Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2013) Volume 4, Issue 1
The ionic contents and osmolality of seminal plasma of beluga spermatozoa to determine and their physiological relationship with sperm motility characteristics (total duration of sperm motility and percentage of motile cell) were studied. The results obtained show that Na+ (82.54 ± 5.46 mM L-1), Cl- (15.95 ± 0.72 mM L-1) and K+ (3.57 ± 0.15 mM L-1) were predominant ions in the seminal plasma. The osmolality of seminal plasma ranged from 57 to 93 mOsm kg-1. Significant positive correlations were observed between osmolality vs. Na+ (p<0.01) and Cl- (P<0.05) and Na+ vs. Cl- (P<0.05). No significant correlations were found between sperm motility and others composition of the seminal plasma. Probably, the Na+ and Cl- are the main electrolytes playing a key role in maintaining the osmolality of the seminal plasma and the viability of the spermatozoa in vivo. However analysis of ionic contents of the seminal plasma revealed some species-specific characteristics especially for K+ which is important because of its inhibitory effect on sperm motility. Results of the present study provide new data on beluga spermatozoa quality and can be utilized for successful reproduction management and cryopreservation protocols of this endangered species.
Keywords: Ionic content; Osmolality; Seminal plasma; Sperm motility; Beluga
Sturgeon are currently generating interest, as most belong to endangered or critically endangered species and are listed in The International Union of Conservation of Nature (IUCN) Red List of Threatened Species mostly due to overfishing for caviar and meat, poaching, habitat destruction, and late maturation and reproduction [1]. The beluga, Huso huso, belongs to the largest family of fresh water chondrostean fish, the Acipenseridae (containing 27 species), which appeared about 200 million years ago [2,3]. This fish inhabit the northern hemisphere [2] and is one of the sturgeon species with high growth rates and appears to be very suitable for aquaculture [4].
In order to evaluate the quality of semen in fish, study on sperm physiology in the testis, in the genital tract and after collection and fertilization is necessary [5]. The quality of sperm is usually assessed by the acquisition of and capacity for motility, the metabolism and respiration rate before and during motility, the biochemistry of sperm and seminal plasma, the capacity for short-time storage and the resistance to cryopreservation and the fertilizing ability of sperm [5-9]. The percentage of motile sperm cells [10] and total period of sperm motility [11] are valid parameters that are used by specialists in sperm physiology.
Seminal plasma has a unique composition: some components support the spermatozoa, while others reflect the functions of the reproductive system and the spermatozoa [12]. Sturgeon spermatozoa, like those of teleostean fish, are immotile in the seminal plasma or the sperm duct [13,14]. As in salmonidae, K+ ion is the main inhibitory factor preventing sperm activation in the seminal plasma [13,15].
Several papers have been published on spermatozoa motility and its physiological correlation with composition of seminal plasma in sturgeon and teleost species: Siberian sturgeon, Acipenser baerii [13,16], Shovelnose sturgeon, Scaphirhynchus platorynchus and Paddlefish, Polydon spathula [17], Lake sturgeon, Acipenser fluvescens [15] and Persian sturgeon, Acipenser persicus [18], Atlantic salmon, Salmo salar [19], Common carp, Cyprinus carpio [20], Bleak, Alburnus alburnus and Rainbow trout, Oncorhynchus mykiss [21,22], Ctenopharyngodon idella [23], Chinook salmon, Oncorhynchus tshawytscha [24] and Salmo trutta macrostigma [25].
The aim of the present study is to determine some parameters of the seminal plasma of the beluga, including ionic contents and osmolality and to try establishing if there is any correlation with sperm motility characteristics (percentage of motile sperm and total duration of sperm movement).
