats subjected to one and two injections of the extract at a concentration of 0.2 μg/uL (0.45 and 0.55, respectively, ± 0.071) p < 0.01 (Table 1 and Figure 5).
| Control 1 | Control 2 | SGE2 | ||||
|---|---|---|---|---|---|---|
| Healthy | With tumor and submitted to 1 injection of PBS | 0,04μg/μL | 0,2μg/μL | |||
| 1 injection | 2 injections | 1 injection | 2 injections | |||
| Parameters | ||||||
| Creatinine(mg/dL) | 0,75 ± 0.0707 | 0,95 ± 0.0707 | 1,15 ± 0.0707(*,**,1) | 1,15 ± 0.0707(*,1) | 0,45 ± 0.0707(*,2) | 0,55 ± 0.0707(**,1) |
| Leukocytes(n°/μL) | 5899.5 ± 0.7071 | 12699.5 ± 0.7071(*,1) | 7699.5 ± 0.7071(*,**,1) | 6299.5 ± 0.7071(*,**,1) | 21699.5 ± 0.7071(*,**,1) | 26599.5 ± 0.7071(*,**,1) |
Table 1: Results of biological parameters: creatinine level and leukocytes count.
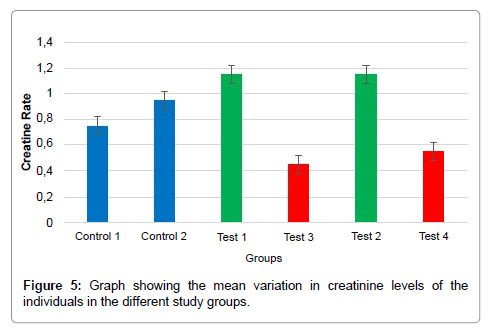
Figure 5: Graph showing the mean variation in creatinine levels of the individuals in the different study groups.
The results related to the quantity of leukocytes showed that in all the rats inoculated with the tumor cells (12699.5; 7699.5; 6299.5; 21699.5; 26599.5 ± 0.7), there was a significant increase in the total number of leukocytes compared with the healthy rats (5899.5± 0.7), while there was a significant increase in rats inoculated and subjected to one (21699.5 ± 0.7) and two (26599.5 ± 0.7) injections of the extract at a concentration of 0.2 μg/uL compared with those that were only inoculated (12699.5 ± 0.7) p < 0.01 (Table 1; Figure 6).
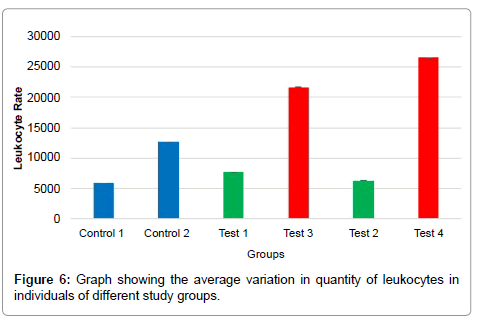
Figure 6: Graph showing the average variation in quantity of leukocytes in individuals of different study groups.
The search for substances that have an inhibitory action on the growth of tumor cells has generally been ongoing, because these cells have morphological and physiological characteristics that make them special and difficult to control. On the other hand, the scarcity of information available in the literature with respect to the countless differences found in the salivary secretions produced by the various genera and species of ticks. Reinforcing these information the in vitro study performed by Furquim et al (personal information) showed that gland extracts compromised the viability of tumor cells of lines DH 82 and HL-60 by inducing apoptosis as well as their proliferation rate. In this sense these factors were determinating to motivated us to perform this study, which was primarily focused on demonstrating the potential action of salivary gland extracts from female R. sanguineus ticks fed for 2 days on rabbits hosts on Walker 256 tumor cells inoculated in vivo into the right posterior leg of Wistar rats.
The results obtained in the present study demonstrated that the invasion of tumor cells via inoculation was very aggressive, and in a matter of 2 days, because of the high rate of proliferation of these cells, almost the entire intercellular inter fibers space (the muscle cells are also called muscle fibers, because they are long rather than wide) of the targeted musculature was occupied. This occupation led to significant morphological alterations in the muscle, such as disorganization of both the tissue as well as the muscle fibers themselves, which certainly damaged the physiology of this muscle. It was evident that the muscle fibers themselves lost their typical morphological characteristic, namely the repeated sequence of uniform bands along the length of the cells [22], which is the origin of both the transverse striations (arrangement of sarcomeres) and the longitudinal striations (arrangement of contractile filaments), which have already been comprehensively described in the literature [21].
The invasion of tumor cells in the muscle tissue promoted the emergence of eosinophilic masses, which pushed and shifted the muscle fibers from their place of origin. Because movements to contract and relax the muscles must be synchronized, this disorganization certainly makes it impossible for the dynamics of muscular physiology to occur with precision.
