Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 1
Although PII signal transduction proteins have been described in bacteria, archaea and plants, only one PII homolog has so far been characterized in green algae. Using a specific antibody to the PII protein from Chlamydomonas (CrPII), we provide the evidence that the model plant organism Chlorella variabilis NC64A (NC64A) possesses a protein that is immunologically related to the CrPII. The coding region of the CvGLB1 gene was sequenced, and it contained six introns and seven exons. In comparison to PII proteins from plants and cyanobacteria, the predicted amino acid sequence showed its highest degree of identity with proteins from Chlamydomonas (55.2%) and cyanobacteria, Synechococcus PCC 7942 (52.3%) and Synechocystis sp PCC 6803 (51.8%). Here, we demonstrated that like its plant PII homologs, the C. variabilis PII protein (CvPII) is appeared to be localized in the chloroplast. Expression analysis indicated that CvGLB1 transcription was independent on growth rates or nitrogen availability. The results of this study will facilitate not only characterization of PII signaling network but also the transcriptional analysis in this alga.
Keywords: PII signal transduction protein; Chlorella variabilis NC64A; GLB1 gene; Quantitative real-time RT-PCR; Gene expression; Reference gene
CrPII: PII protein of Chlamydomonas reinhardtii; CvPII: PII protein of Chlorella variabilis NC64A; NC64A: Chlorella variabilis NC64A; Ct: Threshold cycle; qRT-PCR: quantitative Realtime RT-PCR
• The coding region of the CvGLB1 gene was sequenced and characterized
• CvPII is immunologically related to CrPII and appeared to be localized in the chloroplast
• The expression of CvGLB1 gene was independent of nitrogen availability
PII signal transduction proteins constitute an abundant protein family, present in bacteria, archaea and plants [1-4]. In all cases studied so far, they coordinate the central C/N anabolic metabolism. Depending on the organisms, PII proteins control different metabolic pathways and their regulatory functions in most bacteria involve various posttranslational modifications [2]. Functionally, these proteins can be regarded as signal integrators, whereby the various signals are integrated by the inter-conversion of different PII states. Depending on these states, PII proteins can bind to various receptors, affecting all levels of metabolic regulation (metabolic enzymes, gene expression, transport activity). Interestingly, although the structure of PII proteins is highly conserved, the PII regulated targets are distinct in different phylogenetic lineages. The cyanobacteria and plant PII proteins regulate the controlling enzyme of arginine biosynthesis, N-acetyl-Lglutamate kinase (EC 2.7.2.8) [2,5]. The high degree of conservation in sequence, structure, and function between Synechococcus elongatus and Arabidopsis thaliana PII-NAGK complexes implies a strong selective pressure for maintaining PII-regulated arginine biosynthesis in the evolution of plastids from an ancestral cyanobacterium [6]. In addition, the key enzyme of fatty acid synthesis, acetyl-CoA carboxylase (EC 6.4.1.2), seems to be a target of PII control [7]. Furthermore, there are indications that PII might support other, yet uncharacterized PII-specific roles within the plant cell [3]. To study plastid-localized PII signaling, the simple unicellular green algae represent the most suitable model systems. The green algae are a phylogenetic link between the cyanobacterial ancestor that gave rise to chloroplasts via endosymbiosis and higher plants. The only algae PII protein that has been studied in depth is that of Chlamydomonas reinhardtii [5,8]. Our recent comparative studies on ChlamydomonasPII protein has led to the identification of a general glutamine sensing mechanism in Chloroplastida (plant kingdom) [5]. However, the question, whether glutamine sensing is specific for PII-NAGK interaction or whether PII proteins from Chloroplastida also transmit glutamine signals to other targets remains unclear. The characterization of additional PII proteins in unicellular green algae is now required to understand their roles in Chloroplastida. To study fundamental problems in plant physiology, another unicellular green alga, Chlorella variabilis NC64A, has been recognized as the excellent model organism [9]. For this reason, we started studying PII protein in C. variabilis NC64A. However, the CvPII sequence (GenBank: XP_005842899) was incomplete on amino acid end. In this report we identified the complete amino acid sequence for the C. variabilis GLB1 gene product as deduced from the cognate CDS, and examined the sequence conservation among PII proteins from cyanobacteria and Chlorophyta. The relative abundance of CvGLB1 transcripts from NC64A cells grown in different nitrogen-replete and in nitrogen-deficient media has been estimated using quantitative real-time RT-PCR. Here, it is shown that this gene is invariably expressed under all the conditions used in this work. A selection of CvGLB1 is proposed as reference gene. This is the first report to evaluate reference gene for the transcriptional analysis of algae of the Chlorella genus.
