Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2017) Volume 5, Issue 1
Intertidal epilithic microalgae play very important role as feed of higher organisms. Taxonomical composition of epilithic algae and their seasonal dynamics in relation with environmental factors were studied at middle coast of Saudi Arabia, namely Yanbu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea. Temperature, salinity and pH varied from 22.10-32.02°C, 38.72-39.35 psu, and 8.40-8.63, respectively throughout the study. A total of 87 epilithic microalgal species belonging to 4 families was identified: 79 Bacillariophyceae, 5 Cyanophyceae, 2 Dinophyceae, and 1 Cryptophyceae. The epilithic microalgal abundance varied from 2.32 × 105 cells/L to 7.66 × 105 cells/L during the study with an average of 4.03 × 105 cells/L. The lowest and highest abundance of epilithic microalgae occurred in winter and summer at Mastorah and Rabigh, respectively. Among the microalgae taxa, diatoms were the dominant group which was 91.01%, followed by Cyanophyceae (5.62%), Dinophyceae (2.25%) and Raphidophyceae (1.12%). Among diatom, the centric and pennate diatoms were 74.07% and 25.93%, respectively throughout the study. The occurrence frequency analysis revealed that high frequently observed epilithic microalgae were Licmophora flabellata, L. paradoxa, L. remulus, Navicula vula, N. incerta and N. transitans throughout the study. Therefore, it could be said that diatoms are the major group of epilithic microalgae which show high abundant in summer at the northwest coast of Jeddah, Saudi Arabia, the Red Sea. Second dominant group was Cyanophyceae, and the dominant species was Spirulina subsalsa among epilithic microalgal community.
<Keywords: Epilithic microalgae; Environmental factors; Seasonal dynamics; Jeddah coast; The Red Sea
Microalgae perform a major part of primary production and support food webs from small pond to oceans. Microalgae composition with environmental factors is important to understand the structure and dynamics of any marine ecosystem. Microalgal populations fluctuate from season to season and from region to region due to the interaction among physical, chemical and biological variables [1,2]. Among the microalgae, the benthic microalgae play an important role in intertidal biomass production [3,4]. The intertidal flats are the sites of intense biological activity and often with high productivity [5] though intertidal benthic microalgae have to survive in a harsh environment. To distinguish the groups of benthic organism, different terms such as bioflim, benthos, periphyton, epilithon have been used by several researchers [6]. Aquatic system, these epilithon can comprise the most abundant producers. The epilithon grow on stones and rocks which are generally considered as an important base of the food chain [7]. Natural substrates present advantages to epilithic microalgae because the stones supply nutrients that support community development. Diatoms are the major components of epilithon community because of their continuous presence with different abundances along the aquatic system and their quick response to the environmental changes although the term epilithon encompasses a great diversity of microalgal taxa [8].
A study was conducted on epilithic microalgal composition and dynamics at Jeddah Coast. Jeddah is a Saudi Arabian city located at the coast of the Red Sea (21.50° N 39.1667° E). The Red Sea is a unique area in the tropics for several reasons which having the highest salinity water body in the world’s seas with the deep water salinity value of 40.06 psu while the surface water salinity values ranged from 40.10- 40.65 psu [9,10]. The high salinity phenomenon occurs due to its location in an arid and hot climatic zone, isolation from open ocean, without riverine inputs, the high evaporation rate (>210 cm yr-1), and scant rainfall restricted from October to May, resulting in a negative hydrological budget. The hydrological deficit is compensated by inflows from the Indian Ocean through Bab-Al-Mandeb. The Red Sea is seasonally influenced by variable wind-driven monsoon currents such as southwest monsoon in summer (May to September) and north to northwest winds blow on whole axis of the Red Sea in winter. The Red Sea surface waters pile up from December to February along the African coast and some upwelling occurs along the Arabian coast [9], and reversal of summer condition because of the south-southeast winds only affect the southern half of the Red Sea. These two circulation patterns seasonally converge and surface waters sink at Jeddah Coast. In these zones, coastal water downwelling occurs due to the warming of surface layer and nutrient-deficient water layer. Downwelling reduces biological productivity and transports heat, dissolved materials, and dissolved oxygen rich surface waters in to greater depths. The convergence zone is classified as the poorer zone in microalgal species [11].
