Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2013) Volume 1, Issue 2
Abstract
Background: Mechanisms of action and optimal doses have not yet been determined for the immunomodulator Del-Immune V®, a supplement containing dry fermental cell lysate and DNA fragments of the probiotic strain Lactobacillus rhamnosus V (DV Strain).
Objective: To study the mechanisms of Del-Immune V® immunomodulation and to investigate dose-dependent effects on immunoregulatory cytokines production in vivo and in vitro.
Design: One hundred forty inbred laboratory mice with body masses ranging from 14-16 grams were divided into 7 test groups. Groups I, II, and III received 0.5 ml of aqueous solution of Del-Immune V® by mouth in doses of 5, 50, and 500 μg per mouse, respectively, for 5 days at 24-hour intervals. Group IV mice received 0.5 ml Bifi dim suspension by mouth in a dose of 50 μg/mouse on the same schedule. Group V mice (control group) received 0.15 M NaCl. Group VI and VII mice received a single dose of 50 μg/mouse Del-Immune V® (Group VI) or Bifi dim (Group VII) on day 1 of the test period. Eight hours after administration and every 24 hours thereafter for 5 days, blood serum, peritoneal exudate macrophages, and splenocytes were obtained from each group for testing. Interferonogenous activity of cultured splenocytes and serum levels of interferon were assessed.
Results: Groups I-IV showed a marked increase of IFN levels in blood serum after administration of Del-Immune V® or Bifi dim; the optimal daily dose was found to be 50 μg/mouse. The highest serum IFN level was reported 24 hours after administration. The control group remained unchanged. Maintenance of elevated circulating IFN was possible only through repeated administration.
Conclusions: In this study, a 50 μg/mouse dose of Del-Immune V® was shown to actively induce IFN and moderately stimulate the production of TNF, showing signifi cant promise as an immunomodulating preparation.
Keywords: Lactobacillus rhamnosus; Bifidim; Del-Immune V; Interferon; TNF
Bacterial components are well-established to have an immunomodulatory eff ect. Del-Immune V® is a US Food and Drug Administration-registered food supplement containing dry fermental cell lysate and DNA fragments of the probiotic strain Lactobacillus rhamnosus V (DV Strain). Th e product contains fragments of DNA and cell peptidoglycan of the lactic acid bacteria. Preliminary experimental clinical data suggest that components of Del-Immune V® may be eff ective in stimulating innate immunity [1,2]. However, the mechanisms of activity of the formulation have not yet been verified.
Recent years have been marked by increasing study of the mechanisms of the immunobiological eff ects of probiotics and bacterial-based treatments [2-5]. As a result, supplements such as liastenum (blasten), deodan, licopid, prodigiosanum, salmosanum, sodium nucleinate, MC (molecular composition; yeast DNA and Tilorone), biostim, BCG, rumurtide, ribomunyl, and lactolin are being evaluated, in both trials and clinical practice, for diff erent pathologies [1,6]. Th e adjuvant eff ect of BCG and the immunomodulating activity of formulations containing derivatives of lactobacilli, such as liastenum (Lactobacillus delbrueckii) and deodan (Lactobacillus bulgaricus), have been associated with peptidoglycans and their structural components, muramyl dipeptides (MDP). Th e most active analog of MDP, MurNac-L-Ala-D-Glu-NH2, has demonstrated adjuvant and pleiotropic eff ects and is capable of inducing a number of cytokines: IL-1, tumor necrosis factor (TNF-α), IL-2, IL-6, IL-8, IL-12, and interferon gamma (IFN-γ) [7,8]. Th ese cytokines in turn stimulate nonspecifi c cytotoxicity of lymphocytes and natural killer cells (NK), and coordinate the body’s immune response, depending on the nature of the aggressive agent and the T-helper differentiation (Th 1 or Th 2) [5,7,9]. Th ese properties of peptidoglycans indicate a basis for creating immunomodulating formulations for clinical use. Lactobacilli, which are in the generally recognized as safe (GRAS) group, are good sources of peptidoglycans. Other data indicate that the ability of probiotics to infl uence TLR receptors, which recognize microbe associated molecular patterns (MAMPS) such as peptidoglycan, lipoteichoic acid, and lipopolysaccharide [10], determines their impact on production cytokines. TLR4 and TLR2 have been identifi ed on the surface of lymphocytes and macrophages [5]. TLR2 activation plays an important protective role against infectious diseases caused by gram-positive pathogens, but seems to also be important in providing protective eff ects in infl ammatory diseases. Th e TLR2 mediated inhibitory eff ect occurs through production of TNF-α, but not IL-12p70 [11]. Fragments of probiotic bacterial DNA, which contains unmethylated CpG, are interesting because of their capacity to stimulate production of cytotoxic lymphocytes and NK cells, activate the complement system, heighten cytostatic and cytotoxic activity of macrophages, and induce inflammatory cytokine production [9]. Owing to the ТТТСGТТТ DNA pattern of the strain, Lactobacillus rhamnosus GG was found to have immunostimulatory properties [3]. These DNA fragments from Lactobacillus ramnosus GG induced immune responses via TLR activation in the intercellular endosomes [12].
