Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2014) Volume 3, Issue 2
In multicellular organisms, growth and development need to be precisely coordinated and are strongly relying on positional information. Positional control is achieved through exchanges of molecular messages between cells and tissues by means of cell-to-cell communication mechanisms. Especially in plants, accurate and well-controlled cell-to-cell communication networks are essential because of the complete absence of cell mobility and the presence of rigid cell walls. For many years, phytohormones were thought to be the main messengers exchanged between cells. Nevertheless, identification of systemin as the first plant signaling peptide in tomato hinted that peptide hormones were acting in plants as they were in animals. During the last decade, our knowledge of plant signaling peptides has progressed considerably and a number of signaling peptide families have been discovered and partially characterized. Here, we provide an overview of the current knowledge in signaling peptides in the model species Arabidopsis thaliana and discuss their proposed functions during plant growth and development.
<Response to developmental cues and environmental stimuli in multicellular organisms is highly dependent on cell-to-cell communication. For many years, the so-called classical phytohormones - auxin, cytokinin, abscisic acid, gibberellin and ethylene - were thought to be the main players of intercellular communication in plants. Later on, several other molecules, including brassinosteroids, jasmonate, salicylic acid and strigolactones, have been added to the list of plant growth regulators [1,2]. However, it remained puzzling that such a relatively small group of molecules could coordinate multiple and very diverse cellular responses. Research over the last decade made clear that, besides the plant hormones, other molecules are important players in cell-to-cell communication networks, including signaling peptides (also called “peptide hormones” or “secreted peptides”), small RNAs and transcription factors [1,3,4]. Secreted peptides are now fully recognized in plants as molecular messengers because of their involvement in key developmental processes such as meristem maintenance, organ abscission, cell elongation, cell proliferation and differentiation, gravitropism and defense against biotic aggressors [5-10]. Here, we will describe recent advances made in the characterization of signaling peptide function during plant development including a reflection on some recent developments in signaling peptide identification strategies.
Peptides are generally defined as small proteins containing 50 or fewer amino acids in their mature form [4]. Systemin was the first plant signaling peptide identified and was shown to be involved in systemic response to wounding in tomato [11]. Following the discovery of systemin, several signaling peptides have been characterized in plants, especially in Arabidopsis (Table 1). Nevertheless, considering the myriad of signaling peptides in mammals [12], it is likely that many more still remain to be identified.
| Peptide family | Mature peptide | Gene family in Arabidopsis |
Conserved motif | Receptor | Function | References |
|---|---|---|---|---|---|---|
| CLAVATA3/ENDOSPERM SURROUNDING REGION (ESR)-RELATED | CLV3/CLE | 32 | CLE or mCLV3, 12–14 AA |
CLV1, BAM1, CLV2, RPK2, ACR4 |
Stimulates organogenesis and inhibits meristematic growth; can stimulate vascular development |
[41] [5] [98] [99] [28] [100] [101] [58] |
| C-TERMINALLY ENCODED PEPTIDE | CEP | 15 | CEP1, 14 AA | - | Inhibits root growth | [29] [23] |
| EMBRYO SURROUNDING FACTOR1 | ESF1 | 3 | ESF1, 68 AA | SSP | Early embryo patterning in flowering plants | [94] |
| EPIDERMAL PATTERNING FACTOR /EPF-LIKE | EPF | 11 | - | TMM, ER, ERL1, ERL2 |
Promotes epidermal cell division leading to guard cell (stomata) formation |
[18] [19] [72] |
| GOLVEN/ROOT GROWTH FACTOR/CLE-LIKE | GLV/RGF/CLEL | 11 | 13-18 AA | - | Maintain root stem cell Niche, gravitropic response, lateral root and root hair development. |
[9] [10] [22] [46] |
| INFLORESCENCE DEFICIENT IN ABSCISSION and IDA-LIKE | IDA /IDL | 6 | EPIP | HAE, HSL | Inhibits floral abscission Lateral root primordium development, cell separation |
[7] [69] [71] |
| PHYTOSULFOKINE | PSK | 6 | PSK-a, 5 AA | PSKR1 | Promotes cell proliferation and longevity, root and hypocotyl elongation |
[24] [102] [86] [87] [88] |
| PLANT NATRIURETIC PEPTIDE | PNP | 2 | - | - | Extracellular, cell expansion, water/ion movement, stomatal opening, inhibits ABA-induced stomatal closure |
[103] [104] [105] [106] [107] [108] [109] |
| PLANT PEPTIDE CONTAINING SULFATED TYROSINE | PSY | 3 | PSY1, 18 AA | PSYR1 | Promotes cellular expansion and proliferation, upregulated by wounding |
[8] |
| POLARIS | PLS | 1 | 36 AA | - | Required for root elongation, lateral root formation, leaf vascular patterning |
[97] [110] |
| PROPEP | AtPep | 7 | Pep1, 23 AA | Pep1R | Promotes innate immune responses {a danger signal} |
[111] [112] [113] [114] |
| RAPID ALKALINIZATION FACTOR and RALF-LIKE |
RALF/RALFL | 39 | RALF, 25-30 AA |