The Red Sea is considered as an oligotrophic water body due to deficient of several major nutrients such as nitrate, ammonium, phosphate, and silicate [12]. The required nutrients for photosynthetic activity have to come through water intrusion (from the Gulf of Aden), the sub-surface (below the nutricline), or via aerial deposition. Higher nutrient concentrations occur in the southern areas, and gradually decline towards the north. The reservoirs of nutrients in the Red Sea are trapped below the stratified zone due to the persistent pycnocline. The deep water renewal is generally prevented by high stratification levels and the resulting homogeneity of the northern Red Sea deep waters which limits nutrient availability and biological productivity in the euphotic zone. Jeddah is the second biggest city of Saudi Arabia, located at the Coast off the Red Sea and is extending rapidly. It receives huge waste water from fish farms, industries and domestic use which may increase the nutrients concentration. A varying supply of nutrients, among other factors, permits microalgae species richness to increase beyond the number of limiting nutrients, while changes in nutrients are important drivers of change in species composition of primary producers [13,14]. The field studies have shown that episodic nutrient flushing and loading can result in enhancing microalgae species diversity [15-17]. Moreover, temperature values have been increasing abruptly since mid-90’s to present, therefore, the oceanic warming may have a direct and/or indirect impact on intertidal microalgae. Studies of macro and microalgae are very few at the Jeddah coast though algae are an important source of food and shelter for aquatic organisms and also indicator of pollution. Presence of new microalgal species was studied at Obhur creek of Jeddah coast [18]. Recently some studies have been performed to monitor the effect of environmental factors on chlorophyll and microalgae abundance near desalination plant, marine fouling and polluted area of Jeddah coast [19,20]. Studies dealing with littoral benthic microalgal ecology are rare compared to those done on water column throughout the world [3,4], and in the Red Sea, the study on benthic microalgae in intertidal zone, especially on epilithic microalgae has not been done. The present study was conducted to monitor and interpret the structure and seasonal dynamics of epilithic microalgae in relation with environmental factors at northwest coast of Jeddah, Red Sea.
Study area, sampling and sample analysis
The sampling was done seasonally at the middle time of season which was in March (spring), June (summer), September (autumn) and December (winter) in 2015. The sampling was conducted at the northwest coast of Jeddah, Saudi Arabia, the Red Sea. The sampling sites were: (i) Yanbu (23°59´45″N and 38°10´35″ E) that is a major Red Sea port in the Al Madinah province of western Saudi Arabia. It is approximately 300 Km northwest of Jeddah. There are oil refineries and petrochemical industry; (ii) Ar-Ryias (23°33´55″N and 38°36´17″E) is an industrial city of Madinah province; (iii) Mastorah beach city (23°08´80N and 38º 47´14″E) which is famous for fishing; (iv) Rabigh (22°41´35″N and 3900´34″E) that has several industries such as a factory of cement, electricity station, large refinery, Petro Rabigh; and (v) Thuwal (22°16´44″N and 39º 05´21″E) that had long been a fishing center. The town is experiencing an increase in industries due to its proximity to the King Abdullah Economic City (Figure 1).
Figure 1: Location of the study sites; Northwest coast of Jeddah, Saudi Arabia, namely Yambu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea.
15 pieces of gravel stones were collected from each sampling station, then the entire surface area of each stone was cleaned and washed with tooth brush and filtered autoclaved natural seawater, respectively to get the microalgae attached on stones. The whole stones collected from each station were washed three times. Finally, the samples of stones washed and filtered water volume was made 0.5 L and was preserved with Lugol’s solution of 2% and was kept for further study. Additionally, the surface area of the stones was estimated with digital vernier slide caliper after collection of eipilithic microalgae.