Strains and culture conditions
C. variabilis NC64A was obtained from Dr. J. Van Etten, University of Nebraska-Lincoln, USA. NC64A cells were grown in MBBM [10,11] or C-medium [12,13] under continuous light at ~ 30 μmol m-2 s-1 and shaking at 25°C. To determine growth rates, different media were used depending on the nitrogen source: the BBM (MBBM-N) medium containing 25 mM NH4Cl or the BBM medium containing 2, 10 or 15 mM arginine. The number of cells was recorded microscopically with use of a counting chamber. The growth rate was evaluated as a doubling of cell number per hour.
Chlorophyll content was determined using acetone extraction. 0.6 mL of culture was centrifuged and the pellet resuspended in 83% acetone and vortexed with zirconia-silica beads (0.1 and 0.5 mm) using homogenizer Minilys (Bertin Technologies, France) to extract pigments. Cellular debris was pelleted by centrifugation and chlorophyll a and b levels were determined spectrophotometrically (SmartSpec Plus, BioRad) in the supernatant, by measuring optical absorbance at 645 nm and 663 nm. Calculations of total chlorophyll (μg mL-1) were performed as previously described [14]. Mean cell diameter was determined microscopically (Leica TCS-SP5, Leica-Microsystems, Germany) using Leica Application suite.
Chlamydomonas reinhardtii strain CC-124 (mt‾), obtained from the Chlamydomonas Culture Collection at Duke University, Durham NC, USA, was used as the control in Western blotting. Chlamydomonas cells were grown at 22°C under a 12-h light/12-h dark regime in Trisacetate- phosphate (TAP) medium [15].
Protein isolation, SDS-PAGE and Western-blot analysis
C. variabilis NC64A (1–2×107 cells mL-1 in 100 mL) and C. reinhardtii CC-124 (2–4×106 cells mL-1 in 100 mL) were collected by centrifugation (3,000g, 5 min) and resuspended in 0.1M DTT, 0.1M Na2CO3. Chlorella samples were additionally sonicated and vortexing at room temperature for 10 min. The protein concentration was determined by staining with amido black, using BSA as a standard [16]. After separation of the proteins by SDS-PAGE on a 15% polyacrylamide gel [17], they were transferred to nitrocellulose membranes (Protran, Whatman) by a semidry blotting (Trans-blot SD, BioRad). Blots were blocked in 5% milk in Tris-buffered saline solution with 0.1% Tween 20 prior to an incubation of 1 h in the presence of anti-CrPII primary antibodies [8]. Specific antibodies against HSP70B [18] were kind gifts from Dr. Michael Schroda (Technische Universität Kaiserslautern, Germany). The dilutions of the primary antibodies used were 1:5,000. A 1:10,000 dilution of horseradish peroxidase-conjugated anti-rabbit serum (Sigma) was used as a secondary antibody. The peroxidase activity was detected by an enhanced chemiluminescence assay (Promega).
Cloning and sequencing of DNA fragments
For isolation of genomic DNA Chlorella cells (1–2×107 cells mL-1 in 100 mL) were collected by centrifugation (3,000g, 5 min) and resuspended in 50 mM Tris-HCl (pH 7.5), 300 mM NaCl, 10 mM EDTA, 4% SDS. Then, Chlorella samples were vortexing with zirconia-silica beads (0.1 and 0.5 mm) using homogenizer Minilys (Bertin Technologies, France). PCR procedures are illustrated in Supplemental Figure 1S. Each PCR was made in a final volume of 25 μL with the following components: 0.2 pmol of each primer, 0.2 mM dNTPs, 0.5U Taq DNA polymerase (SybEnzyme, Russia), 2.5 μL specific buffer (containing 2.5 mM MgCl2), 50–100 ng DNA, and 2–3% dimethylsulfoxide (DMSO). PCR-amplification was performed like described earlier [19]. For band reamplification, we inserted a 200-μL pipette tip directly into the agarose gel in just the desired band. Specific PCR products were isolated from the gel using QIAquick Gel Extraction kit (QIAGEN, Germany), cloned using the pAL-TA PCR product cloning kit (Evrogen, Russia), and then transformed into Escherichia coli DH5α cells. DNA sequencing was performed at the Research resource center for molecular and cell technologies of Saint- Petersburg State University with Big Dye (R) Terminator v3.1 Cycle Sequencing Kit (Life Technologies, USA) in an automatic sequencer (model 3500xL Genetic Analyzer, Life Technologies, USA).
Synthesis of cDNA, RT-PCR and real time qPCR
Total RNA was isolated as described previously [20] with the following modifications: C. variabilis NC64A (1–2×107 cells mL-1 in 100 mL) were harvested and then the pellets were resuspended in Trizol reagent (Invitrogen, USA) and vortexing with zirconia-silica beads (0.1 and 0.5 mm) to ensure the rupture of the algal cell walls. The quality of the RNA preparations was estimated by agarose gel electrophoresis, and RNA concentration and purity were determined spectrophotometrically (SmartSpec Plus, Bio-Rad). RNA (1μg) was treated with RNase-Free DNase I (Fermentas) in 10 μl at 37°C for 15 min. The reaction was stopped with 0.43 μL of 50 mM EDTA at 80°C for 10 min.
