Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 3
Grape seed extract (GSE), a nutraceutical rich in polyphenol proanthocyanidins, was evaluated for its toxicity to human oral cells. Using the neutral red cytotoxicity assay, human squamous carcinoma (HSC-2) cells were shown to be more sensitive to GSE than were gingival fibroblasts. Polyphenols undergo auto-oxidation reactions in cell culture medium, to generate reactive oxygen species, in particular, hydrogen peroxide. Using the FOX assay to quantify hydrogen peroxide, GSE, when added to cell culture medium, generated hydrogen peroxide, albeit at relatively low levels. No hydrogen peroxide was detected in the presence of catalase, which catalyzes the decomposition of hydrogen peroxide, and minor levels were detected in the presence of superoxide dismutase, which stabilizes polyphenols. Superoxide free radical, detected with nitroblue tetrazolium, was identified in GSE-amended medium. Focusing on carcinoma HSC-2 cells, the 24-hr toxicity of GSE was unaffected by the hydrogen peroxide scavengers, catalase and pyruvate, indicating that hydrogen peroxide played no role in toxicity. For HSC-2 cells cotreated with GSE and D,L-buthionine-[S,R]-sulfoximine, a depleter of intracellular glutathione, no potentiation of toxicity occurred. Over the 24 hr toxicity range, GSE did not affect the level of intracellular glutathione; however, some depletion occurred, but only at elevated GSE concentrations. GSE toxicity, apparently, was independent of oxidative stress. The 24-hr cytotoxicity of GSE to HSC-2 cells was potentiated in the presence of SOD, indicating that the proanthocyanidins per se, rather than their auto-oxidation products, accounted for toxicity. To confirm the instability of GSE in cell culture medium with a concomitant lowering of potency, studies compared medium freshly-amended with GSE to “spent” medium, i.e., GSE-amended medium left in the incubator for 24 hr prior to usage. The potency of GSE to HSC-2 cells was significantly lowered following incubated in “spent” medium for 24 hours.
<Epidemiological evidence has indicated that the dietary consumption of fruits, vegetables, and grains is a prime factor in promoting health and in reducing the risk of malignancy. Research in the chemoprevention of cancer, using cell lines derived from human tumors as the biological targets, has benefitted from the discovery that many dietary plant products, polyphenols in particular, have anticancer properties. These in vitro studies have evaluated the anticarcinogenic properties both of purified polyphenols, such as epigallocatechin gallate (EGCG), the main polyphenol in green tea, and of polyphenol extracts from specific plants, either freshly prepared extracts or commercially available extracts. The study herein evaluated the anticancer properties of a commercially available grape seed extract (GSE) towards human squamous carcinoma HSC-2 cells isolated from the oral cavity.
Oral squamous cell carcinomas, one of the most common head and neck carcinomas, exhibit high morbidity and mortality and are one of the leading malignancies in the USA [1]. Because of the adverse side effects of chemotherapeutics and because of the severity of malignancies, patients often gravitate towards complementary and alternative medicines, such as the consumption of health and dietary supplements. GSE, a nutritional supplement with known disease prevention characteristics, including anticarcinogenic, antiinflammatory, and procardioprotective effects, is rich in polyphenol compounds termed proanthocyanidins, composed primarily of dimers, oligomers, and polymers of (+)-catechin and (-)-epicatechin and their gallic acid esters [2-6].
Studies conducted in vitro have shown GSE to be selectively cytotoxic to cancer cells. At the cellular level, the biological effects of GSE included inhibition of cell proliferation, induction of cell cycle arrest, inhibition of the metastatic processes of cell migration and invasion, and induction of apoptosis. At the molecular level, some of the effects of GSE on carcinoma cells were attributed to loss of mitochondrial membrane potential, modulation of pro- and antiapoptotic signaling proteins (e.g., a decrease in the level of the antiapoptotic-promoting protein Bcl-2 and an increase in the level of the apoptotic-promoting protein Bax), inhibition of cyclins (e.g., brought about by up-regulation of p21), suppression of matrix metalloproteinases (e.g., such as MMP-2 and MMP-9), inhibition of invasive potential (e.g., brought about by a reduction in epidermal growth factor receptor (EGFR) expression), and involvement of both caspase-independent and caspase-dependent apoptotic pathways (e.g., evidenced by the upregulation of caspases-2, -3 and -8) [4,7-9].
It is well known that purified polyphenols and polyphenol extracts are unstable in cell culture medium and undergo auto oxidation reactions to generate reactive oxygen species (ROS), including hydrogen peroxide (H2O2) and, to a lesser extent, superoxide radical [10]. The extent of the prooxidant characteristic of polyphenols and polyphenol extracts differs for each compound and extract and is mediated, in part, by the chemical composition of the specific cell culture medium. The aim of the studies herein was to identify the main trigger accounting for the cytotoxicity of GSE to HSC-2 cells. The mode of toxicity of GSE could be due solely to the generation of ROS, solely to the proanthocyanidins per se, or to a combination of the generated ROS and of the proanthocyanidins.
