Journal of Geology & Geophysics
Open Access
ISSN: 2381-8719
ISSN: 2381-8719
Research Article - (2014) Volume 3, Issue 6
This article is a short, theoretical analysis exploring the possibility of convergences of (similar) organs at identical morphologic positions in different organisms. The latter criterion normally characterizes homology, and is an important character in understanding systematic relations. Homology, however, depends on a shared latest ancestor in common, which in fossils cannot always be proved. We illustrate that organs similar in structure and function may have found different ways along a time axis and in space to have appeared independently in the same morphological position, and although being convergent, they are indistinguishable from homologies.
Keywords: Cladistics; Homology; Cambrian; Ecological niche; Convergence; Phylogeny
There exist several formulations for describing “Homology“, and Homology is an important tool for assessing phylogenetic relationships, especially in modern cladistic analyses in palaeontology. Correlating morphological patterns of homologous organs contrasts greatly with sequence analysis of genes, where genetic pathways and their mutations are pursued as part of the process of establishing phylogenetic trees. The history of “Homology“, of course, relates to the understanding of the origin of species. ομολογειν from ancient Greek means “to coincide“, and in pre-Darwinian times, organs were considered as homologous, if they had a similar position in the context of morphology or ontogenetic development, whatever their form and function was [1]. A fine example of such a “homology“ is the position of the premaxillary in the skull of humans in comparison with other mammals, which became important for early discussions of any possibility of evolution. J. W. von Goethe in 1790 [2] believed that he had found this bone, well known from other mammals, in the human embryo, together with J.C. Loder in Jena in 1784. Neither was aware that it had been discovered independently by F.Vicq d’Azyr a short time before [3,4]. At the beginning of the 19th century Georges Cuvier, a pioneer of Palaeontology, started to order fossils and recent animals using homologous organs, in this sense, to create trees showing affinities, and Richard Owen in 1843 defined “Homology“ as “the same organ in different animals under every variety of form and function”.
When Charles Darwin discovered the mutability of species [5], it became necessary to change the understanding and thus the definition of the term “Homology“. Since “Homology“ thereafter had to be seen strictly in an evolutionary context: organs in two species could be homologous only if the same structure was present in their last common ancestor. The fact that recent structures of at least two organisms are based on an earlier structure shared with the last ancestor in common, known as “synapomorphy“ is thus a precondition for homology in the modern sense. The basic trait they derive from is the “plesiomorphic organ“ (Figure 1).
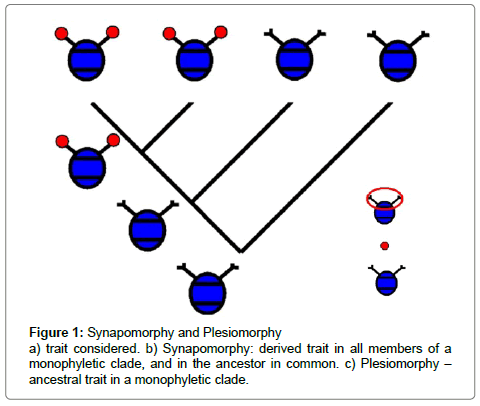
Figure 1: Synapomorphy and Plesiomorphy a) trait considered. b) Synapomorphy: derived trait in all members of a monophyletic clade, and in the ancestor in common. c) Plesiomorphy – ancestral trait in a monophyletic clade.
In palaeontological research, how exactly this point is to be securely fixed can be problematic, as will be pointed out later. It is easy to define homology in this sense, if the most recent structure under consideration, in an evolutionary sense, can be traced backwards through intermediate forms to that of an earlier, and differently formed ancestor. Thus the hoof of modern horses (Equus) is homologous to the third toe on the foot of of the Eocene Hyracotherium. This situation, as a criterion for establishing homology was formulated by A. Remane (1952, “Criterion of continuity“), and can be deduced by the definition mentioned above [6].
The opposite to homology is analogy, where the organs being considered do not have a common ancestor, but show the same function. The animals that bear them may belong to related or quite unrelated groups, and the organs may be situated at morphologically different positions. Analogous organs, for example are the “digging hand“ of a mole (Talpa europea) and a European mole cricket (Gryllotalpa gryllotalpa), or the wings of a bat, a bird and a butterfly. Such analogues develop, if organisms, often of different taxa, in different areas or even continents, or different geological periods, invade the same ecological niche. Such processes are known as convergences (Figure 2). Referring to Moore and Willmer [7] or Conway Morris [8], identical ecological niches offer identical constraints of life and enforce an optimal body plan which enables its possessors to survive most efficiently.