FERONIA | Associated with danger signals, affects growth-inhibits root growth |
[6] [14] [90] [115] |
| ROT FOUR LIKE/ DEVIL | RTFL/DVL | 24 | Not processed | - | Polarity, cell porolifration, leaf development | [116] [117] [118] |
| S_LOCUS CYSTEINE_RICH PROTEIN/SCR_LIKE | SCR/SCRL | 28 | Not processed | SRK | Prevents self-fertilization | [15] [119] |
| SYSTEMIN | - | Not present | 18AA | SR160 | Systemin defense response | [11] |
| TAPETUM DETERMINANT | TPD | 2 | TPD | EMS1 | Anther development Promoting tapetum formation |
[42] [120] |
Table 1: Plant peptide signaling molecules identified so far
Based on structural characteristics, signaling peptides are divided into two distinct groups [13]. Group I consists of cysteine-rich peptides in which intramolecular disulfide bonds determine the three-dimensional structure of the mature protein. Peptides in this group may or may not undergo proteolytic processing. Mature cysteine-rich peptides are generally longer than 20 amino acids [14]. Secreted peptides belonging to this class include the S-locus Cys-rich or S-locus protein 11 (SCR/SP11) [15,16], LURE [17], RAPID ALKALINIZATION FACTORs (RALFs) [6] and EPIDERMAL PATTERNING FACTORs (EPFs) [18,19] including STOMAGEN [20].
Group II includes cysteine-poor peptides, also called “small posttranslationally modified peptides”. In contrast to the aforementioned group, this class is characterized by the small size of the mature peptide (<20 amino acids) always resulting from proteolytic processing, and the presence of a C-terminal conserved motif that often carries proline residues and posttranslational modifications. Like cysteine-rich peptides, they are generally encoded by multiple paralogous genes. Most signaling peptides characterized so far in plants belong to this class. Small posttranslationally modified peptides share a common tripartite structure: (i) a signal peptide at the N-terminal region, (ii) a C-terminal region that is usually conserved among different members of the family and corresponds to the mature peptide and (iii) a variable segment that links the two terminal domains (Figure 1).

Figure 1: Structure of Group II signaling prepropeptides. SP, signal peptide recognized and cleaved upon secretion. CM, conserved motif yielding the mature peptide.
In contrast to the majority of the peptides, which are produced by the cleavage of larger precursors, EARLY NODULIN40 (ENOD40) peptides are encoded as such [21]. Interestingly, it has been shown that several signaling precursors contain multiple copies of the C-terminal motif or exhibit a second conserved one in the variable segment [22,23]. However the functional consequences of these peculiar arrangements remain unknown.
In plants, the C-terminal region of small posttranslationally modified peptides often carries one or more of three types of posttranslational modifications: tyrosine sulfation [8-10,24], which is catalyzed by the plant-specific TYROSYL PROTEIN SULFOTRANSFERASE1 (TPST) [25], proline hydroxylation [8,9,26-30] that is mediated by PROLYL 4-HYDROXYLASE (P4H) [31] and hydroxyproline arabinosylation. Hydroxyproline arabinosylation [8,30] is a plant-specific posttranslational modification [32] and, only recently, three HYDROXYPROLINE O-ARABINOSYLTRANSFERASE (HPAT) enzymes have been discovered in Arabidopsis [33]. Posttranslational modifications contribute to the native structure, and may enhance the binding capacity of the peptide ligand to its receptor. In some studies, the lack of posttranslational modification has been shown to reduce the peptide activity [13,34,35]. Posttranslational modifications are thus, probably necessary for full bioactivity and downstream signaling [13,34,35].
The mechanisms by which plant-secreted peptides are produced are not well understood, but are thought to be somewhat similar to the maturation process of their mammalian counterparts. In general, signaling peptides are translated as large precursors (so called pre-propeptides) followed by one or more posttranslational modifications and/or proteolytic processing. Based on what is known from the study of animal neuropeptides, the maturation of signaling peptides is assumed to start by the co-translational removal of the N-terminal signal peptide by Signal Peptide Peptidases (SPPs) in the endoplasmic reticulum to yield the propeptide [12,13,36]. The cleavage site in the precursor can be predicted with a high degree of accuracy [37]. The resulting propeptide is likely to be directed through the Golgi apparatus and, together with processing proteases, loaded into newly formed secretory vesicles. Later on, as the vesicle matures, proteolytic processing occurs with removal of the variable region and release of the mature peptide contained in the C-terminal region (some cysteine-rich peptides do not require this proteolysis step). Finally, mature peptides will be released to the extracellular space and will bind to the cognate receptor(s) in the plasma membrane of neighboring cells, or more distant cells if the peptides are somehow channeled away, where they trigger a signaling pathway that initiates specific physiological responses [12].