In this study, two approaches were used to elucidate whether oxidative stress was involved in the cytotoxicity of GSE to HSC-2 cells. The most direct technique to demonstrate that polyphenol-induced cytotoxicity was correlated to the generation of ROS, specifically of H2O2, is to evaluate the cytotoxic effect in the absence and presence of catalase [11]. The complete reversal of polyphenol-induced cytotoxicity in the presence of catalase is indicative that the adverse effect was mediated entirely through the auto oxidation of the polyphenol with generation of H2O2, as noted in the induction of apoptosis in lymphoma HS-Sultan cells cotreated with EGCG and catalase [12]. Partial, but not total, reduction of polyphenol-induced cytotoxicity in the presence of catalase is indicative of a dual mode of action, toxicity due to the generated H2O2 and to the polyphenol, each triggering distinct pathways leading to cell death. Such a two-fold mode of cytotoxicity was seen in the response of human oral carcinoma cells to black tea theaflavin monogallates and catalase [13]. At times, although H2O2 may be detected in cell culture medium amended with a polyphenol, no protection occurred in the presence of catalase. In such cases, the level of generated H2O2 was not sufficient to exert a cytotoxic effect, with cytotoxicity due solely to the polyphenol itself, as seen with oral carcinoma S-G cells coexposed to catechin gallate and catalase [14].
Another research strategy to discern whether oxidative stress is a factor involved in a cytotoxic response is to modulate and to analyze the intracellular content of reduced glutathione (GSH), a ubiquitous sulfhydryl-containing tripeptide with the major roles of maintaining the intracellular redox balance and eliminating ROS toxicity. In this approach, cells are cotreated with the polyphenol extract in the presence of a GSH depleter. An increase in toxicity to cells cotreated with an extract and a GSH depleter is indicative that the test agent generated levels of ROS sufficient to adversely affect viability. For example, in the presence of a GSH depleter, the cytotoxicity of a pomegranate extract, a strong prooxidant, was potentiated, whereas that of a cranberry juice extract, a weak prooxidant, was unaffected. Related to this strategy is the monitoring of intracellular GSH levels after exposure to a polyphenol extract. Decreases in intracellular GSH, as noted in cells treated with pomegranate extract but not with cranberry juice extract, were indicative of oxidative stress [15,16].
These two approaches, cotreating the HSC-2 cells with GSE in the presence of H2O2 scavengers and in the presence of GSH depleters, as well as monitoring the intracellular status of GSH upon exposure to GSE, were employed to discern the chemical trigger evoking a cytotoxic response. Such basic information is needed prior to recommending GSE, or any other polyphenol extract, as a dietary supplement to maintain health.
Cell lines
Human squamous carcinoma HSC-2 cells derived from the oral cavity were obtained from H. Sakagami, Department of Dental Pharmacy, Meikai University School of Dentistry. Saitama, Japan; HF-1 human fibroblasts from P. Sacks, New York University School of Dentistry, New York, NY, USA; and human gingival (GT1) fibroblasts, immortalized by transfection with human telomerase reverse transcriptase, from N. Kamata, School of Dentistry, University of Tokushima, Japan [17]. Cells were grown in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA, USA) containing 110 mg/L pyruvate (cat. No. 11995-080), supplemented with 10% fetal bovine serum (FBS), 50 U/ml penicillin G, 50 μg/ml streptomycin sulfate, and 1.25 μg/ml amphotericin B, termed the growth medium, and maintained in a humidified atmosphere with 5.5% CO2 at 37°C. Cell dissociation was achieved with 0.1% trypsin-0.04% EDTA.
Chemicals
Purified grape (Vitis inifera) seed extract, Leucoselect™ (batch 12297), a gift from Indena S.A.S., France, was solubilized in dimethyl sulfoxide at a stock concentration of 100 mg/ml; fresh stock was made for each experiment. The proanthocyanidin constituents of Leucoselect™ include approximately 15% of catechin and epicatechin monomers, 80% of epicatechin 3-O-gallate monomers, dimers, trimers, tetramers, and their gallates, and 5% of pentamers, hexamers, heptamers, and their gallates [18]. Catalase (bovine liver), superoxide dismutase (SOD) (bovine liver), D,L-buthionine-[S,R]-sulfoximine (BSO) and nitroblue tetrazolium (NBT) were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
Cell proliferation assay
Individual wells of a 96-well microtiter tissue culture plate were inoculated with 0.2 ml of the growth medium containing 2 x 104cells/ well for the carcinoma cells and 1.5 x 104 cells/well for the fibroblasts. After 1 day of incubation, the growth medium was removed and replaced with exposure medium, with or without varied concentrations of GSE. Exposure medium consisted of DMEM lacking pyruvate (Cat. No. 11995-065) and amended with 10% Serum Plus (JRH Biosciences, Lenexa, KS, USA), 2% FBS, and antibiotics. In some studies, cells were coexposed to the extract in the presence of catalase (100 Units/ml), SOD (25 and 50 Units/ml), pyruvate (110 mg/L), or 5 mM (BSO). In all such experiments, the GSE was freshly prepared in exposure medium. Some studies used “spent” GSE, i.e., 200 μg/ml GSE in exposure medium incubated at 37°C for 24 hr prior to use. One complete row, i.e., 8 wells, was used for each concentration of test agent, with and without the accompanying variable. After exposure to the test agents for 1 day, viability was assessed with the neutral red (NR) assay, which is based on the uptake and lysosomal accumulation of the supravital dye, NR, as described elsewhere [14,19,20].