Broodfish and sampling
The experiments were carried out during March-April 2011. Male H. huso broodstocks (30-45 kg, 1-2 m) were captured from the Caspian Sea (The temperature of water was 10-11.5°C). After the transportation to a hatchery by boat (6 hrs duration), they were kept in broodstock pond at Rajaei Sturgeon Hatchery Center (Sari, Mazandaran, Iran; Lat 36°37 N, Long 53°05 E). The semen was collected from teen males by hand stripping 14-18 h after inductions of spermiation by intramuscular injection of LHRH-A2 (D-Ala6 GnRH Pro9-Net, Ningbo Hormone Product Co., China) hormone (5 μg kg-1). Contamination with water or urine was carefully prevented. Semen was stored in microtubes on ice (+4°C) during transportation to a laboratory (upto 3 h). The water temperature, oxygen content and pH were at 15-16°C, >5 mg l-1 and 7.6-7.9, respectively during experiments.
Preparation and determination of seminal plasma composition
Semen samples were centrifuged (Spectrafuge16M, USA) using a Two-step method, first at 500 rpm for 2 min, and second at 3000 rpm for 10 min. The supernatants were collected and stored as frozen in -20°C until use for analysis. The osmolality of seminal plasma was measured in duplicates per sample with osmometer (Osmomat 030-m, Germany) using a freezing point depression. To determine the ionic concentration of seminal plasma, the samples were transferred to the Medicine Laboratory of the Alavi Hospital, Ardebil, Iran. Magnesium, chloride and calcium were measured with colorimetric method using an Autoanalyser BT 3500 plus (Biotecnica, Italy). Potassium and sodium were determined with a flame photometer (SEAC, FP20, Italy).
Evaluation of sperm motility indices
Sperm motility was evaluated visually for the percentage of motile sperm after activation (%) and total period of sperm motility (sec), i.e., when almost all of the spermatozoa (near to 100%) were immotile. For this purpose a 10 μl aliquot of semen was pipetted onto a 1% (w/v) BSA-coated microscope slide. Activation was achieved by adding 25 μl of 0.3% NaCl solution. Motility observation was carried out under a pre-focused inverted microscope (400X, Olympus CK2, Tokyo, Japan) at 10°C. The percentage motility was determined arbitrarily on a 0 to 10 point scale, with 0 denoting 0% motility and 10 denoting 100% motility.
Statistical analysis
Results are presented as means ± SEM. Statistical analyses were performed with the SPSS 16 for Windows statistical software package. Correlations between seminal plasma parameters (ionic and osmolality) were estimated using Pearson correlation test. Linear and non-linear regression models were investigated using regression fits. The total duration of sperm movement and the percentage of motile sperm were used as dependent and seminal plasma parameters as independent variables.
The ionic composition and the osmolality of the seminal plasma of H. huso are shown in Table 1. The Na+, Cl- and K+ ions predominate in H. huso seminal plasma with values of 82.54 ± 5.46, 15.95 ± 0.72and 3.57 ± 0.15 mM L-1 (mean ± SEM) respectively. The seminal plasma osmolality was 73.16 ± 4.01 mOsm kg-1 (mean ± SEM). The Pearson correlation between the seminal plasma parameters are shown in Table 2. A significant positive correlations between seminal plasma parameters were found for osmolality vs. Na+ (r2=0.856, P<0.01), for Cl- vs. osmolality (r2=0.681, P<0.05), and for Na+ vs. Cl- (r2=0.580, P<0.05). In contrast, there was no significant correlation between other parameters (Table 2, P>0.05).
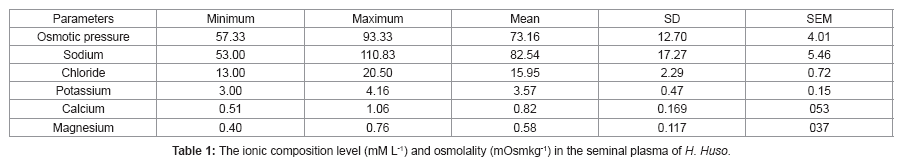
Table 1: The ionic composition level (mM L-1) and osmolality (mOsmkg-1) in the seminal plasma of H. Huso.