Although the overall pathways involved in the production of signaling peptides in plants and animals are most probably similar, some details in these processes are still uncharacterized. Plant peptides are generally small and not functional when associated with large tags. Their intracellular localization has thus not yet been achieved, and in plants, the detection of peptide-carrying vesicles is still pending. Furthermore, in mammals, neuropeptides and peptide hormones are produced by specialized cells, such as neurons and neuroendocrine cells, and in certain cases stored in mature vesicles releasing the peptide signal when triggered by a stimulus. In plants, peptide production does not seem to be confined to certain cells or organs and any plant cell type appears to be able to deliver a peptide signal. Also, it is unclear whether plant signaling peptides are pre-packed and stored, or synthesized on demand. Only future insights will tell us to what extent peptide production and release have diverged between vertebrates and higher plants.
Membrane-localized receptors function as master switches of complex intracellular signaling networks. Receptors are activated upon binding of their ligand, often through autophosphorylation. Activated receptors will, in turn, phosphorylate downstream targets, which might result in a direct response and/or a phosphorylation cascade, including mitogen-activated protein (MAP)-kinases, and eventually lead to transcriptional regulation of specific genes [12,13,36,38].
A bottleneck in the functional study of signaling peptides in plant growth and development has been the identification of the encoding genes. Genome sequencing of different plant species has led to the prediction and identification of a number of signaling molecules. However, because the precursors are rarely more than 120 amino acids long, genes encoding small peptides tend to be overlooked in genome annotations because they are difficult to distinguish from short, random open reading frames. Additionally, the detection of mature small signaling peptides in crude plant tissue extracts is also a difficult task given their very low physiological concentration (in the nanomolar range) and the presence in large amount of other peptides derived from the degradation of abundant proteins. It is therefore likely that only a few of the total number of secreted peptides are known to date. Thus, attempts to identify new signaling peptide families in plants by various approaches are still highly valuable [4,39,40]. They can be classified in three main types of approaches: genetic, bioinformatic and biochemical assays.
Forward and reverse genetic approaches have been used as tools to identify new signaling peptides. For example, the clavata3 mutants were identified because they resulted in enlarged Shoot Apical Meristems (SAM) in a forward genetic mutant screen [41]. Thanks to similar forward genetic approaches, several other signaling peptides were identified, such as INFLORESCENCE DEFICIENT IN ABSCISSION (IDA) and TAPETUM DETERMINANT1 (TPD1) [7,42]. Reverse genetics in combination with bioinformatics tools resulted in the identification and characterization of the GOLVEN/Root Growth Factor/CLE-Like (GLV/RGF/CLEL) signaling peptide family [9,10,22]. However genetic studies have been hampered by two major hurdles: firstly, because signaling peptide genes are small, corresponding T-DNA insertion alleles are often not available; secondly, signaling peptides are usually encoded in functionally redundant multigene families impeding forward genetic screens.
Several successful examples of the discovery of signaling peptides by means of in silico approaches have been reported. Typically, several criteria are combined to search for new signaling peptide families: (i) the presence of a signal peptide sequence that would target the propeptide to the secretory pathway (for example using the SignalP algorithm, [37], (ii) the small size of the precursors, (iii) the enrichment or lack of cysteine residues in the propeptide, and (iv) the presence of sites susceptible to posttranslational modification(s). Application of one or more of the abovementioned features for in silico genome sequence searches led to the identification of multiple plant signaling peptide families including GLV/RGF/CLEL, C-TERMINALLY ENCODED PEPTIDE (CEP), IDA and CLAVATA3/ESR-RELATED (CLE) members [4,7,9,10,22,29].
Despite their usefulness, bioinformatics approaches have limitations because many small genes remain poorly annotated and homology searches may not be particularly useful for the identification of functional homologs that only share a few amino acids.
Assays have been developed to test for the presence of bioactive compounds in crude extracts, followed by the purification of the responsible secreted peptides. For example, medium alkalinization assays led to the identification of the RAPID ALKALINIZATION FACTOR (RALF) from tobacco and Phytosulfokine (PSK) was discovered in conditioned media because it promoted the proliferation of cultured cells [6,24].
Reversely, once candidate signaling peptides have been identified via genetic or bioinformatics studies, they may be chemically synthesized and applied to plant tissues or organs to analyze their action in vivo. This strategy creates opportunities to investigate the output of the peptide signaling pathway and to complement a genetic loss-of-function. But it also suffers from several limitations: peptides may act specifically in a small concentration range or in specific locations that may not be reflected when exogenously applied. Finally, the structure of the native signal, possibly including complex posttranslational modifications, may be insufficiently characterized or difficult to copy via chemical synthesis.
