Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2014) Volume 5, Issue 1
Forty mature catfish of both sexes (n=10 per season; 5 males and 5 females) were used to study the effect of different seasons on the histological and histochemical structure of the gonads. The histological results showed that both testes and ovaries of the catfish were degenerated during winter that was considered as a resting season of the catfish gonadal activity. Both testes and ovaries began to restore their intact and fully mature structure during spring and continue the same during summer where the testes show distended seminiferous lobules with all spermatogenic cells and spermatozoa also, the ovaries showed the different developmental stages including mature follicles therefore, both spring and summer were considered as spawning season of the catfish. During autumn, both testes and ovaries appeared as spent gonads where the testes showed many empty seminiferous lobules and the ovaries showed many atretic follicles therefore, autumn was considered as post-spawning or spent season. The results of Gonado-Somatic Index (GSI) were coincided with the histological structure of the gonads where they show peak value during spring and summer (spawning season) and showed the lowest value during winter (resting season).
Keywords: Histology; Testis; Ovary; Catfish; Seasons
Catfish is considered as the cheapest source of high quality animal protein and rich in calcium, phosphate, iodine and vitamins [1]. Generally, fish is not only used for human consumption, but also used as a good source of animal meal [2]. The study of the gonads of the different teleost fish attracts the attention of several investigators in the African catfish, Clarias gariepinus; in Bagrus docmac and Bagrus bayad; in Nile tilapia, Oreochromis niloticus; in mullet, Mugil cephalus [3-9]. The purpose of this work was to study the effect of different seasons of the year on the histological and histochemical structure of the gonads of the catfish.
Animals
A total of 40 mature catfish of both sexes (n=10 per season; 5 males and 5 females) were collected alive from Toukh fish market in Kalyubia governorate, Egypt during the different seasons of the year (2010- 2012). Winter season in Egypt extends from December to February, spring extends from March to May, summer extends from June to August, and autumn extends from September to November. The fish were captured for marketing from River Nile in Al-Qanater, Kalyubia governorate, Egypt. The fish were captured for marketing from River Nile in Al-Qanater, Kalyubia governorate, Egypt.
Preparation of tissue specimens
The catfishes were transported alive to the Histology and Cytology Department, Faculty of Veterinary Medicine, Benha University. Immediately, the fish was weighted, decapitated and the gonads were removed and weighed after opening the fish belly. After weighing, the gonads were immediately fixed in Bouin’s solution for 24 hours, dehydrated through ascending grades of ethanol, cleared in xylene, embedded in paraffin wax and sectioned at 5 μm. The sections were stained with Haematoxylin and Eosin, Periodic acid Schiff (PAS), and Crossman’s trichrome. Fixative and staining methods were used as outlined by Bancroft et al. [10].
The Gonado-Somatic Index (GSI)
The GSI (according to Dougbag et al.) was used for following up the seasonal variations in the gonads by the formula:
Weight of gonad
GSI = ×100 [11]
Fish body weight
Our results revealed that the gonads of catfish revealed variable structure according to seasons of the year.
Testis
During winter, testis showed very thick tunica albuginea and interstitial connective tissue and showed degenerated spermatogenic cells except spermatogonia (Figure 1). During spring, testicular lobules showed all spermatogenic developmental stages (Figure 2) and continue the same structure during summer where testicular lobules were filled with spermatozoa also, showed thin tunica albuginea and interstitial connective tissue compared to (Figure 3). During autumn, many testicular lobules appeared empty from sperematozoa while, other spermatogenic cells were present and began to degenerate (Figure 4).
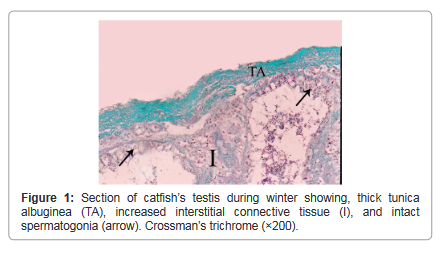
Figure 1: Section of catfish’s testis during winter showing, thick tunica albuginea (TA), increased interstitial connective tissue (I), and intact spermatogonia (arrow). Crossman’s trichrome (×200).