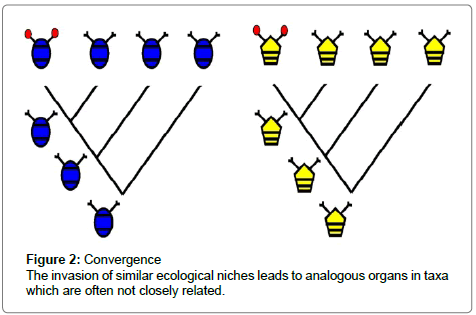
Figure 2: Convergence The invasion of similar ecological niches leads to analogous organs in taxa which are often not closely related.
Homologous organs may have very different functions, and it is only their origin that is relevant for systematic discussion (Figure 3). Critical in this contexts are those homologous organs of uncertain history which look rather similar to each other, because it is easy to mistake these for convergences. Synapomorphic organs with different forms (and functions) however may be challenged as strictly homologous by the genes controlling them.
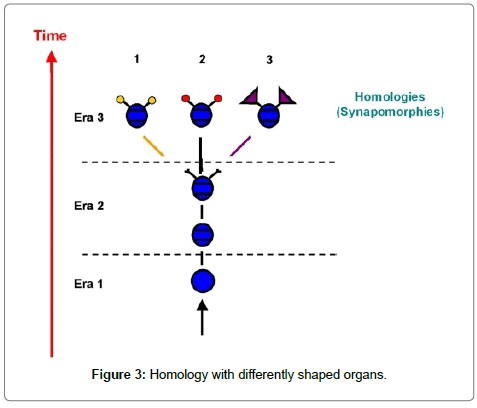
Figure 3: Homology with differently shaped organs.
As mentioned before the term “Homology“ is the basis for many cladistc analyses. Cladistic methodology (in Biology or Palaeontology) is an appproach intended to establish the evolutionary context of living and fossil organisms. Such cladistic classifications are based on decisions as to whether (or not) one or more traits of the organisms under consideration are shared with the group´s latest common ancestor. Thus, cladistic definitions are based on synapomorphies, while these synapomorphies are characters shared by more than one organism descending from one ancestor in common as mentioned before (Figures 2 and 3). Within the phylogenetic trees each synapomorphy becomes plesiomorphic to the next descendant, with exception of the last synapomorphy.
Cladistic analysis since it was founded by Willi Henning in the 1960s has become established as a widely used method, especially for establishing the systematic relationships of fossil and living organisms, and the results are often interpreted in a phylogenetic context. The cladistic approach, however, especially the use of synapomorphies as a safe criterion, has been challenged in great detail (e.g. and overview see [9]). Whereas the use of molecular systematics, in this context, has indisputable advantages over morphological criteria. Such information is not generally available in fossils [7].
One of the main critical points in considering cladistic analyses is the lack of any time axis. This is most important at the beginning of the fossil record of hard shelled invertebrates, in other words the Lower Cambrian, where for many organisms no obvious predecessors exist. This leads, inevitably, for example in the realm of arthropods, to the concept of “stem arthropods“, as ancestors of subsequent lines of descent. Taking these stem arthropods as a starting point of subsequent evolution, heterochrony may be a further source of confusion in interpreting phylogeny – an apparently new, synapomorphic structure may be present in the early stages of development of an ancestor, to achieve expression in the adult of the descendant through paeodomorphosis. Moreover, it seems, that some organs such as eyes develop, in an evolutionary sense, much faster than others, appendages for example.
The intention of this short article is not to give an overview of the history of discussion about the value of cladistics, nor to discuss and disentangle the multitude of terms which have been differentiated meanwhile, nor is intended to be a challenge to particular papers or authors. The intention is, rather, to illustrate in a brief systems analysis, what happens to a phylogenetic system based on cladistic analysis, if the homologous organs are reasonably similar, an ancestor in common is not present, but only assumed, and when the identical morphological position of any particular organ is overrated in its relevance. The main problem, as always, is how may the latest common ancestor be identified onjectively. In many cladistic tables, it is noted: only trait x in the morphological position yz is present (1) or not present (0).
As cladistic analyses lack any time axis, we should consider here a brief systems analysis of what kinds of convergence may appear over space and time, and which of them are difficult to distinguish from homologies, and thus create problems in establishing systematic relationships.