However, recent observations show that not all functions of the host response to MDP can be explained by TLRs receptors alone. There are other innate immunity components that may be involved, such as nucleotide oligomerization domain (NOD)-like receptors (NLRs) and retinoic acid-inducible gene-I (RIG-I)- like receptors [13-15]. Induced bacterial ligands, including MDP, NOD1, and NOD2, play a key role in the regulation of inflammatory response.
NOD2 recognizes MDP, after which NOD2 undergoes conformational changes to interact with its effector molecules [16]. This interaction is accompanied by induction of IFN regulatory factor 4 (IRF4), a negative regulator of TLR signal [17,18]. Probiotic strains of lactobacilli have been reported to activate NOD2 [19]. Thus, probiotic lactobacilli strains and their particulate derivatives are capable of cytokine induction (IL-10, IFN-γ, TGF-α) and may affect the Th1/Th2 balance. Here, we determine the mechanisms of the immunomodulating effects of Del-Immune V®, describing dose-dependent effects on production of immunoregulatory cytokines in vivo and in vitro.
Mice
The study examined the dose-dependent effect of Del-Immune V® on production of immunoregulatory cytokines in nondescript mice with body masses of 14-16 g. Mice (N=140) were selected on the basis of the analogue principle, and were divided into 7 groups of 20. Animals were fed balanced rodent food and water ad libitum.
Dose finding
Group I, II, and III mice received 0.5 ml of aqueous solution of Del-Immune V® orally by enteral tube feeding in doses of 5, 50, 100 and 500 μg/mouse, respectively, for 5 days at 24-hour intervals. Group IV mice were administered 0.5 ml Bifidim suspension (control probiotic medication) orally in a dose of 50 μg/mouse for 5 days at 24-hour intervals. The Bifidim was a dry mass of Bifidobacterium bifidum N1 (1 mg per vial containing 1×108 CFU) immobilized on enterosorbent in combination with ascorbic acid (Intervetmed Ltd., Kiev, Ukraine). Group V mice were administered 0.15 M NaCl. Group VI and VII mice were used to study the interferonogenous activity of a single administration of 50 μg/mouse of Del-Immune V® (Group VI) or Bifidim (Group VII). Cytokine production by IFN and TNF was examined in control and mice treated with preparation after initial administration 8 hours and then every 24 hours for the next 5 days. For this purpose, several mice from each group were humanely euthanized by cervical dislocation; blood serum, peritoneal exudate macrophages (PEM) and spleen, from which splenocytes were harvested from each group of mice for testing [20,21].
Cytokine assays
The optimal dose of Del-Immune V® was tested via in vitro induction of immunostimulatory cytokines in splenocytes and PEM (1×107 cell/ml) of experimental and control mice by culturing cells with the formulation in final concentrations of 5, 50, 100 and 500 μg/ml. Interferonogenous activity of the tested formulations was assessed in comparison with Bifidim 50 μg/ml and standard inducers (IFN-α; Newcastle Disease Virus, NDV–10 TCD50/cell; IFN-γ; phytohemagglutinin, PHA–20 μg/ml; Difco; TNF, LPS E. coli 0111– 4 μg/ml–Sigma USA). Levels of cytokine production (IFN and TNF) were determined 6, 24, and 48 hours after incubation of the cell with the formulations by cytopathic effect inhibition microtiter assays [22,23].
Biological activity of TNF was assessed by cytotoxicity in the passaged culture of murine fibroblasts L-929 [23]. The result was recorded on a multiscanner (Dynatech, Switzerland) with a wavelength of 540 nm. The cytotoxicity index was calculated using the formula CI=К-О/Кх100%, where K and O represent optical density values for the cell in the culture medium (RPMI 1640 with 10% FCS). The calibration curve based on standard recombinant TNF formulation Sigma was used for standardization of the cytotoxicity index [23]. IFN levels in cell cultures and serum were measured using standard microtitration in the passaged cell culture L-929 against 100 TCD50 indicator virus (vesicular stomatitis virus, Indiana VSV) with constant CO2 level [22].