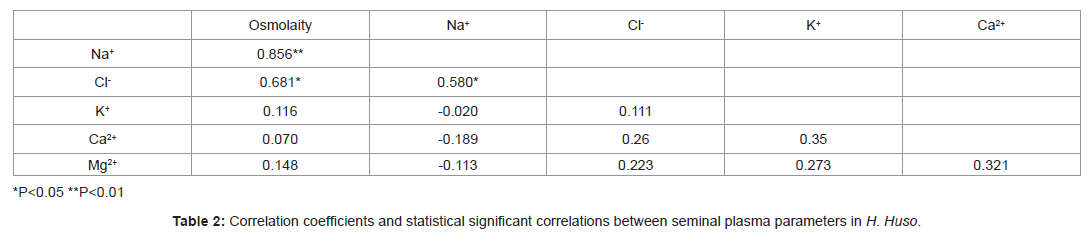
Table 2: Correlation coefficients and statistical significant correlations between seminal plasma parameters in H. Huso.
Statistically correlations and their regression functions are shown in Figure 1, where seminal plasma parameters (including ionic contents and osmolality) and sperm motility are compared. An increase in Na+ ion concentration in seminal plasma leads to increase in the total duration of sperm motility (Figure 1a). In contrast, the percentage of motile sperm cell was approximately constant (Figure 1a). When increasing Cl- content in the seminal plasma, the total duration of motility and the percentage of motile spermatozoa increased and decreased respectively (Figure 1d). Also, with increasing K+ concentration in the seminal plasma, the total duration of motility decreased but the percentage of motile sperm was approximately constant (Figure 1b). Maximum total duration of spermatozoa movement was observed when the Ca2+ concentration was about 0.6 mM L-1 in the seminal plasma. However, the percentage of motile sperm cells was lowest at 0.8 mM L-1 (Figure 1c). Minimum total duration of sperm motility was observed when the Mg2+ content was about 0.6 mM L-1 (Figure 1e). Besides, the percentage of motile cells was constant when the Mg2+ level increased (Figure 1e). Neither the percentage of motile cells nor the total duration of sperm motility decreased when the osmolality of seminal plasma was increased above 80 mOsm kg-1 (Figure 1f).
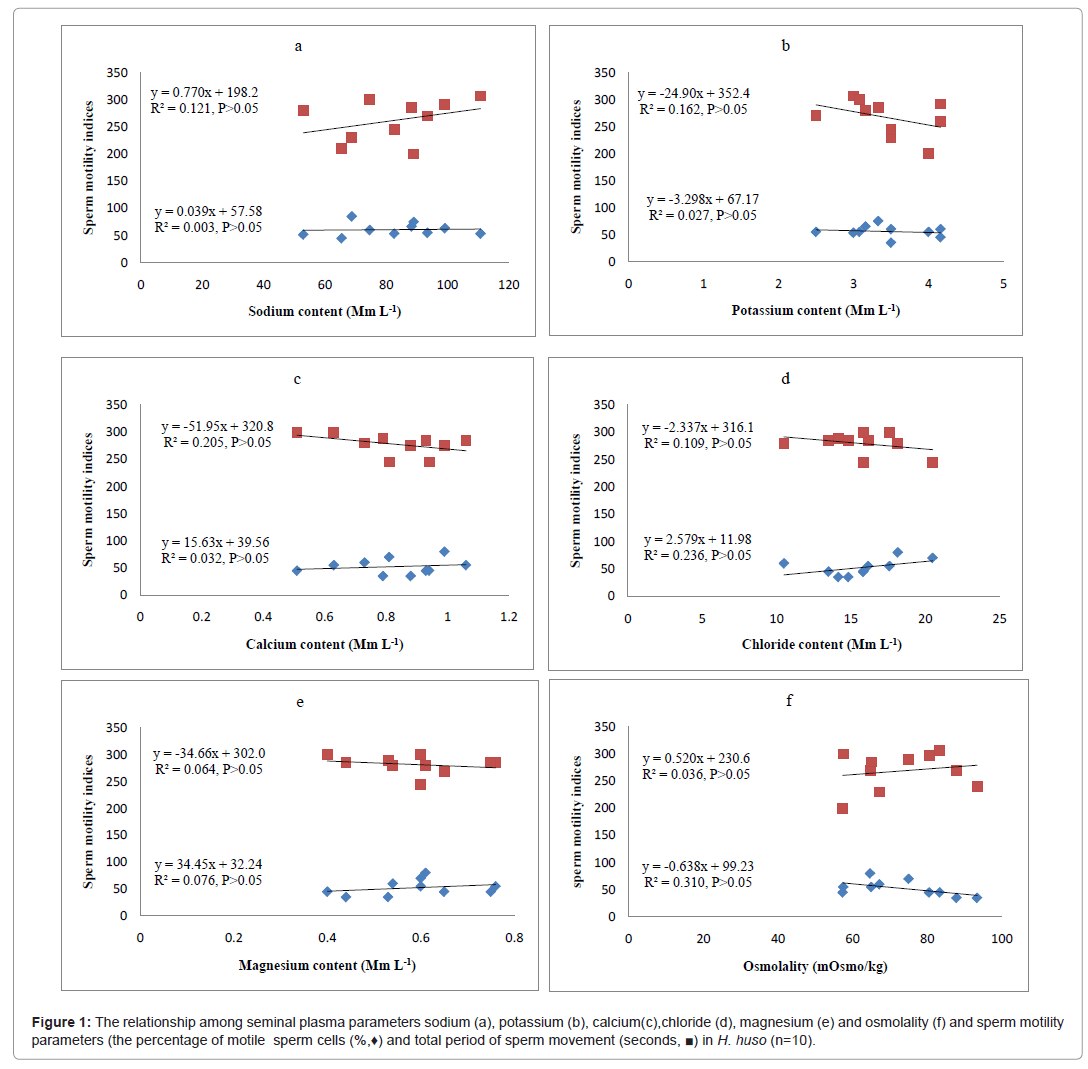
Figure 1: The relationship among seminal plasma parameters sodium (a), potassium (b), calcium(c),chloride (d), magnesium (e) and osmolality (f) and sperm motility parameters (the percentage of motile sperm cells (%,♦) and total period of sperm movement (seconds, ¦) in H. huso (n=10).
Studies on seminal plasma composition and osmolality provide information to understand the regulating mechanism affecting sperm motility and behaviour [12,26,27]. Like in teleost species, the Na+, K+ and Cl- ions predominate in sturgeon seminal plasma but their concentrations are lower than the concentrations observed in teleost seminal plasma. The ionic composition of the seminal plasma is in the range of salmonids and cyprinids reported by Billard et al., Morisawa et al. and Kruger et al. [5,11,20]: Na+: 130-140 and 94-107 mM L-1, K+: 20-66 