Intracellular glutathione assay
Measurements of intracellular reduced glutathione (GSH) were performed according to the procedures of Weisburg et al. [19]. Cells, maintained in growth medium and grown to about 80% confluence in 60 mm tissue culture plates, were washed with phosphate buffered saline (PBS) and treated for 4 hr in PBS containing 0, 50, 100, 150, and 200 μg/ml GSE. Three plates of cells were used per concentration of extract. Thereafter, the cells were washed with PBS, lysed with 0.06 ml of 0.2% Triton X-100, and proteins were precipitated with 0.06 ml of 5.0% sulfosalicyclic acid. After cell harvesting by scraping and centrifugation at 12,000xg for 5 min, GSH was determined in 0.1 ml aliquots of the acid-soluble extract by determining the oxidation of GSH by a 6 mM solution of 5,5′-dithiobis (2-nitrobenzoic acid), prepared in a phosphate buffer/EDTA (pH 7.5), to glutathione disulfide (GSSG), with a stoichiometric formation of 5-thio-2-nitrobenzoic acid, a yellow chromagen measured spectrophotometrically at 412 nm. With each assay, a standard curve was generated with known amounts of GSH and the data were recorded as GSH, in nmoles/106 cells/ml.
H2O2 assay
Measurement of H2O2 was carried out by the ferrous ion oxidation xylenol orange (FOX) method using the PeroXOquant Quantitative Peroxide Assay Kit, lipid-soluble formulation (Pierce Biotechnology, Rockford, IL, USA). The principle of the FOX assay is based on H2O2 oxidation of Fe2+ to Fe3+, which reacts with xylenol orange to yield a xylenol orange-Fe3+ complex measured spectrophotometrically. The protocol was followed as suggested by the manufacturer, but with some modifications. Briefly, after a 2- or 4- hr incubation at room temperature, 90 μl samples of exposure medium, unamended DMEM, or of PBS, adjusted to pH 5.4, 6.4, and 7.4, and amended with GSE were mixed with 10 μl of methanol and incubated at room temperature for 20 min. In other studies, GSE in the exposure medium was coincubated for 4 hr with 200 Units/ml catalase or SOD. After the appropriate time of incubation, FOX reagent (0.9 ml) was added, followed by vortexing and 20 min incubation at room temperature. The solutions were centrifuged at 12,000 x g for 5 min and absorbance of the supernatant was read at 595 nm against a blank consisting of 90 μl of exposure medium or PBS, 10 μl of methanol, and 0.9 ml of FOX reagent. A standard curve of H2O2 from 0 to 200 μmol/L was used to quantify the level of hydrogen peroxide generated by GSE [20].
Superoxide assay
The generation of superoxide by GSE, performed according to the method of Yen et al. [21], is based on reduction of yellow-colored NBT by superoxide to a blue formazan product. One ml of a 50 μM phosphate-100 μM EDTA buffer (pH 7.4), containing 100 μM NBT, 200 μg/ml GSE, and without or with 200 Units/ml SOD were combined in a cuvette and absorbance was measured every 10 min at 560 nm. A decrease in absorbance in the presence of SOD, as compared to the absence of SOD, indicated generation of superoxide by GSE.
Microscopy
HSC-2 cells, grown on coverslips in 35 mm cell culture plates, were exposed for 24 hr to fresh GSE in the absence and presence of SOD and to “spent” GSE. Thereafter the cells were fixed in methanol, stained with aceto-orcein, and subsequently observed with bright field microscopy.
Statistics
All experiments were performed a minimum of three times. Data for the FOX assay were presented as the arithmetic mean ± standard errors of the mean (S.E.M.) and for the cytotoxicity assay data were presented as the mean arithmetic percentages of control ± S.E.M. Experimental data were analyzed with a one-way analysis of variance (ANOVA) followed by Tukey’s multiple range test for significant differences. The P-value of the effect had to be ≤ 0.05 to be considered significant.
Cell-free systems
Purified polyphenols (e.g., black tea theaflavin-3,3′-digallate [20]) and complex polyphenol plant (e.g., apple [22]) extracts amended in cell culture medium are unstable and undergo auto oxidation reactions to generate various ROS, principally, H2O2. The generation of H2O2 in cell culture exposure medium amended with GSE was both concentration- and time-dependent. Progressively increasing the GSE concentration from 50 to 200 μg/ml and increasing the incubation time from 2 to 4 hr resulted in concomitant increases in H2O2 (Figure 1). Roques et al. [23], in studying the autoxidation of GSE and of purified polyphenols in DMEM, concluded that the degradation reactions were triggered primarily by the inorganic salts mixture in the cell culture medium, with lesser effects by other cell culture medium components, such as FBS, vitamins, and antibiotics.