Thus a clear definition of both terms is necessary. We consider all synapomorphies as homologous; all being derived traits in all members of a monophyletic clade, with one ancestor in common (Figure 3). Morphology and function may be different in different homologous organs, while the position in the morphological context is the same for all homologous traits. Analogies are the result of a convergent evolution. Within a certain limit of morphological similarity, organs develop into a functionally similar system, while structural details, such as the position in the body plan can vary. Often analogous organs develop in different morphological positions (Figure 4 position 1 vs 2-3). In any discussion about the homology of such convergences which emerge in the identical positions of the body plans of different organisms, and indeed whether they exist, become relevant. This is particularly the case at the base of possible phylogenetic trees, in other words, in the Cambrian. Such a convergence along a time axis may be called “Vertical Convergence“:
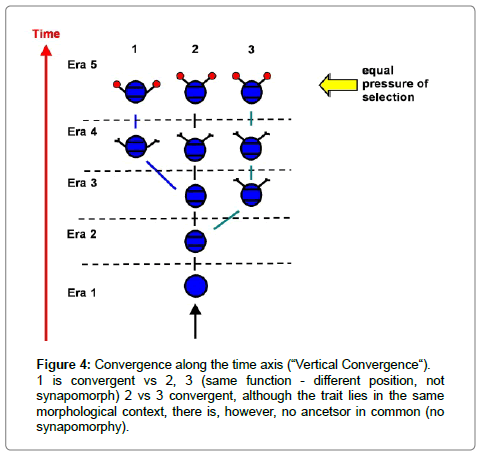
Figure 4: Convergence along the time axis (“Vertical Convergence“). 1 is convergent vs 2, 3 (same function - different position, not synapomorph) 2 vs 3 convergent, although the trait lies in the same morphological context, there is, however, no ancetsor in common (no synapomorphy).
“Vertical Convergence“ takes place when at different times, throughout the geological record convergent developments result from similar selection pressures, as for example, the result of climatic changes which have happened often during the history of the Earth. In relation to homology these become problematic, if they appear at the same morphological positions, especially if they did so in organisms which belong to the same systematic groups.
Classic cases of convergence such as the examples mentioned before, appear in parallel in the same time interval, contemporarily, thus here we are considering time “horizontally“. They are separated by spatial or geographical mechanisms of isolation (continents, mountain ranges, etc). Naturally, both vertical and horiozontal convergences may take place within an evolving lineage. If such horizontal and vertical convergence both occur this may be defined as “Cross Convergence“, which is illustrated in Figure 5. It may indeed be the most common type.
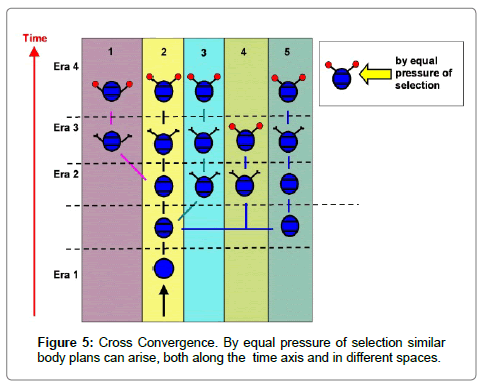
Figure 5: Cross Convergence. By equal pressure of selection similar body plans can arise, both along the time axis and in different spaces.
Differentiated vertical convergence, in a spatial context is shown in Figure 5. All identical traits are not homologous but convergent, even when situated in the same morphologic position, because none of them have latest ancestors in common. They are separated by time and/or space. In fossils these similar emergents may not be distinguishable from homologies without additional information, such as the origin of the fossils, and their geologic history etc. (If convergent characters developed at different morphologic positions (1), then convergence cannot be confused with homology).
A more complicated case of vertical convergence, would be the result of a “weaker“ gene, prone to mutating more readily than others, (and with additional equal selection pressure such as a climate change) in such a case a character may develop independently, to be (more or less) identical at several times during geological history, and result in a convergent trait. Again, these traits were not homologous, because they are not synapomorphic, as shown in Figure 5.