We anticipated that these approaches, on their own or combined, will accelerate the discovery of novel plant signaling peptides in the near future.
Signaling peptides are involved in different biological processes
Signaling peptides have been associated to diverse developmental processes in plants, such as apical meristem maintenance (CLAVATA3 (CLV3), GLV/RGF/CLEL), defense (systemins), floral organ abscission (IDA), and stomatal patterning (EPF), among others [4,38,43-46]. To highlight the diverse roles of signaling peptides in various developmental processes, some examples of the function of the signaling peptide families are outlined below.
CLAVATA3/ESR-Related (CLE)
The Arabidopsis CLE peptide family was originally identified through the club-shaped fruit phenotype of some of the corresponding mutants (“clava” Latin for club) [41]. Further investigations revealed that the CLV3 gene is exclusively expressed in a limited number of cells in the SAM and restrains stem cell division in the SAM [5,28,47] (Figure 2). The clv3 mutants exhibit overproliferation of stem cells, resulting in a meristem almost 1,000 fold larger than that of the wild type. Overproduction of CLV3 causes loss of stem cell activity and differentiation of the shoot meristem [48]. After binding to the leucine-rich-repeat-receptor like kinase (LRR-RLK) CLV1, the CLV3 peptide promotes the repression of the homeodomain transcription factor WUSCHEL (WUS) and, through a negative feedback loop, regulates SAM homeostasis. In this model, WUS binds directly to the CLV3 promoter and activates its transcription. The CLV3 peptide suppresses WUS expression with decreased CLV3 transcription as a result, thereby reducing the number of stem cells [4,5,48-52].
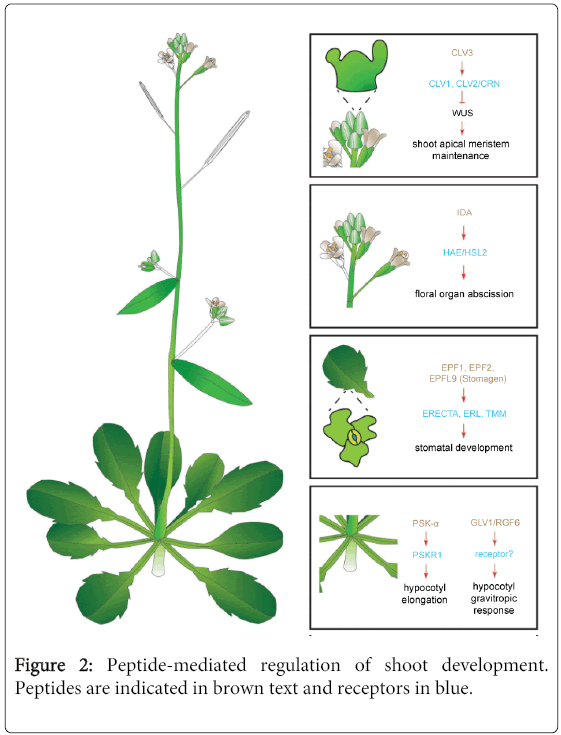
Figure 2: Peptide-mediated regulation of shoot development. Peptides are indicated in brown text and receptors in blue.
Additionally, CLV2 and CORYNE (CRN) have been shown to form a receptor complex contributing to SAM maintenance. However, CRN lacks kinase activity and no direct CLV3-CLV2/CRN binding has been demonstrated. Possibly, this receptor complex relays the CLV3 signal independently from CLV1 [53,54]. Interestingly, a clavata-like phenotype in maize has recently led to the discovery of a downstream component in this signaling pathway [55]. COMPACT PLANT2 (CT2) encodes the predicted a-subunit (Ga) of a heterotrimeric GTP-binding protein. The maize orthologue of CLV2, FASCIATED EAR2 (FEA2), codes for a receptor without a signaling domain. FEA2 interacts with CT2 to transmit CLAVATA-dependent signals to regulate the SAM maintenance. However, genetic evidence suggested that FEA2 also signals through other pathways besides CT2/Ga [55].
A molecular mechanism controlling stem cell activity, similar to the SAM CLV-WUS pathway, has been proposed in the root apical meristem (RAM) (Figure 3). In the RAM, it is the binding of the signaling peptide CLE40 to CLV1 that would repress, together with the receptor-like kinase ARABIDOPSIS CRINKLY4 (ACR4), the expression of the transcription factor WUSCHEL-related homeobox 5 (WOX5). WOX5 controls the communication between the quiescent center (QC) and the surrounding stem cells and promotes the stem cell identity in these cells. Plants overexpressing CLE40 display a stunted primary root growth. It is thought that the CLE40-ACR4/CLV1 signaling restrains the production of WOX5 to the QC, thereby restricting the stem cell identity to the cells in contact with the QC cells. According to this model, increased levels of CLE40 prevents WOX5 to reach the neighboring cells that ultimately lose their stem cell nature and differentiate. In cle40 and acr4 mutants, the inhibition is lost and the WOX5 expression domain is expanded resulting in the overproliferation of stem cells [56-58] (Figure 3).