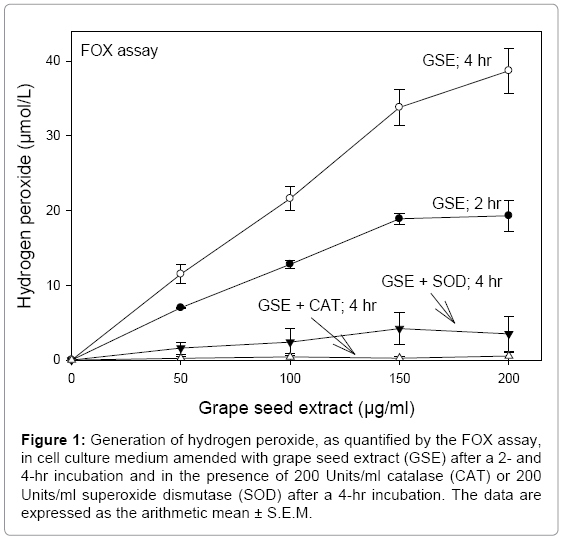
Figure 1: Generation of hydrogen peroxide, as quantified by the FOX assay, in cell culture medium amended with grape seed extract (GSE) after a 2- and 4-hr incubation and in the presence of 200 Units/ml catalase (CAT) or 200 Units/ml superoxide dismutase (SOD) after a 4-hr incubation. The data are expressed as the arithmetic mean ± S.E.M.
To obtain a better understanding of the magnitude of H2O2 generated, for comparative purposes, the prooxidant nature of GSE was compared to that of a pomegranate ellagitannin-enriched polyphenol extract [15] and of a cranberry juice extract, rich in proanthocyanidins [16], after a 2-hr incubation at concentrations up to 200 μg/ml in exposure medium. At a concentration of 200 μg/ml of extract, pomegranate extract generated levels of H2O2 about 7 times greater than that for GSE and about 8½ times greater than for cranberry juice extract (Figure 2). Thus, as compared to the pomegranate extract, the GSE extract and cranberry juice extract are weak proooxidants.
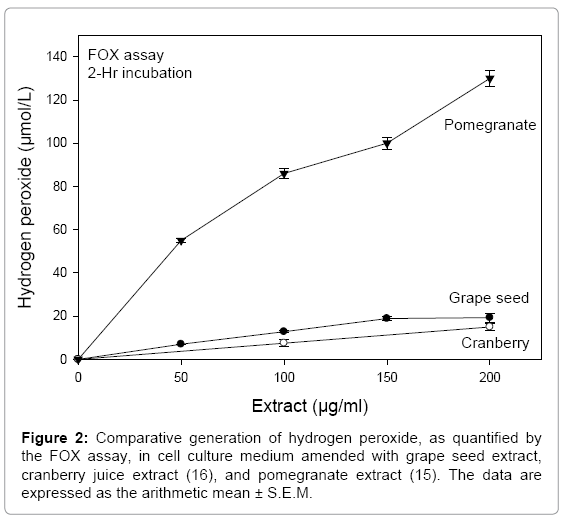
Figure 2: Comparative generation of hydrogen peroxide, as quantified by the FOX assay, in cell culture medium amended with grape seed extract, cranberry juice extract (16), and pomegranate extract (15). The data are expressed as the arithmetic mean ± S.E.M.
Incubation of 200 μg/ml GSE for 4 hr in PBS, maintained either at pH 5.4, 6.4, or 7.4, did not lead to detectable levels of H2O2. Prior studies with Gingko biloba leaf extract showed that this extract was relatively stable at pH 5.4 and 6.4, with some H2O2 detected at pH 7.4 [24]. Comparatively, GSE is a very poor generator of H2O2, which probably accounted for the inability to detect H2O2 in GSE-amended PBS at pH 7.4. A 4-hr incubation of 200 μg/ml GSE in unamended DMEM (i.e., DMEM without Serum Plus or FBS), yielded a level of 78 ± 6.5 μmol/L H2O2, whereas 200 μg/ml GSE in exposure medium (DMEM+10% Serum Plus and 2% FBS), yielded 38 μmol/L H2O2. The higher level of H2O2 detected in unamended DMEM versus that detected in exposure medium likely reflected scavenging of the H2O2 by the additional proteins in the exposure medium.
After a 4-hr incubation of GSE in exposure medium, the amount of H2O2 was almost completely eliminated in the presence of 200 Units/ ml catalase and was lowered by greater than 90% in the presence of 200 Units/ml SOD (Figure 1). Catalase enzymatically degrades the H2O2 (H2O2 → 2H2O+O2), thereby accounting for the inability to detect H2O2 in medium coamended with GSE and catalase. As catalase is specific for H2O2, these studies also indicated that H2O2 was the prime ROS generated by GSE in cell culture medium. Greatly lowered levels of H2O2 were detected in exposure medium coamended with GSE and SOD. Similar findings were noted for mixtures of SOD and epigallocatechin gallate [12,25], quercetin, an apple polyphenol extract [22], and cranberry juice extract [16] in cell culture medium and of SOD and myricetin [26], EGCG [27], and theaflavin-3,3′-digallate [20] in inorganic buffers. Hou et al. [25] demonstrated that the lesser levels of H2O2 detected in cell culture medium coamended with EGCG and SOD reflected stabilization of the polyphenol by SOD, which prevented its auto oxidation.
Auto oxidation of GSE generated, in addition to H2O2, the superoxide radical (Figure 3). Tyagi et al. [28] suggested that superoxide radical, rather than H2O2, was the main ROS produced by the auto oxidation of GSE. However, as demonstrated by the FOX assay, the elimination of ROS in exposure medium coamended with GSE and catalase clearly indicated the generation of H2O2. The slightly higher level of H2O2 detected by the FOX assay in medium coamended with GSE+SOD, as compared to medium coamended with GSE+catalase (Figure 1), also was observed in studies with cranberry juice extract [16] and theaflavin-3,3’-digallate [20]. The detection of ROS in media amended with these polyphenols and SOD indicated some minor degree of auto oxidation, leading to the generation either of H2O2 directly or to low levels of superoxide radical, which subsequently were catalyzed by SOD to H2O2.