For taxonomical study, the preserved sample was observed under the phase-contrast microscope (Zeiss Axioplan, Germany) at 400X magnification. Taxonomic identifications were made with reference to Prescott [21-24]. Specfies diversity index (H) was calculated accordingly [25]. For quantitative study, a 1 mL preserved sample was taken after mixing and counted in a Sedgewick-Rafter (S-R) counter chamber with same microscope as mentioned above. Counted results were summarized as cells per liter, and used for graphical presentation. To estimate the concentration level of nutrients (NO3-N, NO2-N, NH3-N, PO4-P and SiO3-S), the water sample was collected from the respective stations. The concentration of NO3-N, NO2-N, NH3-N, PO4-P and SiO3 were determined with a spectrophotometer (Shimadzu UV-1201, Japan) according to the methods described by Parsons et al. [26]. The water temperature, salinity and pH were measured on spot with portable meter HACH (Model HQ14d). Correlations between the abundance of epilithic microalgae and environmental factors were determined following MS-Excel statistical program.
Dynamics of physical factor
The water temperature showed three distinct seasonal cycle characterized by a maximum in summer-autumn, followed by spring and winter. Temperature values varied from 22.10-32.02°C with an average of 28.48°C, showing minimal and maximal in winter and summer at Yanbu and Rabigh, respectively (Figure 2). Salinity values fluctuated from 38.72-39.35 psu with an average of 38.92 psu. The salinity value was lowest in spring at Ar-Rayis, and highest was in summer at Yanbu (Figure 3). The pH values varied from 8.40 to 8.63 throughout the study with an average of 8.52. The highest and the lowest pH values were recorded at Rabigh and Mastorah. The highest pH value was counted in spring followed by winter pH values among all the stations (Figure 4). Among nutrients, the concentrations levels of NO3-N, NO2-N, NH3-N, PO4-P and SiO3 were (0.26-2.96, 0.25- 3.24, 0.13-3.18, 0.43-1.47 and 0.25-8.35) μg/L, respectively among the stations of northwest coast of Jeddah, the Red Sea, from winter to autumn 2015 (Table 1).
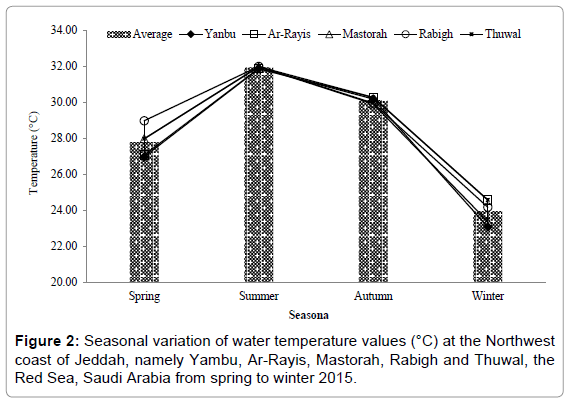
Figure 2: Seasonal variation of water temperature values (°C) at the Northwest coast of Jeddah, namely Yambu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, Saudi Arabia from spring to winter 2015.
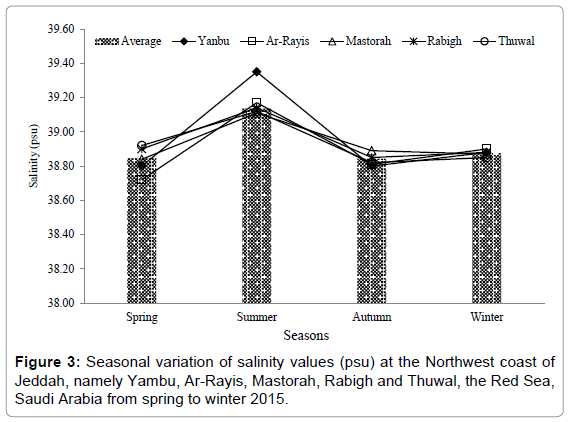
Figure 3: Seasonal variation of salinity values (psu) at the Northwest coast of Jeddah, namely Yambu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, Saudi Arabia from spring to winter 2015.