Reverse transcription was performed with RevertAid H Minus First Strand cDNA Synthesis Kit according to the manufacturer´s instructions (Thermo Scientific). Primers for RT-qPCRs were chosen based on ≥ 90% primer efficiency and on a single melt curve. They are listed in Supplemental Table 1S. Each reaction contained the master mix, 5% DMSO, 200 nM of each primer, and cDNA corresponding to 10 ng input RNA in the reverse transcriptase reaction. Real time qPCR was performed on the Light Cycler Instrument (CFX96 Real-Time PCR Detection System, Bio Rad) using SYBR Green I as a fluorescent dye. The reaction conditions were as follows: 95°C for 3 min, followed by cycles of 95°C for 30 s, and 72°C for 60 s. up to a total of 40 cycles. All reactions were performed in triplicate with at least two biological replicates. Controls without template or reverse transcriptase were included. The threshold cycle (Ct), defined as the fractional cycle at which the fluorescence signal becomes significantly different from the baseline was manually determined by the fit-point method provided by the LightCycler software. Fold difference was calculated as 2ΔCt.
Accession numbers
The accession numbers for the genes used in this article are as follows: GLB1 (GenBank: KJ524571), EF1A (GJI: 30705, GenBank: 17356204), GPD1 (GJI: 23323, GenBank: EFN55541), MDH3 (GJI: 59812), ACT (GJI: 56324), PYK (GJI: 34881, GenBank: XM_005849050.1), CBLP (GJI: 31033, GenBank: EFN55888.1), UBI (GJI: 59526, GenBank: 17359075), AMT1 (GenBank: 17352992).
Immunological identification of CvPII
The first step in characterization of the Chlorella PII was its identification. A search of the C. variabilis NC64A databases identified candidate protein (GenBank: XP_005842899, EFN50797), which we designated as CvPII. The deduced amino acid sequence of CvPII shares high overall identity with PII from Chlamydomonas reinhardtii (55.2%). We have used a polyclonal antibody against the C. reinhardtii PII (CrPII) [8]. As shown in (Figure 1A), the antibody reacted with a protein from C. variabilis with the molecular weight (~17 kDa) similar to mature PII from C. reinhardtii. Additionally, the protein corresponding to the predicted full-length CrPII size (~22 kDa) was also detected in cell total proteins (Figure 1A). Hence, the apparent molecular weight of CvPII is significantly larger than the 11.3 kDa calculated from the amino acid sequence of XP_005842899.
Sequence analysis of the gene encoding CvPII protein
A number of molecular techniques used [9] were not suitable to complete the coding sequence encoded a PII protein that often occur in organisms with a high-complexity genome such as Chlorella. To isolate the region adjacent to the known part of the gene, we set up a PCR approach that allows amplification of products in a consistent and efficient way with enough quality for direct sequencing. This PCR amplification technique is based on the random distribution of frequent restriction sites in a genome and on a special design of primers [21]. All of the steps, cycles and theoretical products obtained, and sequences of the primers used are summarized in Supplemental Figure 1S. Four restriction sites (PvuII, BanI, PstI and TaqI) were used for the corresponding primers. To reduce the probability of finding these sites in the genome, one extra nucleotide after the restriction site at the 3´end was also added. As a result, after the second amplification round, specific products usually appeared in the gel as high-intensity bands that were directly sequenced. The PCR performed on genomic DNA using primers DegTaqI/GLB1R2 and Q0/GLB1R1 allowed us to isolate a 2,196 bp fragment whose sequence corresponded to the gene, which was designated as CvGLB1 (GenBank: KJ524571). The amplification of an approximately 896-bp internal fragment (GLB1F1/GLB1R1) suggests the specificity of the PCR product. As shown in Figure1B, the coding region of the CvGLB1 gene contained six introns and seven exons. The sizes of the introns ranged from 87 bp to 348 bp. The splice sites for the exon/intron and intron/exon junctions perfectly fitted the two consensus motifs G↓GTGAG and (C/A)CAG↓G (invariant bases are shown in bold) identified in nuclear genes [9]. Using primers GLB1F3/GLB1R3, GLB1F4/GLB1R4 and GLB1F5/GLB1R5, designed from different exons, a PCR amplification on cDNA yielded fragments of the expected sizes (Figure 1B, Supplemental Table 1S). The CDS encoded a protein of 210 amino acids with calculated weight of 21.96 kDa. This size corresponded to the band of about 22 kDa detected by immunological identification (Figure 1A). A potential chloroplasttargeting presequence of 47 amino acid residues could be identified in the protein sequence (ChloroP 1.1 and TargetP1.1 Center for Biological Sequence Analysis) [22,23]. If the mature CvPII corresponded to the sequence of 162 amino acids, its predicted molecular weight should be 17.1 kDa. The second band corresponding to a protein of this size was immunologically detected in the total cell extract proteins, together with the unprocessed form (Figure 1A). The data suggest that like its plant PII homologs [8,24,25], CvPII protein may reside in chloroplast.