Student-Fisher t-test was applied to perform comparison between individual groups and the control group [24].
A marked increase in blood serum IFN levels was observed following daily oral administration of Del-Immune V® or Bifidim to Groups I-III over the course of 5 days at all doses (Figure 1). The optimal interferonogenous dose was 50 μg/mouse (Group II). After 24 hours of observation, circulating IFN levels in Group II reached 4.5 ± 0.5 log2 U/ml. After repeated administrations, levels reached 5.5 ± 0.7 log2 U/ml, in comparison with 2.0 ± 0.7 log2 U/ml in the control group (Group V). Further, administration of Del-Immune V® in a dose of 50 μg/mouse on day 3 allowed for maintenance of the 5.5 ± 0.5 log2 U/ml level. Administration of the formulation on days 4 and 5 resulted in nonsignificant decreases in circulating IFN levels. When Del-Immune V® was administered in doses of 5 and 500 μg/mouse (Groups I and III), findings were similar, although maximum interferon levels were not as high.
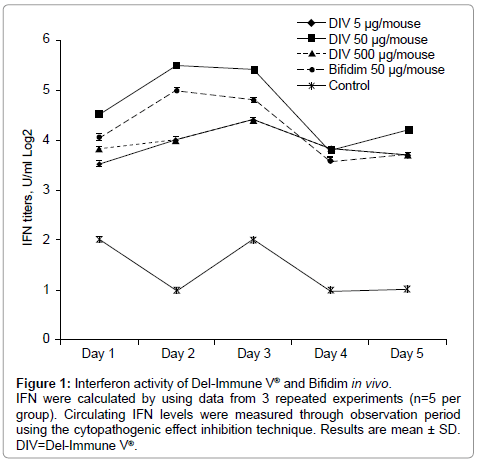
Figure 1: Interferon activity of Del-Immune V® and Bifidim in vivo. IFN were calculated by using data from 3 repeated experiments (n=5 per group). Circulating IFN levels were measured through observation period using the cytopathogenic effect inhibition technique. Results are mean ± SD. DIV=Del-Immune V®.
One-time oral administration of Del-Immune V® or Bifidim to mice in a dose of 50 μg/ml resulted in increased circulating IFN level 8 hours after administration. The highest serum IFN level was reported 24 hours after administration, while levels in control group animals remained unchanged (Table 1).
| Studied formulations; doses of 50 µg/mouse | Serum IFN titers, log2 (mean ± SD) | ||
|---|---|---|---|
| 8 h | 24 h | 48 h | |
| Del-Immune V® | 4.0 ± 0.03* | 4.4 ± 0.03* | 3.41 ± 0.03* |
| Bifidim | 3.6 ± 0.01* | 4.0 ± 0.03* | 2.7 ± 0.03 |
| Control | 2.0 ± 0.02 | 2.0 ± 0.03 | 2.0 ± 0.03 |
Table 1: Murine Serum IFN Titers after One-Time Administration of Del-Immune V® or Bifidim.
Forty-eight hours after administration of Del-Immune V®, serum IFN levels in all active groups remained reliably enhanced in comparison with the control group, but IFN was later eliminated from the body. Maintenance of circulating IFN levels was possible only through repeated administration.
comparative analysis of interferonogenous activity induced Bifidim or Del-Immune V® was performed by testing the interferon-synthesis activity of leukocytes. Splenocytes of the mice receiving experimental formulations were cultured with NDV and PHA inducers, resulting in a 2-fold increase of interferon response in comparison with control animal cells (Figure 2), indicating that the experimental formulations positively affected immune response status. Interferon status was determined by assessing circulating IFN titers (serum IFN), IFN-α, and IFN-γ production by immunocompetent cells as a response to in vitro stimulation and spontaneous IFN production. Enhanced production of IFN-α and IFN-γ was seen in splenocytes 24, 48, and 72 hours after administration of the experimental formulations.
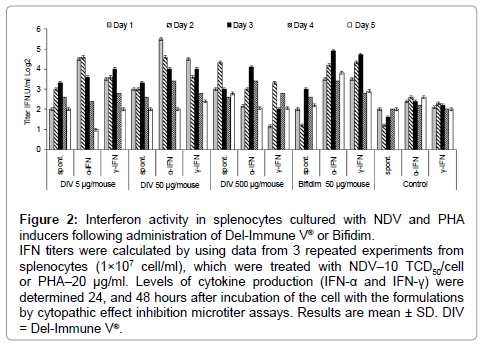
Figure 2: Interferon activity in splenocytes cultured with NDV and PHA inducers following administration of Del-Immune V® or Bifidim.