The assays performed in this study resulted in different answers according to the concentrations of the extracts used (0.2 and 0.04μg/ μL) and the number of injections (one or two) administered. The results showed that animals receiving only one injection of the 0.04μg/ μL extract had a lower rate of tumor cell invasion, because the inter fibers spaces held smaller and less frequent eosinophilic masses, as well as the presence of defense cells (leukocytes in general) and active mast cells (degranulation). The presence of active mast cells in tissue invaded by tumor cells, according to [23] and [21], would indicate that this process, when in inflammatory exudates, would mediate the cellular and vascular responses of the tissue, and in different microenvironments, it would have effects both promoting and inhibiting tumors [24]. In addition, according to other authors, mast cells would be closely associated to tumor progression via promotion of angiogenesis [25].
In general, the results obtained with this concentration of extract were very similar to those for Control Group 1 (healthy rats). On the other hand, animals that received two injections of the extract at this same concentration presented, in addition to an increased number of tumor cells, clear disorganization of the muscle fibers (which were already visible at the light microscopy level) represented by the absence or small presence of transverse striations, meaning that the sarcomeres had been disorganized and the presence of inter myofibrilar spaces, which certainly compromised the physiology of the muscle tissue.
With regard to the results obtained for the 0.2μg/μL concentration (regardless of the number of injections), it was found that in both situations, there were significant morphological and physiological changes in the muscle tissue, with the following observations: intense invasion of tumor cells in the intercellular matrix and extensive disorganization of the muscle fibers. It should be noted that despite the fact that two tests detected damage to the tissue, where the animals were subjected to two extract injections, the areas of invasion of tumor cells were much larger, showing that the extract was unable to contain them.
Therefore, the results obtained here indicated that, although ‘collateral damage’ was detected, the injection of different concentrations of extracts from the salivary glands of female R. sanguineus ticks fed for 2 days on rabbit hosts promoted a decrease in the rate of invasion of Walker 256 lineage cells in the musculature of Wistar rats in vivo, representing a possible strategy for controlling the disordered growth of tumor cells.
It was clear that the single, lower-concentration injection of the extract was more effective in containing tumor invasion and caused less ‘collateral damage’ to the muscle tissue, which was the object of this study.
Alongside the morphological and histological bioassays, the following parameters were also clinically evaluated in the blood of the animals: creatinine levels and quantification of leukocytes. The former was evaluated because there is a direct relationship between creatinine kinase (enzyme responsible for the degradation of creatinine) levels and muscle injuries [23,26-28], while the latter was evaluated because of the involvement of leukocytes in the inhibition of tumor cells (defense behavior).
The results revealed that creatinine levels were higher in rats subjected to one and two injections of the extract at a concentration of 0.04μg/μL compared with those in rats subjected to one and two injections of the extract at 0.2μg/μL, suggesting that in the first group, the injection of the extract contributed to maintaining the integrity of the muscle tissue. This hypothesis is supported by the fact that there was no statistically significant difference in the parameter in question when comparing the results of rats that received injections of 0.04μg/ μL and those only inoculated with tumor cells, demonstrating that the extract did not affect the muscular morphology and physiology.
With regard to the increase/decrease in the number of leukocytes, the results suggested that, in all the rats inoculated with tumor cells, there was a significant increase in the total number of leukocytes, because neoplastic cells acted as an antigen stimulating the defense response. Furthermore, in the inoculated rats receiving injections (one and two) of the extract at a concentration of 0.2μg/μL, there was a significant increase in the number of leukocytes compared with that in rats only inoculated and not exposed to the extract. This result can be explained by the fact that beyond the tumor cells, the extract itself acted to boost the defense response at this concentration. However, inoculated rats subjected to injections (one and two) of the extract at a concentration of 0.04μg/μL showed a significant decrease in the total number of leukocytes compared with rats that were only inoculated and those inoculated and injected with the extract at a concentration of 0.2μg/uL. These results reinforced that the 0.04μg/μL concentrations was not only more effective in inhibiting the tumor cells but also did not act as a stressor, because the number of leukocytes was lower.
Therefore, the data obtained in this study through in vivo tests using Walker 256 tumor cells and exposing them to different concentrations of extracts obtained from the salivary glands of female R, sanguineus ticks with 2 days of feeding on rabbit hosts indicated that molecules produced by the salivary glands of this species of ticks have the ability to inhibit tumor growth, as well as minimize collateral damage to the organism. It should be noted here that in vivo study models, unlike in vitro models, introduce the variable of the body’s physiological response when exposed to stressors (in this case, tumor cells and salivary gland extracts) and that tick saliva in general is considered to be a reservoir of bioactive modulators of the immune-inflammatory system.