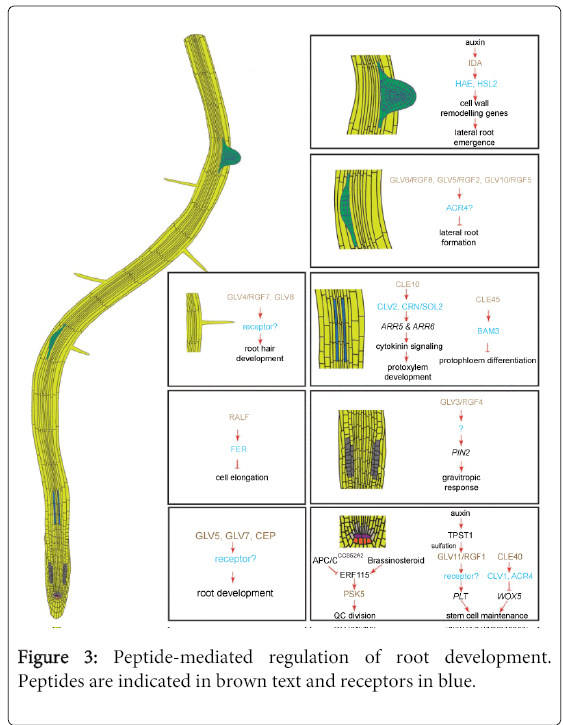
Figure 3: Peptide-mediated regulation of root development. Peptides are indicated in brown text and receptors in blue.
The role of plant hormones, transcription factors and microRNAs signals is well established in vasculature development. Recent studies have also highlighted the action of signaling peptides in this context. The TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF) peptide stimulates procambial cell proliferation in the leaf and hypocotyl vasculature and at the same time constrains the differentiation of procambial cells into tracheary elements. TDIF was isolated first from mesophyll cell cultures of Zinnia elegans in which the tracheary element differentiation was arrested. Later investigation led to the isolation of an extracellular 12-amino acid peptide and a homology search revealed that TDIF was identical to the C-terminal motif of CLE41 and CLE44 and highly homologous to CLE42 and CLE46 in Arabidopsis [27,59]. Accordingly, cle41 and cle44 mutants show a reduced number of procambial cells and exogenous application of TDIF also increases procambial cell proliferation in Arabidopsis hypocotyls [27,60-62]. The TDIF RECEPTOR/PHLOEM INTERCALATED WITH XYLEM (TDR/PXY) gene was identified as coding for an LRR-RLK receptor potentially involved in TDIF recognition. TDR/PXY is mainly expressed in procambium and cambium cells, whereas CLE41 is transcribed in neighboring phloem cells, indicating that CLE41 must be secreted towards the procambium where it is perceived by TDR/PXY [60,62,63]. Intriguingly, another WUS-related homeobox gene, WOX4, is essential for the regulatory function of TDIF in the vascular cell fate, but not for TDIF inhibition of xylem differentiation [64,65]. The signaling peptides CLE9 and CLE10 have also been reported to be produced in the vasculature and to function in vascular development. CLE10 overexpression results in severe inhibition of the protoxylem formation in roots. The CLE9/CLE10 peptides are assumed to signal through the CLV2 receptor to inhibit protoxylem formation in roots via cytokinin signaling. Accordingly, protoxylem formation is no longer inhibited by CLE9/CLE10 peptides in clv2 mutants, indicating a link between the CLE9/CLE10 ligand and the CLV2 receptor in vascular development [66] (Figure 3). Another CLE peptide, CLE45, was recently reported to be involved in protophloem differentiation. Recent studies have suggested that CLE45 might interact directly or indirectly with the leucine-rich repeat receptor-like kinase BARELY ANY MERISTEM 3 (BAM3) to negatively regulate protophloem differentiation. CLE45 and BAM3 are both expressed along the developing protophloem, up to the beginning of the elongation zone, and are proposed to function in the transition from proliferation to differentiation [67].
Abscission is one of the developmental processes in plants that are coordinated by signaling peptides. Abscission is essential for optimal plant growth, because organs that are not necessary or functional anymore have to be removed through precise and programmed cell separation [68-70]. IDA and IDA-LIKE (IDL) signaling peptides have been shown to promote organ abscission by stimulating cell separation or by inhibiting cellular repair mechanisms [7,70] (Figure 2). The IDA and IDL genes are expressed in the cell separation zone that comprises the region where the organs are detached from the plant body [70]. It is believed that they signal through the LRR-RLKs HAESA (HAE) and HAESA-LIKE2 (HSL2) [70]. The ida knockout mutants retain floral organs, whereas plants overexpressing the IDA or IDL genes exhibit premature floral organ abscission, with overproliferation of the abscission zone and, additionally, ectopic abscission of some organs that are normally not shed in Arabidopsis [68,69,71].