IFN titers were calculated by using data from 3 repeated experiments from splenocytes (1×107 cell/ml), which were treated with NDV–10 TCD50/cell or PHA–20 μg/ml. Levels of cytokine production (IFN-α and IFN-γ) were determined 24, and 48 hours after incubation of the cell with the formulations by cytopathic effect inhibition microtiter assays. Results are mean ± SD. DIV = Del-Immune V®.
One limitation of the use of IFN inducers is the development of hyporeactive state, in which IFN production is inhibited following repeated administration of the drug. The criteria for assessing refractoriness of animals were determined by assessing INF-α and IFN-γ levels in response to adequate stimulation. Reduced production of IFN-α and IFN-γ under the influence of Del-Immune V® was observed at 4 days following the first injection, followed by gradual recovery of the ability of immune cells to produce interferon. In the Bifidim group, it was possible on day 5 to see restoration of the immunocyte IFN production capacity, when activation of the IFN synthesis capacity of splenocytes was noted. These findings indicate that administration to mice of optimal doses of the probiotic formulations Del-Immune V® and Bifidim on an appropriate schedule stimulates IFN production and increases efficacy of other interferonogenous inducers.
Preincubation of PEM cultures of experimental and control animal cells with Del-Immune V® and Bifidim resulted in cytokine synthesis stimulation, as measured by IFN titers (Figure 3) and TNF concentrations (Figure 4). Adding Del-Immune V® or Bifidim in doses of 5, 50, or 100 μg/ml to PEM cultures of experimental and control mice resulted in IFN synthesis (Figure 3). It should be noted that the IFN activity was dose dependent. Thus, when the concentration was 5 μg/ml, IFN production was much lower than when it was 50 or 100 μg/ml, although it was still almost 6 times higher than the control level. At the same time, concentrations of 50 and 100 μg/ml resulted in an accumulation of stimulated IFN titers with similar values, indicating that an optimal dose for Del-Immune V® is more likely to be close to 50 μg/ml.
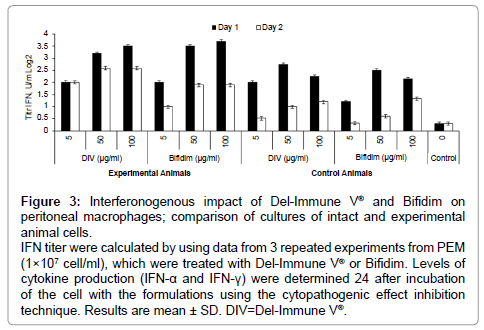
Figure 3: Interferonogenous impact of Del-Immune V® and Bifidim on peritoneal macrophages; comparison of cultures of intact and experimental animal cells.
IFN titer were calculated by using data from 3 repeated experiments from PEM (1×107 cell/ml), which were treated with Del-Immune V® or Bifidim. Levels of cytokine production (IFN-α and IFN-γ) were determined 24 after incubation of the cell with the formulations using the cytopathogenic effect inhibition technique. Results are mean ± SD. DIV=Del-Immune V®.
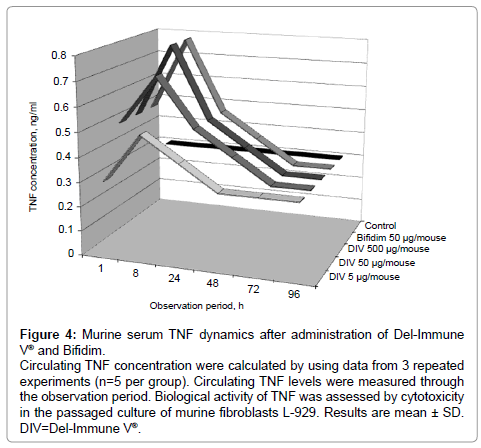
Figure 4: Murine serum TNF dynamics after administration of Del-Immune V® and Bifidim.
Circulating TNF concentration were calculated by using data from 3 repeated experiments (n=5 per group). Circulating TNF levels were measured through the observation period. Biological activity of TNF was assessed by cytotoxicity in the passaged culture of murine fibroblasts L-929. Results are mean ± SD. DIV=Del-Immune V®.