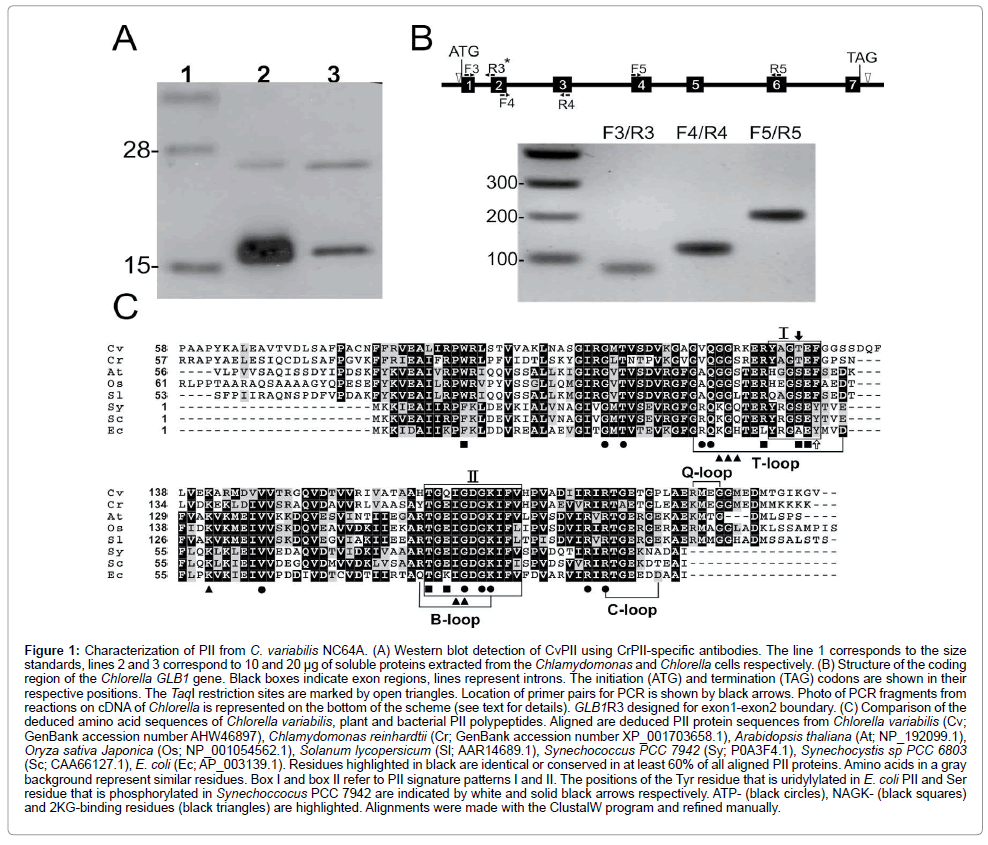
Figure 1: Characterization of PII from C. variabilis NC64A. (A) Western blot detection of CvPII using CrPII-specific antibodies. The line 1 corresponds to the size standards, lines 2 and 3 correspond to 10 and 20 μg of soluble proteins extracted from the Chlamydomonas and Chlorella cells respectively. (B) Structure of the coding region of the Chlorella GLB1 gene. Black boxes indicate exon regions, lines represent introns. The initiation (ATG) and termination (TAG) codons are shown in their respective positions. The TaqI restriction sites are marked by open triangles. Location of primer pairs for PCR is shown by black arrows. Photo of PCR fragments from reactions on cDNA of Chlorella is represented on the bottom of the scheme (see text for details). GLB1R3 designed for exon1-exon2 boundary. (C) Comparison of the deduced amino acid sequences of Chlorella variabilis, plant and bacterial PII polypeptides. Aligned are deduced PII protein sequences from Chlorella variabilis (Cv; GenBank accession number AHW46897), Chlamydomonas reinhardtii (Cr; GenBank accession number XP_001703658.1), Arabidopsis thaliana (At; NP_192099.1), Oryza sativa Japonica (Os; NP_001054562.1), Solanum lycopersicum (Sl; AAR14689.1), Synechococcus PCC 7942 (Sy; P0A3F4.1), Synechocystis sp PCC 6803 (Sc; CAA66127.1), E. coli (Ec; AP_003139.1). Residues highlighted in black are identical or conserved in at least 60% of all aligned PII proteins. Amino acids in a gray background represent similar residues. Box I and box II refer to PII signature patterns I and II. The positions of the Tyr residue that is uridylylated in E. coli PII and Ser residue that is phosphorylated in Synechoccocus PCC 7942 are indicated by white and solid black arrows respectively. ATP- (black circles), NAGK- (black squares) and 2KG-binding residues (black triangles) are highlighted. Alignments were made with the ClustalW program and refined manually.