IDA signaling peptides are mostly known for their role in floral organ abscission [7] but, recently, a new function has been assigned to the IDA-HAE/HSL2 signaling module, namely a contribution to facilitate the passage of Lateral Root Primordia (LRP) through the main root and to assist in the Lateral Root (LR) emergence. In roots, IDA is strongly and continuously induced by auxin, whereas the hormone only transiently upregulates HAE and HSL2. Auxin induction of IDA depends on the auxin influx carrier LIKE AUX1-3 (LAX3) and the AUXIN RESPONSIVE FACTOR7 (ARF7). Auxin, originating from the tip of the primordium, coordinates cell separation in overlaying LRP tissues through signaling mediated by the IDA peptide. IDA and HAE have been shown to regulate LR emergence by promoting the degradation of the cell wall in the tissue layers overlaying the LRP as it grows outward [71] (Figure 3).
Stomatal development relies on asymmetric cell divisions that are precisely coordinated in time and space through cell-to-cell communication networks. Stomata are usually separated from neighboring stomata by at least one cell, following the so-called “one-cell-spacing rule”. In this context as well, signaling peptides have a significant impact on development [44,72-74]. Multiple cysteine-rich signaling peptides that belong to the EPF family, in particular EPF1, EPF2, EPF-LIKE6/CHALLAH (EPFL6/CHAL) and EPFL9/STOMAGEN, are associated with the regulation of stomatal density and positioning [18,19,44,72,75-78]. Overexpression of EPF1 and EPF2 reduces stomatal density. Signaling depending on EPF1 and EPF2 requires the activity of the LRR-receptor-like protein TOO MANY MOUTHS (TMM) and the LRR-RLKs, ERECTA (ER) and ERECTA-LIKE1 (ERL1). EPF1 and EPF2 bind to the ER and ERL1 receptors and EPF2 to TMM. As TMM lacks an intracellular domain [79], another protein with an extracellular domain is required to mediate the signal transduction. Therefore, TMM interacts directly with the ER receptor and forms a complex that initiates the EPF signaling pathway [18,19,78] (Figure 2).
Interestingly EPFL9/STOMAGEN, another member of the EPF family, acts antagonistically to EPF1 and EPF2 in stomatal development. EPFL9/STOMAGEN also requires TMM but, in this case, to positively control stomata formation, indicating that peptide hormones from the same family can have opposite functions through the same receptor in planta [20,80]. Lastly, the EPFL6/CHAL also negatively controls stomatal development in the epidermis of stems and hypocotyls. The ER receptors have been proposed to be involved in the transmission of the EPFL6/CHAL signal [44,75,77].
Recently, a function distinct from stomatal development has been assigned to the signaling module EPF4/EPF6-ER/ERL [81]. EPF4 and EPF6 may be secreted from the endodermis and perceived by ER/ERL1 in the phloem to regulate vascular development [82].
Three independent in silico studies identified a novel signaling peptide family that was designated GOLVEN (GLV), Root Growth Factor (RGF), or CLE-Like (CLEL). As the three research groups used different sequence homology parameters, not all the family members were initially identified by all groups. Overall, the GLV/RGF/CLEL gene family counts 11 members in Arabidopsis [9,10,22]. Although GLV/RGF genes had originally been detected in Arabidopsis they are conserved in all higher plants analyzed so far [10]. Like most other secreted signaling peptides, they code for posttranslationally modified small peptides. The GLV precursor proteins exhibit a typical tripartite structure [83]. The sequence of the native secreted peptide has been dissected for four family members, namely GLV1/RGF6, GLV2/RGF9, GLV3/RGF4 and GLV11/RGF1, which are 14, 15, 18 and 13 amino acids in length, respectively, and carry at least two types of posttranslational modifications in their mature form: tyrosine sulfation and proline hydroxylation. Tyrosine sulfation increases the bioactivity of the mature peptide, but hydroxylation of the proline residue has not been associated with any functional role so far [9,10].