and 39-78 mM L-1, Cl-: 135 and 98-110 mM L-1, Ca2+: 0.3-2.6 and 0.7-10.69 mM L-1and Mg2+: 0.8-3.6 and 0.02-0.8 mM L-1 respectively. Seminal plasma of H. huso has a higher Na+ content than Acipenser gueldenstaedtii (34.9 mM L-1); Acipenserstellatus (20.1 mM L-1) [28] and A. persicus (62.4 mM L-1) [18]. The K+ and Cl- levels in H. huso seminal plasma was lower than that in A. persicus (21.1 mM L-1) [18] but it was higher than those reported for Acipenser ruthenus (7.6 mM L-1) [29]; A. gueldenstaedtii and A. stellatus (13.5 and 7.5 mM L-1, respectively) [28]. Similarly, the Mg2+ and Ca2+ contents of the seminal plasma of the H. huso were higher than that in A. ruthenus (0.2 mM L-1), but their contents were close to that reported by Alavi et al. [18] in A. persicus (0.5 mM L-1). The osmolality of seminal plasma in H. huso was higher in comparison to other sturgeon species (67.2 mOsm kg-1 for A. gueldenstaedtii; 57.9 mOsm kg-1 for A. stellatus [28] and 50.8 mOsm kg-1 for A. ruthenus but it was lower than those obtained in A. persicus (82.6 mOsm kg-1) [18] and A. baerii (93.6 and 95.7 mOsm kg-1 in the first and second stripping) [29]. The intra-specific differences reported by other researchers might be addressed to secretory activity of male reproductive accessory organs (testes, testicular main ducts and sperm ducts) [15,18]. It has been frequently reported that ionic composition of the seminal plasma depends on type of hormone used for spermiation induction, dose of hormonal treatment, time of stripping after hormonal treatments and frequency of stripping [29-33]. In addition, the differences between our results and those published in literature could be related to other parameters, including origin of broodstock [29] fish management [34] spawning time [15], sampling period and sampling methods [31].
Alavi et al. and Li et al. [18,28] have reported significant positive correlations between osmolality vs. Na+ and Cl- in A. persicus and A. stellatus and A. gueldenstaedtii seminal plasma respectively. In sturgeon, sodium and chloride are the most predominant ions, suggesting a contribution to the osmolality of the seminal plasma similar to that in teleostean fish [26,27]. Probably, similar to most sturgeon in H. huso the Na+ and Cl- play a major role in maintaining the osmolality of the seminal plasma [35] and the viability of the spermatozoa in vivo [20], before their release to the environmental media and motility during the spawning activity.