Protein sequence similarities and structural prediction
The CvPII deduced protein sequence was compared to PII proteins from Chlamydomonas, three higher plants and two cyanobacteria (Figure 1C). The predicted amino acid sequence showed its highest degree of identity with proteins from Chlamydomonas (55.2%) and cyanobacteria, Synechococcus PCC 7942 (52.3%) and Synechocystis sp PCC 6803 (51.8%). Similar to PII homologs of Chlamydomonas and higher plants, CvPII has unique N- and C-terminal sequences that are absent from prokaryotic proteins. The region of homology to prokaryotic PII encompasses the entire bacterial PII protein and corresponds to the segment between amino acids 82 and 189 of the CvPII protein (Figure 1C). In this region, the overall identity between the encoded CvPII protein and E. coli PII is 48.5%. Within the region corresponding to E. coli PII, the deduced amino acid sequence of CvPII shares high overall identity with representative plant and cyanobacterial PII sequences including: C. reinhardtii (65.7%), A. thaliana (54.7%), Oryza sativa Japonica (53.7%), Solanum lycopersicum (53.7%), Synechococcus PCC 7942 (53.9%) and Synechocystis PCC 6803 (53.4%).
In addition to the high overall identity between various PII proteins of cyanobacteria, plants and Chlorella, extremely high local identities occur over two signature patterns that have been defined at the PROSITE (PS00496 and PS00638) (Figure 1C). In Proteobacteria the signature pattern I (Y-[KR]-ly-[AS]-[AE]-Y) contains the Tyr-51 residue that is posttranslational modified by uridylylation [26]. In the primary sequence of CvPII as well as in proteins of Chlamydomonas and some higher plants, the corresponding Tyr-51 residue is replaced with Phe [8]. This indicates that the coding sequence of the Chlorella PII protein does not comprise an uridylylatable Tyr. It is important to emphasize that such cyanobacterial PII proteins that contain a conserved Tyr-51 residue in this region are not modified by uridylylation. Instead, they are phosphorylated at Ser-49 residue [27]. PII proteins of higher plants contain homologous Ser-49 residue, but apparently they are not regulated by phosphorylation [28]. CvPII contains a threonine residue at this site (Figure 1C). This could suggest that CvPII protein may not be modified by phosphorylation similarly to PII protein of Chlamydomonas [8]. Furthermore, the C-terminus of CvPII contains the region between amino acids 196 and 203 (RMEGGMED) that is identical to motif in the C-terminal sequence of PII C. reinhardtii. In CrPII, the C-terminal extension forms a small loop structure, the Q-loop (KMEG) that wraps around the bound glutamine molecule and is required for glutamine-dependent complex formation with N-acetyl- L-glutamate kinase [5]. The C-terminal residues of the Q-loop are part of a motif present in CvPII, suggesting that it may act as glutamine sensor via the C-terminal Q-loop extension.
The sequence [ST]-x (3)-G-[DY]-G-[KR]-[IV]-[FW]-[LIVM], which was found in all prokaryotic PII-proteins and corresponds to the B-loop is also conserved within Chlorella, Chlamydomonas and higher plants. This motif is involved in ATP binding between two adjacent subunits of the PII protein trimers. Examination of the aligned sequences shows that residues necessary for formation of the bacterial and plant PII trimeric structures are conserved in Chlorella PII (Figure 1C). These results may indicate that CrPII forms homotrimers just like other proteins of its family [2,8]. Moreover, residues involved in effector molecule binding are also conserved. These findings suggest that CvPII may function as a signaling protein. Elucidation of the precise role of PII in Chlorella, however, requires background data on its expression.
Expression of GLB1 Gene in C. variabilis NC64A
Growth of NC64A under various nitrogen conditions: PIIdependent regulation of nitrate/nitrite transport was shown in cyanobacteria and Arabidopsis [29-31]. In contrast to many cyanobacteria, algae and higher plants, NC64A did not grow on nitrate or nitrite [13]. In ammonia-supplemented medium, C. variabilis NC64A displayed significantly lower growth rates than in MBBM medium containing bacto-peptone as a nitrogen source (Figure 2). The growth pattern was accompanied by a reduction of chlorophyll content, especially in stationary phase (Figure 2A). These values were consistent with data published previously [13]. L-arginine may be used as sole nitrogen source by C. variabilis NC64A [32]. To our data, this strain showed the highest growth rate and maximum cell concentrations with 10 mM of arginine in the medium (Figure 2). NC64A reached stationary phase at ~2×107 cells per ml (Figure 2A). When the strain was grown on increasing arginine concentrations (15 mM), no differences were observed in growth rates compared to the medium containing 10 mM arginine (Figure 2B). Chlorophyll content was also determined. In spite of the fact that the lowest chlorophyll content is attained in the cells grown in MBBM, they had the smallest average cell diameter (4 μm and 4.2 μm at the mid-exponential and stationary phase of growth). Intermediate cell diameters for agrininesupplemented media were 4.9 μm and 5.2 μm, respectively. Although progress in understanding central nitrogen anabolic metabolism in Chlorella has been limited, progress on PII-based control will be aided by gene expression advances.