Collectively, the GLV/RGF genes are transcribed in all plant organs, in both vegetative and reproductive stages. Nevertheless, individual transcription patterns are highly specific and are restricted to a few cells or cell types. The first loss-of-function phenotype reported for GLV genes was a short-root phenotype that was also been observed in the tpst1 mutant [9]. TPST is the only identified enzyme that catalyzes tyrosine sulfation in Arabidopsis [25]. The tpst short-root phenotype could not be complemented by the addition of the known tyrosine-sulfated signaling peptides, PSKs and PLANT PEPTIDE CONTAINING SULFATED TYROSINE1 (PSY1). This observation implied that distinct tyrosine sulfated signaling peptides in Arabidopsis were involved in the positive regulation of root growth and lead to the discovery of the RGF peptide family [9,84]. Although single rgf1/glv11, rgf2/glv5, and rgf3/glv7 loss-of-function mutants did not exhibit any obvious root phenotype, the rgf1 rgf2 rgf3 triple mutant plants have short roots with a decreased number of meristematic cortical cells. Addition of a RGF1/GLV11 synthetic peptide in the growth medium restored the meristem size of both the tpst and rgf1 rgf2 rgf3 mutants. GLV/RGF peptides coordinate the postembryonic maintenance of the root meristem cell niche through defined mRNA and protein levels of the PLETHORA (PLT) transcription factors [9] (Figure 3). However, overexpression and exogenous application of all GLV peptides had not the same effect on root growth, suggesting that not all members of the family are involved in root meristem maintenance [9,10,46] (Figure 3).
All GLV/RGF genes, except GLV1/RGF6, are transcribed during LR development. Furthermore, overexpression of some GLV genes strongly decrease the number of emerged LRs and treatment with some GLV/RGF synthetic peptides has similar effects on wild-type plants. Despite the lack of information on the LR phenotype in GLV/RGF loss-of-function mutants, accumulating evidence suggests the involvement of the GLV/RGF genes in LR initiation and development [22,46].
A striking curly root phenotype resulting from overexpression of the GLV/RGF genes revealed a novel function that differed from root growth regulation. Referring to this phenotype, Whitford et al. [10] designated this family GOLVEN (GLV), which means ‘waves’ in Dutch because the overexpression plants do not form the regular wavy-shaped roots normally observed when grown on the surface of impenetrable inclined agar medium. This observation suggested a possible role for the GLV/RGF genes in gravitropic responses. Moreover, rotation of vertically grown GLV3/RGF4 gain- or loss-of-function mutant seedlings revealed that the root response to gravistimulation was affected in both cases. GLV3/RGF4 gain-of-function plants display partially agravitropic roots, whereas the gravitropic response of amiRglv3 seedlings was enhanced. Addition of the corresponding synthetic peptide mimicked the gain-of-function phenotype. Plants overexpressing the GLV1/RGF6, GLV2/RGF9 or GLV3/RGF4 genes have hypocotyls with an altered gravitropic phenotype reminiscent of the root gain-of-function phenotype.
The phenotype observed in GLV/RGF-overexpressing roots resembles that of mutants affected in either auxin response or transport. Further analysis pointed out that GLV3/RGF4 is involved in the root gravitropic response by influencing PIN-FORMED 2 (PIN2) trafficking. Differential turnover of the auxin efflux carrier PIN2 between the upper and lower side of the gravistimulated root mediates the formation of the lateral auxin gradients by which plants can respond to gravity [85]. Overexpression or treatment with GLV/RGF peptides hampers PIN2 trafficking in the root tip, hence preventing the proper formation or maintenance of the required auxin gradient. Most likely, a regulatory mechanism involving the GLV/RGF signal increases the robustness of the established auxin gradient. However, the molecular mechanism by which the signal controls PIN2 trafficking is still unknown [10].
Contrarily to the other Arabidopsis GLV genes, GLV4/RGF7 and GLV8 are not expressed in the root tips and are thus expected to be involved in other root developmental process(es) than root meristem homeostasis and LR development. The ectopic overexpression of GLV8 caused the production of root hairs with more complex and irregular shapes, whereas the glv8 loss-of-function mutant harbors shorter root hairs with a simpler structure than the wild type. GLV4/RGF silencing also resulted in root hairs that were shorter than those of the wild type, confirming the possible role of these GLV/RGF genes in root hair development [46] (Figure 3).
Phytosulfokines (PSK) and Plant Peptide Containing Sulfated Tyrosine (PSY) sulfated peptides
As mentioned above, TPST is responsible for tyrosine sulfation in Arabidopsis, tpst knockout mutants display severe root and shoot phenotypes, and several sulfated peptides are known to be involved in root growth and regulation, in addition to GLV/RGF/CLEL. PSK-a and PSY1 are thought to control root growth positively by promoting the mature cell size [9,45,86,87].
The sulfated signaling peptide, PSK-a positively regulates cell expansion and hypocotyl length in Arabidopsis. Through the PSKR1 receptor, the PSK-a signal promotes hypocotyl elongation and protoplast expansion. tpst knockout mutants displayed severe phenotypes, including shortening of the hypocotyl that was only partially restored by supplementing the PSK-a peptide. This observation implies that, besides PSK-a, likely one or more additional sulfated peptides contribute to the regulation of hypocotyl elongation [88].