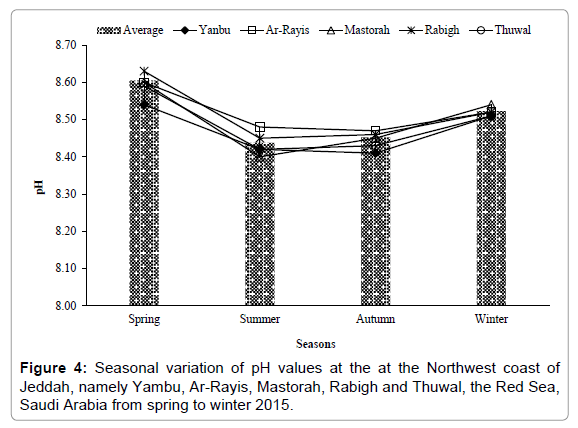
Figure 4: Seasonal variation of pH values at the at the Northwest coast of Jeddah, namely Yambu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, Saudi Arabia from spring to winter 2015.
| Stations | NO3-N | NO2-N | NH3-N | PO4-P | SiO3-S | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spr. | Sum. | Aut. | Win. | Spr. | Sum. | Aut. | Win. | Spr. | Sum. | Aut. | Win. | Spr. | Sum. | Aut. | Win. | Spr. | Sum. | Aut. | Win. | |
| Yanbu | 0.57 | 0.95 | 1.43 | 0.68 | 0.22 | 3.02 | 0.45 | 0.63 | 0.78 | 0.27 | 0.53 | 0.38 | 0.67 | 1.11 | 0.49 | 0.73 | 1.10 | 5.79 | 1.47 | 0.26 |
| Ar-Rayis | 1.02 | 0.83 | 1.27 | 0.72 | 0.39 | 1.40 | 0.52 | 0.75 | 0.44 | 0.20 | 0.52 | 0.42 | 0.83 | 0.85 | 0.52 | 0.42 | 2.06 | 4.91 | 1.81 | 0.24 |
| Mastorah | 0.34 | 1.91 | 2.76 | 1.00 | 0.28 | 1.36 | 0.55 | 0.71 | 0.18 | 0.27 | 0.41 | 1.09 | 0.54 | 1.43 | 1.02 | 0.71 | 2.11 | 5.31 | 2.62 | 1.22 |
| Rabigh | 0.24 | 0.84 | 0.54 | 2.41 | 0.25 | 1.77 | 0.53 | 0.71 | 0.44 | 0.23 | 0.64 | 1.11 | 0.64 | 0.87 | 0.57 | 0.51 | 2.06 | 7.05 | 1.02 | 1.32 |
| Thuwal | 0.61 | 0.91 | 1.37 | 2.30 | 0.40 | 1.93 | 0.65 | 0.91 | 0.55 | 0.37 | 0.46 | 2.18 | 0.48 | 1.05 | 0.64 | 0.49 | 3.08 | 6.13 | 1.80 | 1.53 |
Table 1: Dynamics of nutrients (μg.L-1)at the Northwest coast of Jeddah, Saudi Arabia, namely, Yanbu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, from spring to winter 2015.
The abundance values of epilithic microalgae showed positive correlation with temperature values in winter (r=0.50), spring (r=0.79) and summer(r=0.87). Similarly, the above epilithic microalgae abundance values also showed positive correlation with salinity values in spring (r=0.75) and autumn (r=0.67). The abundance values of epilithic microalgae showed strongly positively correlation with NO3-N (r=0.97) in winter whereas it was highly negatively correlated in spring NO3-N (r=-0.79). The concentration values of NH4-N showed positive correlation with abundance values of epilithic microalgae in winter (r=0.67), spring (r=0.65) and in autumn (r=0.54). SiO2-S and abundance values of microalgae were positively correlated in autumn (r=0.96) and winter (r=0.51).