An almost classic example of such convergences is the one of ground scrabblers, which have developed again a “digging-hand“-morphology. The European mole (Talpa europaea) systematically belongs to the insectivora. It digs with its hand into the ground; its hand has five digits and an additional thumb, which arises by transformation of a sesamoic bone of the wrist. The Cape golden mole (Chyrsochloris asiatica) is also an insectivore, it digs in the same way as the European mole with its hands, but the “digging instruments“ are just the claws of its hands. The Cape golden mole lives in South Africa. Finally the Australian marsupial mole (Notoryctes typhlops) is belongs to the Marsupalia, digging, as does Chrysochloris with its claws. The insectivora have an ancestor in common, their organs are homologous, the marsupalian “mole“ does not share this predecessor and their digging hand is a convergently evolved structure, although it is almost identical to the latter, and at the same morphological position. All organisms cited here live in the same geological age, in other words today, but they inhabit different continents. Isolated from each other, they invaded the same ecological niche, and thus their overall morphology and digging hands represent a classic example of convergence. Regarding time (all live in one time interval) we have a horizontal parallel deveopment of functionally identical forms in different geographical areas. Regarding the European mole and the Australian marsupalian, we are here considering a horizontal convergence, which has developed at different spaces (continents). As regards the insectivores, which show the same morphological structure for digging, in other words their claws, we may have a vertical convergence, because is seems to have developed independently at different geological eras, But surely this is also a horizontal convergence, because both live at different continents. As a result it may be a cross convergence, which, if the claws alone were considered, and the last ancestor in common was not known, would be indistinguishable from a homology. There are many similar examples in fossils; one being the repeated morphotypes in trilobites defined by Fortey & Owens [10], where unrelated forms have remarkably similar overall morphology. These authors defined eight such convergent trends; (i) pelagic trilobites with large eyes, (ii) illaenimorphs, where the dorsal furrows are largely effaceed, (iii) trilobites with marginal cephalic spines, (iv) olenimorphs – flat, thin shelled forms with many segments; adapted for low oxygen conditions (v) trilobites with pitted fringes (vi) miniaturised trilobites (vii) atheloptic trilobites, which lived in deep waters and have reduced or absent eyes (viii) phacomorphs – convex, thick - shelled tuberculate trilobites with well-developed eyes. There is no question that the similar morphology is the result of adaptation to similar niches.
To this we add another kind of trilobite convergence, leptoplastiform, which illustrates nicely the concepts developed above. Leptoplastines are Furongian (upper Cambrian) olenid trilobites, quite small, with genal spines springing outwards from the body. In the Leptoplastus Zone in southern Sweden [11] the early form. L. raphidophorus has such morphology with a dorsal spine on the 11th (of 12) thoracic segments. This trilobite has a short vertical range, and is replaced by other, much more modified leptoplastines, which lack the dorsal spine. Further up the column a very similar morphotype L. stenotus appears, likewise with a dorsal spine on the 11th thoraic segment. Is this homologous with that of L. raphidophorus? Or is it independently derived but at the same locus? How are we to tell? This surely most remain an open question, but would a cladistic analysis, without the time-dimension given by the stratigraphy, assume homology? An additional complexity comes from the Mid-Silurian Harpidella (formerly Cyphaspis megalops M´Coy, 1846). This is likewise small, with outwardly springing genal spines, and a dorsal spine (though on the 6th thoracic segment). It looks very like a leptoplastine But whereas the Furongian leptoplastines were adapted to dysoxic conditions, Harpidella lived in shallow, well-lit, and fully oxygenatic environment of a limestone sea. Things may not always be what they seem (Figure 6).
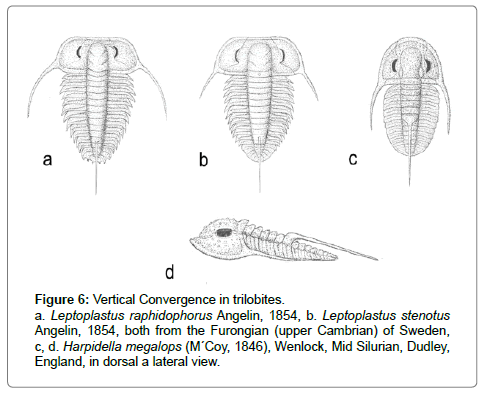
Figure 6: Vertical Convergence in trilobites. a. Leptoplastus raphidophorus Angelin, 1854, b. Leptoplastus stenotus Angelin, 1854, both from the Furongian (upper Cambrian) of Sweden, c, d. Harpidella megalops (M´Coy, 1846), Wenlock, Mid Silurian, Dudley, England, in dorsal a lateral view.
The establishment of systematic relationships due to synapomophies is vulnerable if the direct ancestor for establishing a synapomorphy is not known. This may sound trivial. But for many attempts to discuss examples in the fossil record it becomes important that in cladistic considerations any time axis is missing. As shown here, if functionally similar or different organs even appear in the same morphological context, in consequence of geographic, temporal, or both, geographic and temporal isolation convergent developments may appear, which cannot be distinguished from homologies. Thus the importance of vertical convergence along a time axis may actually be greater than previously, especially when considering organisms at the beginning of the fossil record, when their predecessors lie in the dark of earth history.
We are indepted to thank our anonymous reviewers for their advice that improved this manuscript.