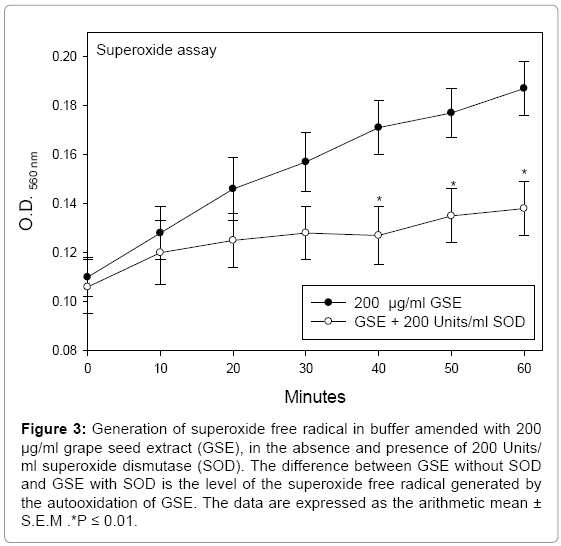
Figure 3: Generation of superoxide free radical in buffer amended with 200 μg/ml grape seed extract (GSE), in the absence and presence of 200 Units/ ml superoxide dismutase (SOD). The difference between GSE without SOD and GSE with SOD is the level of the superoxide free radical generated by the autooxidation of GSE. The data are expressed as the arithmetic mean ± S.E.M .*P ≤ 0.01.
Cellular systems
The comparative sensitivities of oral carcinoma HSC-2 cells, normal gingival HF-1 fibroblasts, and transfected gingival GT1 fibroblasts were studied after a 24-hr exposure to GSE, with cytotoxicity quantified by the NR cytotoxicity assay. For the HSC-2 cells, GSE decreased cell viability in a concentration-dependent manner, with toxicity noted initially (P ≤ 0.01) at 75 μg/ml GSE and with a midpoint cytotoxicity (ED50) value of 85 μg/ml GSE. Over the concentration range of 25 to 150 μg/ml GSE, toxicity was not noted towards the GT1 and HF-1 fibroblasts. Conversely, from 50 to 150 μg/ml GSE the HF-1 fibroblasts exhibited growth stimulation (Figure 4) Ye et al. [29] noted that GSE was cytotoxic to human gastric adenocarcinoma and chronic myelogenous leukemic cells, but stimulatory to normal human gastric mucosal cells and to normal murine macrophages. Shirataki et al. [30] evaluated the relative cytotoxicities of different fractions of grape seed extract to HSC-2 carcinoma cells and gingival fibroblasts. Two of the 4 GSE fractions were cytotoxic to the HSC-2 carcinoma cells (ED50 of 146 and 154 μg/ml GSE), but were without toxicity to the gingival fibroblasts, even at 500 μg/ml GSE. Chatelain et al. [7] noted that at concentrations from 30 to 80 μg/ml, GSE was toxic to human tongue CAL27 carcinoma cells but was without effect to human fibroblasts. Shrotriya et al. [31] observed that head and neck squamous cell carcinoma Detroit 562 and FaDu cells were highly sensitive to 20 and 40 μg/ml GSE after a 24 to 48 hr treatment, but similar treatments to normal human epithelial keratinocytes were without a toxic effect. The lesser sensitivities of normal fibroblasts, as compared to carcinoma cells, to polyphenols, both as purified chemicals and as components of plant extracts, are well documented [32]. As the HF-1 and GT1 fibroblasts were resistant to GSE, further studies to evaluate the mode of toxicity of GSE were directed solely to the HSC-2 carcinoma cells.
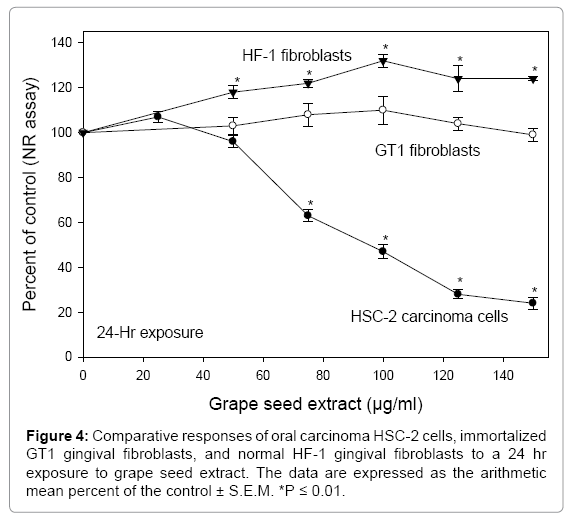
Figure 4: Comparative responses of oral carcinoma HSC-2 cells, immortalized GT1 gingival fibroblasts, and normal HF-1 gingival fibroblasts to a 24 hr exposure to grape seed extract. The data are expressed as the arithmetic mean percent of the control ± S.E.M. *P ≤ 0.01.