Taxonomic composition
Taxonomic community composition of epilithic algal assemblages and their seasonal dynamics were studied for the first time at northwest coast of Jeddah, Saudi Arabia, the Red Sea. A total of 88 epilithic microalgal species were identified, among them 78 Bacillariophyceae, 6 Cyanophyceae, 3 Dinophyceae, 1 Raphidophyceae (Table 2). Among Bacillariphyceae, the abundance levels of pennate and centric diatoms were 58 and 20 species, respectively throughout the study. The variation in species diversity index value (H) ranged from 1.35 to 3.35 with an average of 2.40 from spring to winter 2015. The lowest and highest H values were at Yanbu in autumn, and in spring at Rabigh (Table 3).
| Microalgae name | Winter | Spring | Summer | Autumn |
|---|---|---|---|---|
| Bacillariophyceae | ||||
| Pennate diatoms | ||||
| Amphiprora Ehrenberg | C | |||
| A. alata Kützing | R | |||
| A. paludosa Smith | R | |||
| Amphora sp. Ehrenberg | X | |||
| Amphora arcus Gregory | R | R | ||
| A. coffeaeformis Kützing | C | R | ||
| A. decussata Grunow | F | H | ||
| A. exigua Gregory | R | R | ||
| Bacillaria paradoxa Gmelin | R | R | ||
| B. paxillifer Müller | R | R | ||
| Closterium closterioides Ralfs | X | |||
| Cocconeis placentula Ehrenberg | H | |||
| Cocconeis scutellum Ehrenberg | C | X | ||
| Cylindrotheca closterium Ehrenberg | X | H | R | X |
| Entomoneis punctulata Grunow | R | |||
| Fragilaria capucina Desmazières | H | C | ||
| Grammatophora marina Lyngbye | X | |||
| Gyrosigma sp. | C | |||
| G. acuminatum Kützing | X | |||
| G. balticum Ehrenberg | C | |||
| G. fasciola Ehrenberg | C | |||
| Licmophora sp. | R | |||
| L. abbreviata Agardh | R | X | ||
| L. ehrenbergii Kützing | C | H | ||
| L. flabellata Agardh | H | H | H | C |
| L. gracilis Ehrenberg | H | X | ||
| L. grandis Kützing | C | |||
| L. hyalina Kützing | H | X | ||
| L. paradoxa Lyngbye | R | H | H | H |
| L. remulus Grunow | H | C | H | H |
| Navicula delicatula Cleve | H | C | R | |
| N. distans Smith | C | X | R | |
| N. granii Jørgensen | H | |||
| N. incerta Grunow | C | H | H | H |
| N. peregrine Ehrenberg | H | |||
| N. ramosissima Agardh | H | |||
| N. transitans Grunow | C | H | H | R |
| N. vula Carter | H | H | H | H |
| Nitzschia sigma Kützing | R | |||
| N. seriata Cleve | X | |||
| N. hungarica Grunow | H | R | X | C |
| N. longissima Brébisson | X | X | ||
| Pseudo-nitzschia pungens Hasle | R | H | C | |
| N. socialis Gregory | H | |||
| Pinnularia gibba Ehrenberg | H | |||
| P. viridis Ehrenberg | R | R | ||
| Pleurosigma normanii Ralfs | X | |||
| P. angulatum Smith | X | R | C | X |
| P. formosum Smith | X | |||
| P. intermedium Smith | H | |||
| Pseudo-nitzschia fraudulenta Cleve | C | |||
| P. lineola Cleve | X | |||
| Stauroneis anceps Ehrenberg | C | |||
| Staurosirella pinnata Grunow | C | |||
| Striatella unipunctata Lyngbye | X | |||
| Synedra ulna Nitzsch | X | |||
| Tabellaria fenestrate Lyngbye | X | |||
| Toxonidea insignis Donkin | C | |||
| Centric diatoms | ||||
| Biddulphia alternans Bailey | R | |||
| C. costatus Pavillard | C | |||
| C. curvisetus Cleve | X | |||
| C. laciniosus Schütt | C | |||
| C. nodulifer Schmidt | H | |||
| C. radiates Ehrenberg | R | |||
| C. socialis Lauder | X | |||
| C. wailesii Gran & Angst | R | X | ||
| Chaetoceros affinis Lauder | X | |||
| Coscinodiscus centralis Ehrenberg | X | |||
| Cyclotella sp. Kützing | X | |||
| Guinardia delicatula Cleve | X | |||