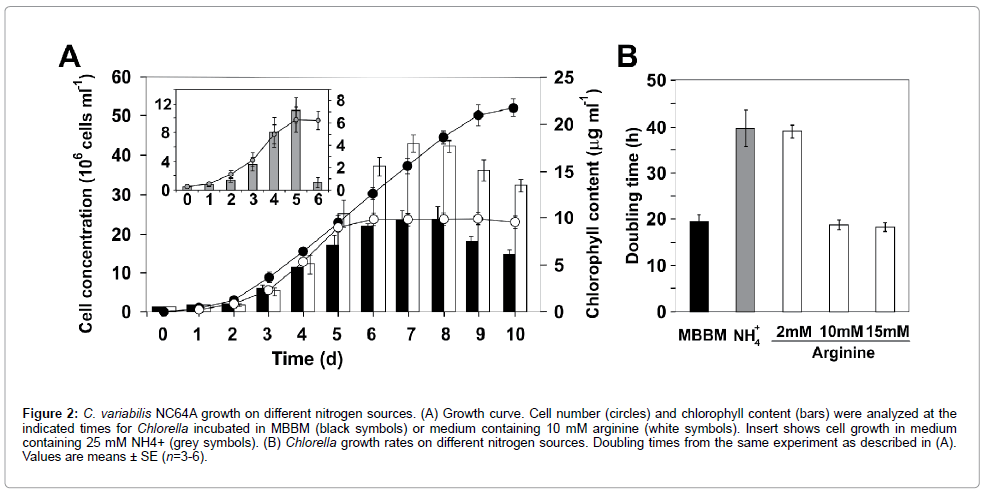
Figure 2: C. variabilis NC64A growth on different nitrogen sources. (A) Growth curve. Cell number (circles) and chlorophyll content (bars) were analyzed at the indicated times for Chlorella incubated in MBBM (black symbols) or medium containing 10 mM arginine (white symbols). Insert shows cell growth in medium containing 25 mM NH4+ (grey symbols). (B) Chlorella growth rates on different nitrogen sources. Doubling times from the same experiment as described in (A). Values are means ± SE (n=3-6).
Selection of reference genes for quantitative real-time RT-PCR expression studies: For reliable gene expression analysis, normalization of gene expression data against reference genes is required. However, evaluation of internal control genes has not been conducted in Chlorella. A search of the C. variabilis NC64A databases identified candidates for reference genes encoding putative GPD1 (glyceraldehyde-3-phosphate dehydrogenase), EF1A (elongation factor 1α), ACT (actin), MDH3 (malate dehydrogenase), PYK (pyruvate kinase), UBI (ubiquitinconjugating enzyme E2) and CBLP (receptor of activated protein kinase C 1). We first analyzed the behavior of selected putative housekeeping genes. The distribution of Ct levels is given in Supplemental Figure 2S. A candidate reference gene was defined as a gene with the most stable expression, i.e. a gene with a small coefficient of variation (CV) and a maximum fold change <2 (MFC, the ratio of the maximum and minimum values observed within the dataset) [33]. All 7 genes had summarized CV above the 4% level (Table 1). Thus, GPD1, EF1A, ACT, MDH3, PYK, UBI and CBLP were not applicable here because of too large a variability under various nitrogen conditions
| Gene symbol | MBBM | Arginine-supplied medium | Ammonium-supplied medium | Nitrogen-free medium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mean | SD | V(%) | MFC | mean | SD | V(%) | MFC | mean | SD | V(%) | MFC | mean | SD | V(%) | MFC | |
| UBI | 25.88 | 0.683 | 2.65 | 1.11 | 27.20 | 1.319 | 4.85 | 1.14 | 28.29 | 0.534 | 1.89 | 1.06 | 27.70 | 0.739 | 2.67 | 1.06 |
| CBLP | 22.75 | 1.323 | 5.82 | 1.16 | 23.97 | 1.259 | 5.25 | 1.06 | 24.94 | 0.552 | 2.21 | 1.08 | 24.51 | 1.131 | 4.62 | 1.11 |
| EF1A | 17.45 | 1.902 | 10.90 | 1.34 | 20.03 | 0.508 | 2.53 | 1.09 | 20.68 | 0.874 | 4.23 | 1.13 | 18.48 | 1.169 | 6.33 | 1.15 |
| GPD1 | 26.21 | 2.149 | 8.20 | 1.28 | 30.34 | 0.994 | 3.28 | 1.10 | 29.78 | 1.189 | 3.99 | 1.12 | 28.57 | 1.667 | 5.83 | 1.15 |
| PYK | 24.58 | 1.758 | 7.15 | 1.24 | 27.93 | 0.812 | 2.91 | 1.08 | 27.93 | 1.204 | 4.31 | 1.12 | 25.65 | 1.097 | 4.28 | 1.12 |
| MDH3 | 21.54 | 1.876 | 8.71 | 1.25 | 24.35 | 1.318 | 5.41 | 1.16 | 24.65 | 1.124 | 4.56 | 1.12 | 23.95 | 0.420 | 1.76 | 1.04 |
| AKT | 26.20 | 1.029 | 3.93 | 1.11 | 29.56 | 1.035 | 3.50 | 1.11 | 29.01 | 0.941 | 3.24 | 1.09 | 28.47 | 0.808 | 2.84 | 1.07 |
Table 1: The variation in expression of the candidate reference genes in C. variabilis NC64A.