Both catalase and pyruvate scavenge H2O2; catalase enzymatically degrades H2O2 and pyruvate undergoes an oxidative decarboxylation reaction with H2O2 (pyruvate+H2O2 → acetate+CO2+H2O) [11]. After a 24-hr exposure of the HSC-2 cells to 75 and 100 μg/ml GSE, cell viability values, as determined by the NR assay, were 63 ± 2.8 and 47 ± 3.2%, respectively, of the control (nontreated with GSE). Upon cotreatment with GSE and 100 Units/ml catalase, the respective cell viability values were 72 ± 2.8 and 48 ± 4.3% and for a cotreatment of GSE with pyruvate (110 mg/L), the respective cell viability values were 71 ± 7.6 and 46 ± 3.8%. The lack of protection against GSE toxicity in the presence of these scavengers of H2O2 indicated that the level of H2O2 generated by the auto oxidation of GSE in cell culture medium was insufficient to evoke a cytotoxic response. These findings were confirmed by the relatively low prooxidant nature of GSE, compared to pomegranate extract, as noted in the FOX assay (Figure 2). Prior studies with pomegranate extract showed that H2O2 was a significant component in its overall cytotoxicity to HSC-2 cells, as the toxicity of pomegranate extract was lessened in the presence of catalase and pyruvate [15]. Conversely, cranberry juice extract, an extract with weak prooxidant activity similar to that of GSE (Figure 2), showed no involvement of H2O2 in its cytotoxic potential, as toxicity was unaffected by cotreatment with catalase and pyruvate [16].
The other research approach to discern whether oxidative stress was a factor in the cytotoxic response is to evaluate the prooxidant nature of GSE by modulating the intracellular level of GSH in the target cells. GSH is enzymatically synthesized in a two-step process, involving γ-glutamylcysteine synthetase and glutathione synthetase. BSO, as a selective inhibitor of γ-glutamylcysteine synthetase, prevents the recycling of GSH from GSSG, thereby enhancing cell susceptibility to oxidative stress. Viability of HSC-2 cells was unaffected by cotreatment with 5mM BSO and 75 and 100 μg/ml GSE; in the presence of BSO, the respective values for cell viability were 98 ± 3.6 and 63 ± 4.0%, i.e., values equivalent to GSE alone. For polyphenols that are strong prooxidants, treatment of the target cells with GSH depleters potentiated their toxicities, as was noted for a green tea polyphenol extract [14], G. biloba leaf extract [24], theaflavin-3,3′-digallate [20], and EGCG [19].
To further evaluate GSE as an agent of oxidative stress, the level of intracellular GSH was monitored in HSC-2 cells incubated for 4 hr in the presence of 50 to 200 μg/ml GSE. A significant decrease of intracellular GSH occurred only at 150 (P ≤ 0.05) and 200 (P ≤ 0.01) μg/ml GSE (Figure 5). However, to achieve these decreases in intracellular GSH, the concentrations of GSE greatly exceeded those for a 24-hr cell exposure and thus, because of their lethality, these elevated concentrations were not of physiological relevance. Shao et al. [33] noted that GSE induced oxidative stress to chick cardiomyocytes only at the elevated level of 500 μg/ml with toxicity reduced by the antioxidant, N-acetylcysteine.
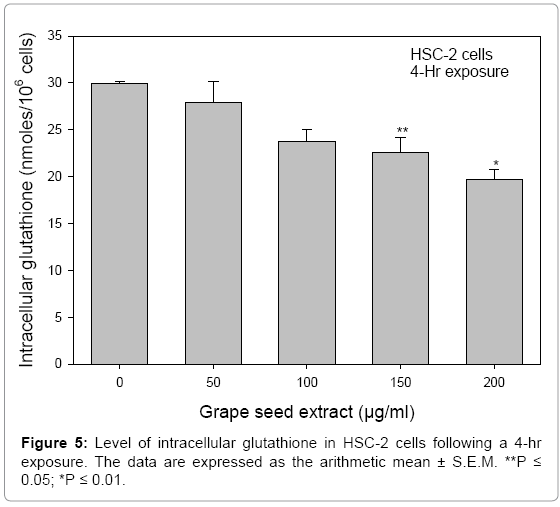
Figure 5: Level of intracellular glutathione in HSC-2 cells following a 4-hr exposure. The data are expressed as the arithmetic mean ± S.E.M. **P ≤ 0.05; *P ≤ 0.01.
In contrast to the studies herein, in which GSE was a weak prooxidant with toxicity independent of oxidative stress, others have reported that moderate concentrations of GSE generated ROS at levels high enough to induce toxicity. Shrotriya et al. [31] demonstrated that GSE-induced growth inhibition, DNA damage, and apoptosis to head and neck squamous carcinoma cells were reversed in the presence of the N-acetylcysteine. The commercially available GSE extract used was described as highly rich in oligomeric proanthocyanidins and other valuable flavonids. Using the same GSE extract as noted above, Raina et al. [34] showed that N-acetylcysteine ameliorated GSEinduced apoptosis of bladder cancer cells and Tyagi et al. [28] observed reductions in intracellular GSH in lung cancer cells exposed to GSE.