| L. adriaticus Schroder | R | |||
| L. danicus Cleve | X | H | X | |
| Lauderia annulata Cleve | C | |||
| Leptocylindrus minimus Gran | R | |||
| Melosira granulate Ehrenberg | X | |||
| R. setigera Brightwell | H | R | X | |
| Rhizosolenia delicatula Cleve | X | |||
| Thalassionema bacillare Heiden | R | X | ||
| Dinophyceae | ||||
| Bysmatrum arenicola Horiguchi | R | |||
| Cerataulus turgidus Ehrenberg | ||||
| Prorocentrum lima Ehrenberg | H | H | ||
| Cryptophyceae | ||||
| Rhodomonas salina Wislouch | H | |||
| Cyanophyceae | ||||
| Merismopedia angularis Thompson | H | R | ||
| Microcystis aeruginosa Kützing | ||||
| Oscillatoria formosa Gomont | C | |||
| O. limosa Gomont | X | F | H | |
| Spirulina subsalsa Gomont | R | C | X | |
| Trichodesmium sp. Ehrenberg | C | H | R | |
Table 2: Epilithic microalgal taxa list and occurrence frequency at the Northwest coast of Jeddah, Saudi Arabia, namely Yanbu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, from spring to winter 2015.
| Season | Yanbu | Ar-Rayis | Mastorah | Rabigh | Thuwal |
|---|---|---|---|---|---|
| Spring | 2.56 | 2.39 | 3.29 | 3.35 | 3.07 |
| Summer | 1.89 | 2.31 | 2.28 | 2.42 | 2.08 |
| Autumn | 1.80 | 2.23 | 2.47 | 2.39 | 1.93 |
| Winter | 2.38 | 1.35 | 2.29 | 2.92 | 2.67 |
Table 3: Epilithic microalgal diversity index value (H) at Northwest coast of Jeddah, Saudi Arabia, namely Yanbu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, from spring to winter 2015.
Abundance dynamics of epilithic microalgae
The epilithic microalgae abundance value varied from 2.32 × 105 cells/L to 7.66 × 105 cells/L during the study with an average of 4.03 × 105 cells/L. The lowest and highest abundance value of epilithic micoralgae occurred in winter and summer at Mastorah and Rabigh, respectively. The second highest abundance value of epilithic micoralgae was 7.20 × 105 cells/L at Mastorah and the third highest was 5.42 × 105 cells/L also at Thuwal in summer, respectively (Figure 5). Among the microalgae taxa, diatoms were the dominant groups which were 91.01%, followed by Cyanophyceae (5.62%), Dinophyceae (2.25%) and Raphidophyceae (1.12%). Among diatoms, the centric and pennate diatoms were 74.07% and 25.93%, respectively throughout the study. Among diatoms, the frequence analysis of microalgal species occurrence revealed that epilithic microalgal species were Licmophora flabellata, L. paradoxa, L. remulus, Navicula vula, N. incerta and N. transitans throughout the study. In winter above 15% total epilithic microalgal species contributed the highest abundance value to the epilithic microalgal species communiities. Licmophora flabellata contributed the highest abundance value which was 63%, followed by Amphora coffeaeformis (50%) and Chaetoceros curvisetus (37%) to the epilithic micoralgal communities. In spring, 8 species were found to be dominated above 15%, and the most dominant species was Toxonidea insignis which contributed 43%, followed by Licmophora remulus (28%) and Navicula ramosissima (27%) of epilithic micoralgal communities. Similarly, 6 and 5 species were found to be abundant above 15% in summer and autumn, respectively. Navicula delicatula was found to be 79%, followed by, Licmophora remulus (51%) and Navicula transitans (26%) among epilithic micoralgae in summer. Licmophora flabellata was found to be majority among epilithic micoralgal communities which was 50%, followed by Microcystis aeruginosa (39%) and Trichodesmium sp. (29%) in autumn (Table 4).