Expression analysis of CvPII under various nitrogen conditions: To test the expression patterns of CvGLB1 gene, we carried out a comparative qRT-PCR analysis on cells in exponential (early, mid and late) and stationary phases of culture grown on MBBM, arginine- and ammonium-supplemented media. The mean Ct data in the samples are shown in Figure 3A. Remarkably, this gene had CV beneath the 4% level under all nitrogen conditions.
To prove that CvGLB1 is expressed at a constant level under various nitrogen conditions, normalization against internal control genes was used (Figures 3B and 3C). UBI was used as internal control for cells grown in MBBM, while ACT was used for cells grown in arginine as nitrogen source (Figure 3C). In arginine-containing medium, the relative expression levels of CvGLB1 were similar in all types of cells, suggesting that CvGLB1 transcription was unaffected by phase of growth (Figure 3B). CvPII protein levels were also quite constant during growth (Figure 3B), upper panel. These data are in agreement with the precise regulation of PII reported in Chlamydomonas [5,8]. The switch from arginine to MBBM medium did not lead to a change in CvGLB1 transcript or CvPII protein levels (Figure 3B). These results clearly indicated that CvGLB1 expression is independent on growth rates or nitrogen source. Analysis of transcription profiles of CvGLB1 gene revealed that it showed stable expression and can therefore be recommended as reference gene.
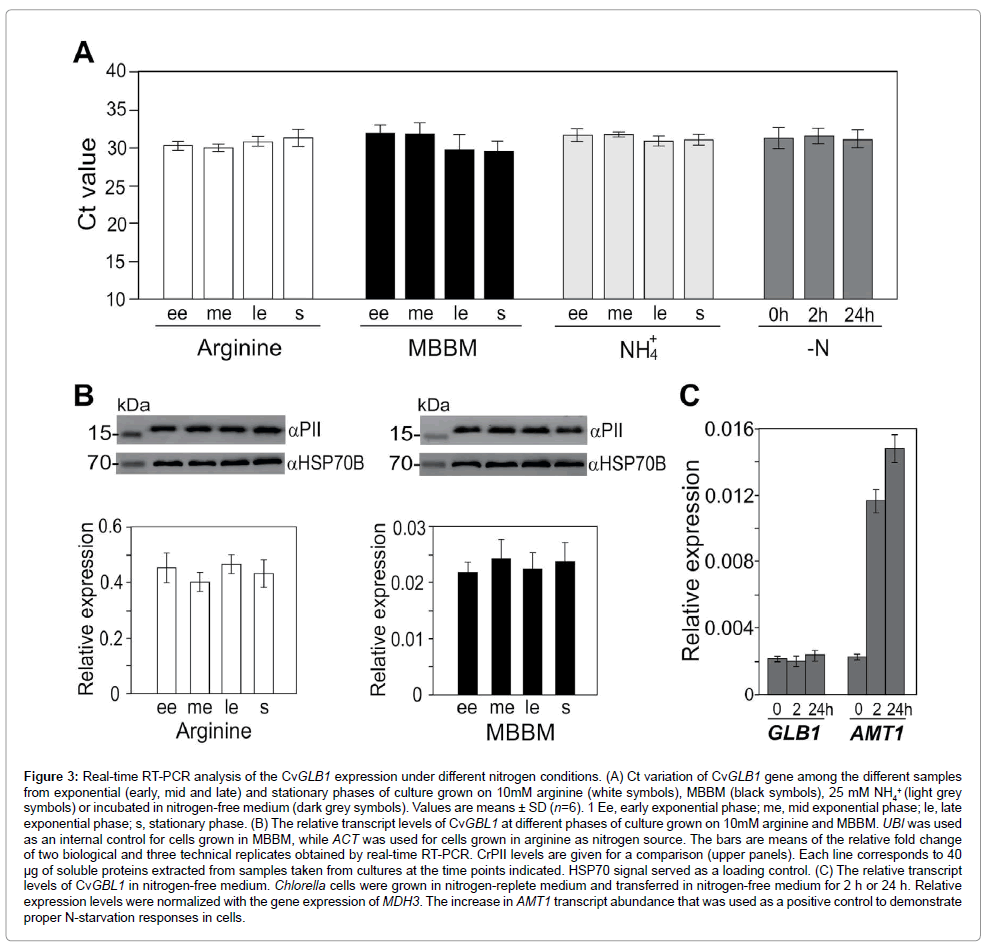
Figure 3: Real-time RT-PCR analysis of the CvGLB1 expression under different nitrogen conditions. (A) Ct variation of CvGLB1 gene among the different samples from exponential (early, mid and late) and stationary phases of culture grown on 10mM arginine (white symbols), MBBM (black symbols), 25 mM NH4 + (light grey symbols) or incubated in nitrogen-free medium (dark grey symbols). Values are means ± SD (n=6). 1 Ee, early exponential phase; me, mid exponential phase; le, late exponential phase; s, stationary phase. (B) The relative transcript levels of CvGBL1 at different phases of culture grown on 10mM arginine and MBBM. UBI was used as an internal control for cells grown in MBBM, while ACT was used for cells grown in arginine as nitrogen source. The bars are means of the relative fold change of two biological and three technical replicates obtained by real-time RT-PCR. CrPII levels are given for a comparison (upper panels). Each line corresponds to 40 μg of soluble proteins extracted from samples taken from cultures at the time points indicated. HSP70 signal served as a loading control. (C) The relative transcript levels of CvGBL1 in nitrogen-free medium. Chlorella cells were grown in nitrogen-replete medium and transferred in nitrogen-free medium for 2 h or 24 h. Relative expression levels were normalized with the gene expression of MDH3. The increase in AMT1 transcript abundance that was used as a positive control to demonstrate proper N-starvation responses in cells.