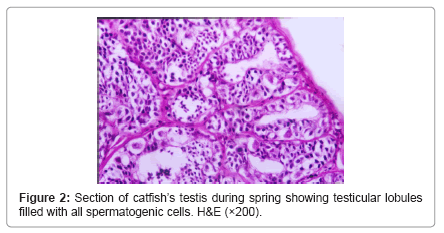
Figure 2: Section of catfish’s testis during spring showing testicular lobules filled with all spermatogenic cells. H&E (×200).
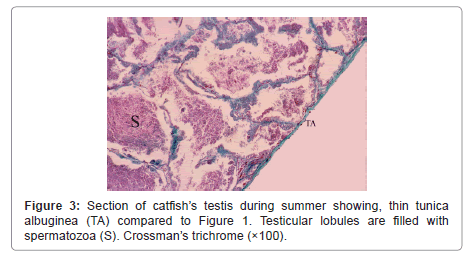
Figure 3: Section of catfish’s testis during summer showing, thin tunica albuginea (TA) compared to Figure 1. Testicular lobules are filled with spermatozoa (S). Crossman’s trichrome (×100).
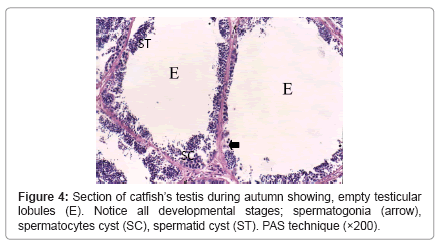
Figure 4: Section of catfish’s testis during autumn showing, empty testicular lobules (E). Notice all developmental stages; spermatogonia (arrow), spermatocytes cyst (SC), spermatid cyst (ST). PAS technique (×200).
Ovary
During winter, ovary showed very thick tunica albuginea and interstitial connective tissue and showed many degenerated follicles which mainly were of pre-vitellogenic type (Figure 5). During spring, ovary showed all the developmental stages but, the common stages were vitellogenic and post-vitellogenic ones (Figure 6) and continue the same structure during summer where post-vitellogenic (mature) follicles were usually seen, the ovary showed thin tunica albuginea (Figure 7). During autumn, many atretic follicles began to appear and the pre-vitellogenic follicles began to increase in number again (Figure 8).
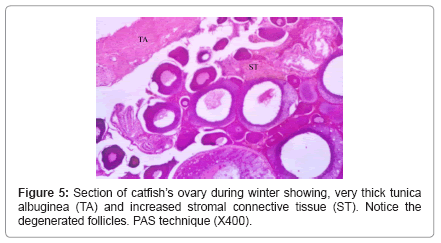
Figure 5: Section of catfish’s ovary during winter showing, very thick tunica albuginea (TA) and increased stromal connective tissue (ST). Notice the degenerated follicles. PAS technique (X400).
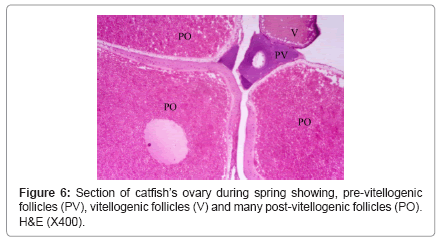
Figure 6: Section of catfish’s ovary during spring showing, pre-vitellogenic follicles (PV), vitellogenic follicles (V) and many post-vitellogenic follicles (PO). H&E (X400).
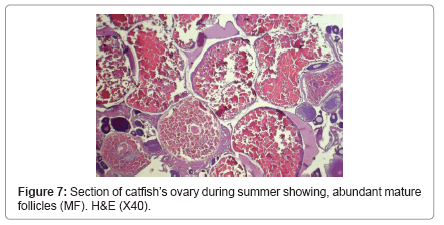
Figure 7: Section of catfish’s ovary during summer showing, abundant mature follicles (MF). H&E (X40).
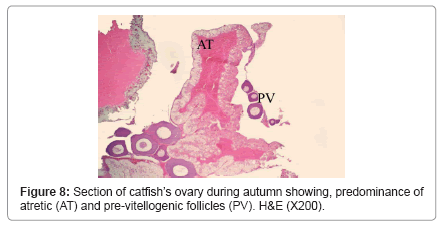
Figure 8: Section of catfish’s ovary during autumn showing, predominance of atretic (AT) and pre-vitellogenic follicles (PV). H&E (X200).