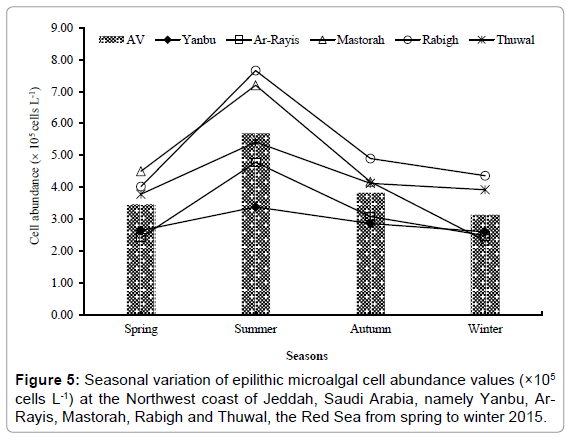
Figure 5: Seasonal variation of epilithic microalgal cell abundance values (×105 cells L-1) at the Northwest coast of Jeddah, Saudi Arabia, namely Yanbu, Ar- Rayis, Mastorah, Rabigh and Thuwal, the Red Sea from spring to winter 2015.
| Species | Yanbu | Ar-Rayis | Mastorah | Rabigh | Thuwal |
|---|---|---|---|---|---|
| Spring | |||||
| Leptocylindrus donicus Cleve | 12 | 18 | |||
| Licmophora grandis Kützing | |||||
| L. remulus Grunow | 19 | ||||
| Licmophra sp. | |||||
| Merismopedia angularis Thompson | 12 | 13 | |||
| Navicula delicatula Cleve | 10 | ||||
| N. longissima Brébisson | 14 | ||||
| N. transitans Grunow | 12 | ||||
| Prorocentrum lima Ehrenberg | 15 | ||||
| Toxonidea insignis Donkin | 16 | ||||
| Summer | |||||
| Amphora sp. | 10 | ||||
| Chetoerros socialis Lauder | 15 | ||||
| Cylindrotheca closterium Ehrenberg | 15 | ||||
| Gyrosigma acuminatum Kützing | 14 | ||||
| Licmophora remulus Grunow | 45 | ||||
| Merismopedia angularis Thompson | 20 | ||||
| Navicula delicatula Cleve | 19 | 17 | |||
| N. transitans Grunow | 27 | 17 | 15 | ||
| N. distans Smith | 17 | ||||
| Nitizschia seriata Cleve | 15 | ||||
| N. sigma Kützing | 14 | 15 | |||
| Plearosigma angulatum Smith | 13 | ||||
| Spirulina subsalsa Gomont | 18 | ||||
| Autumn | |||||
| Coscinodiscus wailesii Gran & Angst | 11 | ||||
| Gyrosigma fasciola Ehrenberg | 13 | ||||
| Licmophora flabellata Agardh | 11 | 44 | |||
| L. gracilis Ehrenberg | 11 | ||||
| Microcystis aeruginosa Kützing | 39 | 14 | |||
| Navicula distans Smith | 11 | ||||
| Nitzschia hungarica Grunow | 28 | 11 | |||
| Oscillatoria Formosa Gomont | 17 | ||||
| Pleurosigma normanii Ralfs | 10 | ||||
| Pseudo-nitzschia lineola Hasle | 11 | ||||
| Rhizosolenia setigera Brightwel | |||||
| Trichodesmium sp. Ehrenberg | 12 | 25 | |||
| Winter | |||||
| Amphiprora alata Kützing | 14 | ||||
| Amphora exigua Gregory | 16 | 14 | |||
| Chaetoceros curvisetus Cleve | 31 | ||||
| Licmophora abbreviate Agardh | 22 | 14 | |||
| L. ehrenbergii Kützing | 10 | ||||
| L. flabellata Agardh | 62 | ||||
| L. paradoxa Gmelin | 12 | ||||
| Navicula granii Jørgensen | 10 | ||||
| Nitzschia hungarica Grunow | 12 | 10 | |||
| N. longissima Brébisson | 12 | ||||
| Spirulina subsalsa Gomont | 17 | ||||
| Stauroneis anceps Ehrenberg | 16 | ||||
Table 4: List of dominant microalgae species (above 10% abundance) at the Northwest coast of Jeddah namely, Yanbu, Ar-Rayis, Mastorah, Rabigh and Thuwal, the Red Sea, Saudi Arabia from spring to winter 2015.