Ammonium depletion has recently been shown to affect GLB1 transcript level in Chlamydomonas [8]. To test if in C. variabilis cells GLB1 is controlled by nitrogen deficiency at the transcriptional level, cells were grown in nitrogen-replete medium (the mid-exponential phase of growth) and then transferred to nitrogen-free medium (Figures 3A and 3C). The gene (GenBank: 17352992), which is similar to ammonium transporter 1 member 2 from A. thaliana, exhibited an increase of transcript level to nitrogen-deficiency, as previously reported for Chlamydomonas AMT1 genes [20,34]. The increase in AMT1 transcript abundance was used as a positive control to demonstrate proper nitrogen-starvation responses in cells. We conclude that the expression of CvGLB1 gene was independent of nitrogen availability. In cyanobacteria the expression of glnB gene encoding PII is under the control of nitrogen, and protein activity is regulated on the posttranslational level [35]. Protein sequence similarities and structural prediction suggest that the regulation of CvPII may depend on other mechanisms typical for members of the plant PII proteins including allosteric regulation by glutamine. In agreement, the residues involved in glutamine binding via the C-terminal Q-loop extension are conserved in CvPII (Figure 1C), suggesting that it may act as glutamine sensor. However, details of the glutamine sensing mechanism of Chlorophyta PII need to be clarified. Because of the amenability of C. variabilis NC64A to molecular analysis, the results from this study will enable us to further dissect the PII signaling network in this alga. The identification of CvPII partners is now required to help to understand the role(s) of the PII protein and how it is involved in host-symbiotic relationships.
PII signal transduction proteins are among the most abundant signalling proteins in nature. Our results provide crucial information for sequence and transcriptional analysis of CvGLB1 gene encoding PII in C. variabilis NC64A, which constitutes an excellent model system for the study of all aspects of plant physiology, including the molecular mechanisms underlying regulation of metabolism. Expression analysis indicates that CvGLB1 transcription is independent on growth rates or nitrogen availability, suggesting that it may be used as a reference gene. We also demonstrated that CvPII shares high sequence identity with PII proteins from plants and cyanobacteria. Specifically, we detected that CvPII is immunologically related to Chlamydomonas PII and appeared to be localized in the chloroplast. Protein sequence similarities and structural prediction suggest that CvPII may function as a signaling protein.
E. Minaeva performed the experiments; E. Ermilova designed the experiments and wrote the manuscript.
We thank Dr. Michael Schroda (University of Kaiserslautern, Germany) for providing antibodies against HSP70B. This work was supported by Saint- Petersburg State University (research grant N 1.38.211.2014) and Russian Foundation of Basic Research to EVE (N 13-04-00087). DNA sequencing was performed at the Research resource center for molecular and cell technologies of Saint-Petersburg State University. Light microscopy and image processing was made on the base of the Core Facility «CHROMAS» of Saint-Petersburg State University.