These three studies showing GSE as an inducer of oxidative stress used GSE from a common source which, apparently, had different properties than the GSE used in the studies reported herein. This issue was recognized by those researchers, as they noted that the lack of availability of uniformly standardized GSE extracts raised questions whether different GSE samples would have comparable biological activities and would trigger similar molecular mechanisms. They speculated that GSE from different cultivars may evoke distinct signaling pathways, although each may exert selective cytotoxicity to cancer cells [35]. The prooxidant nature of the GSE extract as the cause of toxicity noted in these other studies was not evident in the research herein, albeit both extracts were selectively cytotoxic to carcinoma cells.
It is important to note that polyphenols have interesting biochemical properties, as, at low concentrations they may act as antioxidants (and thereby have anticancer properties) and at high concentrations act as prooxidants (and also exert anticancer effects) [36]. GSE is commonly marketed in the USA and Europe for its strong antioxidant properties. As an antioxidant, GSE demonstrated greater free radical scavenging activities and provided greater protection against free radical-induced lipid peroxidation and DNA damage than vitamins C, E, and β-carotene [2]. Acting as an antioxidant, GSE protected human lens epithelial cells from H2O2-induced oxidative stress [37], human lymphoblastoid cells from H2O2-induced chromosomal damage [5], primary rat glial cells from nitrosative/oxidative stress [38], and normal human oral keratinocytes from oxidative stress induced by a smokeless tobacco extract [39].
Supplementing cell culture medium with 25 and 50 Units/ml SOD greatly enhanced the cytotoxicity of 25 and 50 μg/ml GSE to the HSC-2 cells (Figures 6, Figure 7a-d). SOD was shown to potentiate the cytotoxicity of cranberry juice extract to HSC-2 cells [16] and of EGCG to mouse osteoclasts [27] and to human esophageal cancer KYSE 150 cells [25]. Hou et al. [25] showed that SOD stabilized and abrogated the auto oxidation of EGCG, thereby lessening the generation of H2O2, yet potentiated the antiproliferative effects of EGCG. From the studies of Hou et al. [25] and Nakagawa et al. [27] with EGCG and from prior studies with cranberry juice extract [16], it would appear that SOD-stabilized polyphenols exerted greater cytotoxicities than their autooxidative end products.
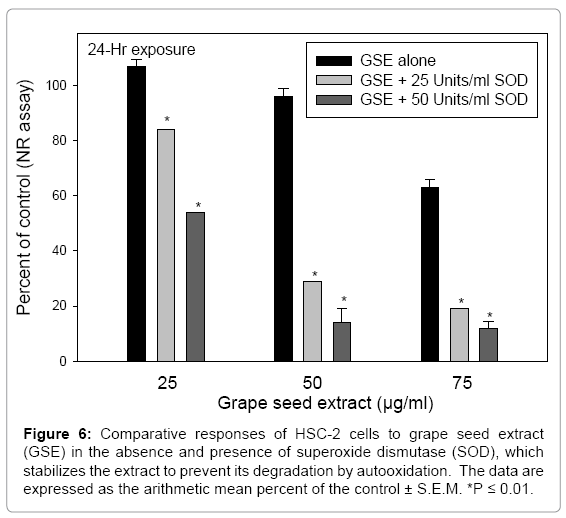
Figure 6: Comparative responses of HSC-2 cells to grape seed extract (GSE) in the absence and presence of superoxide dismutase (SOD), which stabilizes the extract to prevent its degradation by autooxidation. The data are expressed as the arithmetic mean percent of the control ± S.E.M. *P ≤ 0.01.
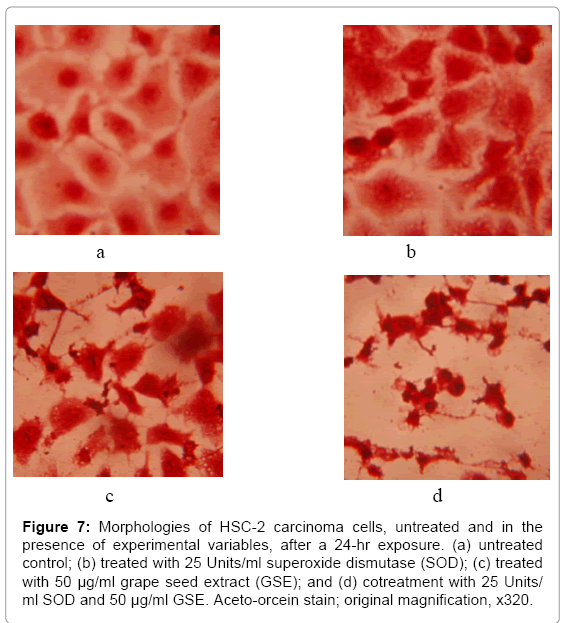
Figure 7: Morphologies of HSC-2 carcinoma cells, untreated and in the presence of experimental variables, after a 24-hr exposure. (a) untreated control; (b) treated with 25 Units/ml superoxide dismutase (SOD); (c) treated with 50 μg/ml grape seed extract (GSE); and (d) cotreatment with 25 Units/ ml SOD and 50 μg/ml GSE. Aceto-orcein stain; original magnification, x320.