The results of GSI were coincided with the obtained histological results where the GSI of both sexes reached their lowest values during winter where the gonads, began to increase greatly during spring, reached their highest values during summer and began to decrease again during autumn (Table 1).
| Winter | Spring | Summer | Autumn | |
|---|---|---|---|---|
| Male catfish | 1.07 | 3.6 | 4.0 | 2.2 |
| Female catfish | 9.3 | 26.4 | 30.7 | 18.8 |
Table 1: GSI values for male and female catfishes during different seasons of the year.
The tunica albuginea of both testes and ovaries had no uniform thickness all over the year but, it reached a maximum thickness during winter and became thin during spring and summer due to pressure exerted on it by the distended testicular lobules or enlarged mature follicles, and began to increase again during autumn.
Our results revealed testicular lobules of variable shapes and sizes according to seasons, where it decreased in size during winter which was cold, rainy and short day light season (resting season) and thereafter increased during spring and reached a maximum size during summer which was hot, dry and long day light season (spawning season) as they were distended by different developmental stages of spermatogenic cells and they showed slight decrease again during autumn which was dry and less hot than spawning season (spent season) but still enlarged and larger as compared to structure during winter. This finding derives support from studies of Yoakim [12] in S. schall, Latif and Salem [13] in L. nebulosus, Resink et al. [14] in C. gariepinus and [6] in Bagrus species. Spermatogonia were found throughout the year, but they were abundant during the winter season to replenish the testes after this resting season where the testes showed degeneration and increased interstitial connective tissue. This finding is in conformity to those of Zaki et al. [15] and Resink et al. [14] in C. gariepinus, Dziewulska and Domaga [16] in salmonid and Guerriero et al. [17] in L. cephalus. In this study spermatocytes were found throughout the year, but they were abundant during the spawning season. Similar observations were also reported by Rosenblum et al. [18] in I. nebulosus and Gaber [6] in Bagrus species. The spermatids increased during spring to produce spermatozoa and became few during summer as most of them were changed into spermatozoa. The spermatozoa began to appear within lumen of testicular lobules during spring while during summer, the spermatozoa increased and the testicular lobules showed different activity where some lobules were filled with spermatozoa and others were spent or empty as they discharged their content during spawning. Similar findings have also been noticed by Latif and Salem [13] in L. nebulosus, Resink et al. [14] in C. gariepinus and Guerriero et al. [17] in L. cephalus. During autumn, most of the lobules were empty indicating spawned testes, but some testes were filled with spermatozoa.
The pre-vitellogenic follicles were found throughout the year, but were common in autumn as the spent ovaries and in winter as the resting ovary and less abundant during spring and summer (spawning seasons). This result was supported by studies of Ismail [19] in C. lazera and Dougbag et al. [11], Salem [20], and Abel El Hafez et al. [21] in other fish. The vitellogenic follicles decreased in autumn as the spawning begun to end and rarely observed during winter as it was resting season but, abundant during spring and summer (spawning season) in order to turn rapidly into mature follicles. Maddoch and Burton [22] reported that the oocytes undergoing vitellogenesis indicate spawning activity. The mature or post-vitellogenic follicles were common and abundant during spring and summer as they were in the the spawning and ready to spawn and ovulate, but rarely observed during the winter (resting) season. The atretic follicles were present throughout the year but they were abundant during autumn (spent ovaries) and winter (resting ovaries) and these atretic follicles indicated the spawned individuals. This finding is similar to those obtained by Yoakim [12] in S. schall, Gaber [6] in Bagrus and Merson et al. [23] in summer flounder.
Another method of studying the seasonal variations of the gonads was the values of Gonado-somatic index (GSI) for both male and female catfish which was used as an indicator of gonadal development as when the GSI reached a maximum value, this gave a perfect indication to the time of spawning. This was supported by Ismail [19] in C. lazera; Dougbag et al. [11] in T. niloticus, Khallaf et al. [24] and Gaber [6] in Bagrus species.
The results about GSI were coincided with the obtained histological results where the GSI of both sex reached their minimum values during winter season where the gonads were in resting season, began to increase greatly during spring season, reached their maximum values during summer where the gonads were in the spawning season and began to decrease again during autumn season. This indicated a long spawning season of the catfish extending from spring to summer and might extend into autumn depending on the temperature, but its peak was reached during summer season.
The histological results were coincided with GSI as both of them revealed that winter is the resting season of catfish’s gonadal activity, spring and summer are the spawning seasons of catfish’s gonads, and autumn is post-spawning or spent season of catfish’s gonads. These information help in understanding the pattern of catfish reproduction in Egypt that may help in aquaculture of catfish.