Epilithic microalgae communities can range from filamentous green algae to silicaceous diatoms to blue-green algae [4,27]. They are very important source of primary producer and contribute energy cycle in inland water and coastal water ecosystem [28]. As epilithic microalgae species of Bacillariophyceae were dominant, followed by Cyanophyceae, Dinophyceae, and Chlorophyceae to the epilithic microalgae at the northwest coast of Jeddah. The average abundance values of those epilithic microalgal species community was high in summer, followed by spring, winter and autumn in the study area. A fluctuation of epilithic species diversity and biomass production can occur due to environmental conditions, geological nature, grazing, resources limitation, habitat disturbance, substrate availability. Moreover, inorganic pollutants can be constrained to epilithic microalgae species diversity, so that the pollutants could consider a limiting factor for growth and dominancy of epilithic microalgal species.
The highest abundance values of the epilithic microalgae were in summer which might be related with the water temperature values, moderate to higher concentration levels of NO3-N and SiO3 nutrients in correlation with high temperature values might have created a suitable substrate for proliferation and growth of epilithic microalgal species communities, since epilithic microalgae abundance values were high in summer. The abundance of diatoms was the dominant group among epilithic microalgae throughout the study. Similarly, diatoms are the dominant group of microalgae in Arabian Gulf and Shatt Al-Arab estuary [11,29,30]. Agusti and Sanchez [31] reported that temperature values and nutrients (N, P and Si compounds) concentrations level, diatoms at different regimes is related with their adaptive ability. Abundance values are controlled by the environmental factors and their adaptive ability, although diatom grazing is another important controlling factor [32].
Among diatoms, the different taxa were found to be dominant in different seasons and different stations. The small size diatoms were found to be occurred and dominated commonly in different sampling stations throughout the year. Among diatoms, two species of pennate diatoms and three species of centric diatoms were found to be dominant during the study. Among pinnate diatoms, Licmophora spp., and Navicula spp., were found to be dominant. Licmophora flabellata was 61.78% of total epilithic microalgal community at Ar-Rayis in spring (Table 4). Abundance of Navicula spp., was high in summer and the highest cell abundance value was for Navicula transitans which was 27% of epilithic microalgae species community at Yanbu (Table 4).
Temperature values were identified as the most effective parameter on the epilithic microalgae abundance values and species composition variation in the Homa Lagoon Izmir, Turkey where Navicula species were dominant [33,34]. Three taxa of centric diatoms were found to be available throughout the study, which were Chaetoceros spp., Coscinodiscus spp., and Leptocylindrus spp.
Among centric diatoms, only Chaetoceros spp., contributed 30.02% to the epilithic microalgae as a dominant species in winter. The species composition and abundance values of all microalgae follow seasonal temperature cycle patterns in the Red Sea [35] that Dowidar et al. [36] studied on water column microalgal species communities and found that diatom was dominant in summer. Similarly, among epilithic microalgal communities, diatoms are the dominant group and they reach at an abundance peak in summer. There were Dinophyceae and Cyanophyceae and they did not show any dominancy though two species (Spirulina subsalsa and Trichodesmium spp.) of Cyanophyceae contributed high cell abundance among epilithic microalgal community. The epilithic microalgal abundance followed the seasonal pattern of temperature and salinity as their abundance was high in summer when the salinity and temperature were also high. The diversity of epilithic microalgae was high in spring in all stations.
This study contributed to the knowledge improvement of the epilithic microalgal species communities at Jeddah, the Red Sea, in Saudi Arabia, by providing the information of seasonal dynamics of taxonomical composition and abundance in relation with environmental factors from the study. It could be concluded that diatoms were the most dominating group of epilithic microalgae and their abundance values became high in summer.
This research was supported by the 2015 deanship research project No.1434/150/355 scientific promotion program funded by King Abdulaziz University, Jeddah.