Like in other sturgeon species [13,15,16,18,36] and in teleosts [5,6,21,37] the total duration of spermatozoa movement of H. huso spermatozoa is observed to increase when the K+ ion level decreases and the Na+ ion level and osmolality (up to 80 mOsmol) increases in the seminal plasma. The study performed on A. baerii shown that the motility of sperm is inhibited by high osmolality (100 mOsmol) [13]. Also, the spermatozoa of the A. persicus were motile in the osmolality range 0-100 mOsmol [18]. Therefore, osmolaity seem not to be the main inhibitor of sperm motility in seminal plasma; rather, motility is controlled by the ionic composition especially K+ ion. Studies of synergistic effects between ions have demonstrated possible control of sperm motility by the membrane potential resulting from the combined effect of several ions [38,39]. The inhibition of sperm motility by moderate concentrations of K+ can be overcome by an increase in external Ca2+ content [16]. The data in literature have indicated that the presence of external Ca2+ is a prerequisite for the initiation of sperm motility in sturgeon, and that the presence of Ca2+, similar to that of Na+, can reduce the inhibitory effects of K+ in solutions used as activation medium for sturgeon sperm [40]. There is a lack of published information on the effect of Mg2+ on spermatozoa motility in sturgeon. However, studies of the intracellular mechanisms of spermatozoa motility in teleosts have confirmed a main role for Mg2+ in its initiation, especially in de-membranated spermatozoa [37].
The present study on the quality of H. huso seminal plasma and its relationships with spermatozoa motility characteristics helps determine the optimal quality of semen to be used for artificial fertilization purposes; the seminal plasma includes Na+ (80-100 mM L-1), K+ (very low level, 0.3 mM L-1), Cl- (15 mM L-1), Ca2+ and Mg2+ (approximately 0.6 mM L-1) and also presents an osmolality in the range of 70-80 mOsm kg-1. Results of this study can be utilized for successful reproduction management and cryopreservation protocols of this endangered species.
The authors would like to thank all the technical staff of Rajaee Sturgeon Hatchery Center, (Sari, Iran) for their help and support during the conduction of this experiment.