Finally, a separate signaling pathway has been proposed recently that controls the division rate of the QC cells via PSK5 [89]. In this pathway, the transcription factor ETHYLENE RESPONSIVE FACTOR 115 (ERF115) binds to the promoter region of PSK5, thereby activating its expression. Upstream, brassinosteroids positively regulate ERF115-PSK5, whereas the CELL CYCLE SWITCH 52A2 subunit of the ANAPHASE-PROMOTING COMPLEX/CYCLOSOME (ACP/CCCS52A2) reduces ERF115 abundance through proteolysis [89] (Figure 3).
Rapid Alkalinization Factor (RALF)
A RALF peptide was first isolated from tobacco leaves as a molecule that triggers fast and strong pH hikes in tobacco suspension cell cultures, that stimulates a mitogen-activated protein kinase (MAPK), and that arrests root growth [6]. The cysteine-rich RALF-coding genes are found throughout the plant kingdom and Arabidopsis counts 34 RALF homologues [39]. The RALF and RALF-like (RAFL) genes are expressed in most tissues including roots, shoots, leaves and flowers, and are suspected to regulate diverse developmental modules, including root development [6,90]. Similar to most of the signaling peptides, RALFs possess a N-terminal signal peptide indicating they are secreted [6] and conserved dibasic sites that could be targeted for subtilase proteolysis to generate the mature peptides.
AtRALF23 is downregulated by Brassinosteroids (BR) and is thought to be a negative regulator of BR-mediated growth-promoting effects [91,92]. AtRALF23 overproduction phenotypes – dwarf bushy plants – were shown to be suppressed in the sbt6.1 subtilase null mutant background and no processed AtRALF23 peptide could be detected in these plants [93]. The plant-purified AtSBT6.1 proteolytic enzyme was demonstrated to process the AtRALF23 in vitro, thereby confirming that the propeptide is a direct target of the subtilase [93].
C-Terminally Encoded Peptide (CEP)
Small posttranslationally modified CEP peptides display diverse functions in various plant tissues [23,29]. Synthetic CEP1 peptide treatment, as well as CEP gene overexpression, resulted in root growth inhibition through repression of meristematic cell division and expansion of the cells in the elongation zone, without interfering with the QC specification. Surprisingly, CEP1 expression was not detected in the RAM, suggesting a nonspecific function of CEP1 in RAM development. In addition, CEP1 is expressed in lateral root primordia, implying a yet to be defined role of CEP1 in lateral root development [29]. Finally CEP5 may be involved in shoot development but further research is required to identify its precise mode of action [23].
Embryo Surrounding Factor1 (EFS1)
An additional cysteine rich signaling peptide family was identified in a recent study that regulates early embryo patterning in flowering plants. EMBRYO SURROUNDING FACTOR1 (EFS1) is derived from the central cell gametes before fertilization and from micropylar embryo-surrounding endosperm cells after fertilization. It regulates the apical-basal development of the neighboring embryo through the receptor-like kinase SHORT SUSPENSOR and the mitogen-activated protein kinase YODA [94].
In the complete genome sequence of Arabidopsis, more than 1,000 genes have been found that encode putative secreted peptides with a potential signaling function [29,40]. In accordance to these studies, new signaling peptides are being identified and assigned to different plant growth and developmental functions, potentially leading to the discovery of unsuspected developmental programs. In parallel, as more information accrues about signaling peptides, additional structural hallmarks will likely result in the identification of new families.
Many aspects remain still to be elucidated concerning signaling peptides in plants. First, little is known about the details of their biosynthesis. Only a few proteases involved in the processing of the precursors have been reported so far [92,93,95] and, to date, a general picture cannot be drawn that explains how the mature peptides are produced in plant tissues. Their small size has hampered their intracellular localization and the route from the initial translation of the precursor to the secretion of the mature peptide is still largely hypothetical. Future work should be oriented towards a better understanding of the mechanisms through which they are produced.
Signaling peptides control diverse processes related to plant physiology, growth, and development. However, we are just starting to uncover the pathways that they trigger. In most cases, the receptors they bind to and the downstream targets are unknown. Ongoing forward and reverse genetic screens will certainly provide some insights on these in the near future. Alternatively, the characterization of changes in the phosphorylation status or the structure of receptors upon binding with their peptide ligand will also yield valuable information. For example, the crystal structure of the flagellin-sensitive 2 (FLS2) and BRASSINOSTEROID INSENSITIVE 1-associated kinase 1 (BAK1) ectodomains was recently resolved as a complex with the bacterial flagellin fragment peptide, flg22 [96].
Finally, another level of complexity has to be taken into consideration. Converging evidence indicates that the signaling peptide and phytohormone pathways are interconnected [10,66,97]. Future studies will have to dissect the cross-talks so we better understand how these molecular signals interact to deliver the messages necessary to shape the plant.
Work in the lab of T.B. was in part financed by grants of the Interuniversity Attraction Poles Programme from the Belgian Federal Science Policy Office, the Research Foundation Flanders (FWO) and by the special research fund of University Ghent.