The instability of GSE in cell culture medium, as related to its potency, was shown by contrasting the cytotoxicity of GSE freshly prepared in cell culture medium with GSE prepared at a stock concentration of 200 μg/ml in cell culture medium, incubated for 24 hr at 37°C prior to use (termed, the “spent” medium). After 24 hr, the “spent” medium stock had a concentration of 145 ± 8.9 μmol/L H2O2. Roques et al. [23], using an in-house preparation of GSE, measured 138 μmol/L H2O2 in DMEM amended with 400 μg/ml GSE after 24 hr incubation at 37°C. The “spent” medium with 200 μg/ml GSE was diluted to yield concentrations of 75, 100, and 125 μg/ml GSE, yielding 47 ± 3.2, 65 ± 3.8, and 85 ± 5.6 μmol/L H2O2, respectively. Comparisons of GSE prepared in fresh medium with those prepared from “spent” medium demonstrated the lesser potency of GSE prepared from “spent” stock incubated overnight (Figure 8). The instability of GSE in medium incubated for 24 hr at 37°C, apparently, lead to degradation products which exerted lesser potencies than of freshly prepared GSE.
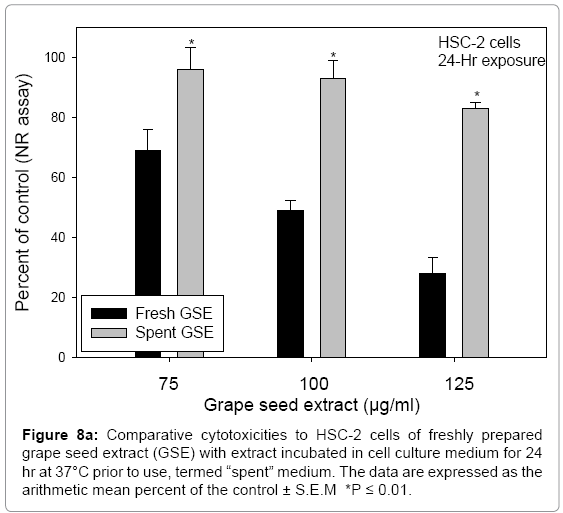
Figure 8a: Comparative cytotoxicities to HSC-2 cells of freshly prepared grape seed extract (GSE) with extract incubated in cell culture medium for 24 hr at 37°C prior to use, termed “spent” medium. The data are expressed as the arithmetic mean percent of the control ± S.E.M *P ≤ 0.01.
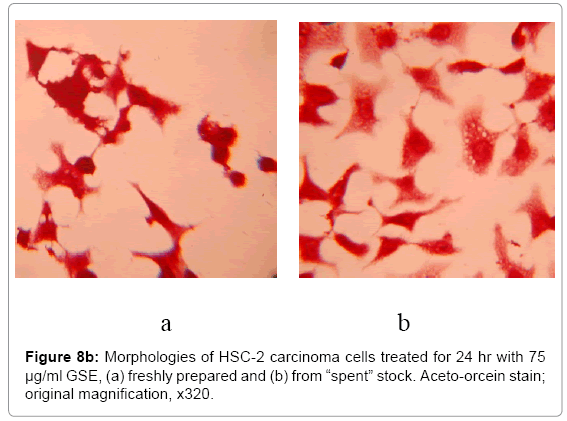
Figure 8b Morphologies of HSC-2 carcinoma cells treated for 24 hr with 75 μg/ml GSE, (a) freshly prepared and (b) from “spent” stock. Aceto-orcein stain; original magnification, x320.
The GSE extract used herein, Leucoselect™, exerted selective cytotoxicity to carcinoma HSC-2 cells via a mechanism independent of the induction of oxidative stress. The GSE concentration range toxic to HSC-2 cells was without an adverse effect to gingival fibroblasts. The lack of amelioration of GSE toxicity to the carcinoma cells upon coexposures with catalase or pyruvate, both scavengers of H2O2, and the lack of potentiation of GSE toxicity upon cotreatment with BSO, a GSH depleter, showed that although GSE generated ROS, the levels were not significant enough to induce oxidative stress. This was confirmed by the slight depletion of intracellular GSH only at elevated GSE concentrations (i.e., exceedingly those of the toxicity concentration range for a 24-hr exposure). These finding are in opposition to those of others, who noted GSE toxicity to be linked to its proooxidant nature. In the studies herein, as further noted with coexposures to GSE and SOD and with GSE prepared from “spent” medium, the cytotoxicity of GSE apparently was related to the constituent intact proanthocyanidins, rather than to their auto oxidation end products.
As prevention of cancer via dietary consumption of natural products has much potential, a clear understanding of the mechanisms that trigger their cytotoxic responses is warranted. For GSE, without standardization, although different extracts may show anticancer potentials, their modes of toxic action vary greatly - from toxicity due to their proanthocyanidins per se (as shown herein) to toxicity from their prooxidant nature arising from decomposition, autoxidation reactions. Coupled with this divergence of toxic mechanisms is that most GSE supplements are manufactured and consumed for their antioxidant nature. Yet, such plant-derived polyphenol nutraceuticals are potentially very important in regard to cancer therapies, especially since a commonality among most studies is that these extracts are selectively toxic to cancer cells, without an adverse effect on normal cells. As noted herein, the GSE concentration range was toxic to oral carcinoma HSC-2 cells but not to gingival fibroblasts. Nutraceuticals, such as GSE, that are selectively cytotoxic to cancer cells, may be important adjuncts to chemotherapeutics administered in the treatment of cancer [40].
Appreciation is expressed to the S. Daniel Abraham Honors Program and to the JFEW Fellowship Program of Stern College for Women, Yeshiva University, for support of this research.