The highest IFN levels in supernatants were reported on day 1 of cell culturing with experimental formulations. However, IFN levels in the control group remained lower than in the experimental groups on both days. Heating serum samples of the animals receiving Del-Immune V® or Bifidim for 30 minutes at a temperature of 60ºC decreased their capacity to inhibit reproduction of vesicular stomatitis virus in cell culture L929 (data not shown). The physical and chemical properties of the IFN produced were characteristic of IFN-α/β and IFN-γ [17]. IFN-γ is produced by sensitized T-lymphocytes CD4+ and CD8+ and NK cells. IFN-γ demonstrates a wide range of immunotropic effects, provides for Th1 differentiation of CD4+ T cells, and stimulates HLA-DR antigen membrane expression; without these functions, identification of bacterial antigens or further activation of T-lymphocytes (including T-helpers stimulating maturation of NK-cells, as well as some subpopulations of B-lymphocytes) is impossible [23]. IFN-γ also participates in the immune response of macrophage cells, inducing production of TNF and IL-1 and modulating their functions [23]. Therefore, the level of TNF was assessed in murine serum (Figure 4) [25-28]. Oral administration of Del-Immune V® or Bifidim in doses of 5, 50, or 500 μg/mouse resulted in endogenous TNF production. After administration of Del-Immune V® or Bifidim in a dose of 50 μg/mouse, serum TNF was 0.6 ng/ml (P<.05) and 0.8 ng/ml (P<.05), respectively, while in the control group it did not exceed 0.3 ng/ml. Maximum production of this cytokine was reported 8 hours after administration of these formulations. Del-Immune V® administered in a dose of 5 μg/mouse resulted in an insignificant increase in circulating TNF concentration to 0.4 ng/ml (P>.05), in comparison with 0.3 ng/ml in the control group. It should be noted that oral administration of Del-Immune V® in doses of 50 and 500 μg/ml resulted in practically equal circulating TNF indices (0.6 ng/ml and 0.7 ng/ml, respectively).
In vitro trials showed that adding Del-Immune V® or Bifidim in concentrations of 5, 50, or 100 μg/ml to macrophages of experimental and control mice resulted in TNF production peaking 8 hours after adding these formulations (Figure 5). TNF production potential of PEMs was dose-dependent. The optimal in vitro concentration of Del-Immune V® and Bifidim was 50 μg/ml.
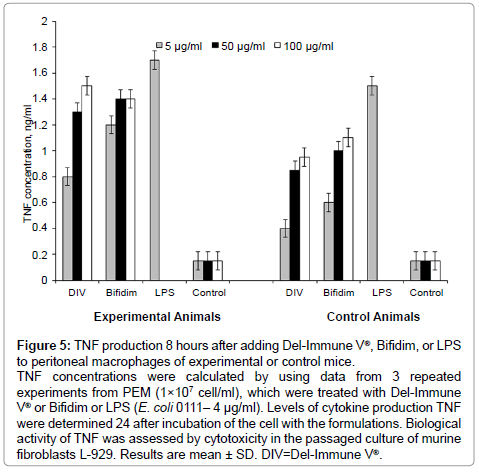
Figure 5: TNF production 8 hours after adding Del-Immune V®, Bifidim, or LPS to peritoneal macrophages of experimental or control mice.
TNF concentrations were calculated by using data from 3 repeated experiments from PEM (1×107 cell/ml), which were treated with Del-Immune V® or Bifidim or LPS (E. coli 0111– 4 μg/ml). Levels of cytokine production TNF were determined 24 after incubation of the cell with the formulations. Biological activity of TNF was assessed by cytotoxicity in the passaged culture of murine fibroblasts L-929. Results are mean ± SD. DIV=Del-Immune V®.
In this study, Del-Immune V®, a US Food and Drug Administration-registered food supplement containing dry fermental cell lysate and DNA fragments of the probiotic strain Lactobacillus rhamnosus V (DV Strain), stimulated the functional activity of murine macrophages and splenocytes. Del-Immune V® demonstrated higher interferonogenous activity in vivo and in vitro than Bifidim. The living cells of Bifidim stimulated in vitro TNF production more intensively than Del-Immune V®. Cytokine production in vitro induced by Del-Immune V® and Bifidim was compared with cytokine production in vivo [29]. Induction of pro-inflammatory cytokines IFN and TNF by Del-Immune V® and Bifidim in vitro suggests that these formulations stimulated a nonspecific immune response in vivo. On the basis of these results documenting the potential of oral Del-Immune V® and Bifidim to stimulate synthesis of IFN-α/β and –γ as well as TNF, it should also be noted that IFN-γ can induce expression of TNF-α receptors on macrophages [30].
These cytokines synergistically stimulate macrophage cells that, in turn, intensify killing activity. IFN- γ increases the expression of class II MHC proteins on professional antigen presenting cells, and so promotes antigen presentation to helper T cells as well. It also enhances the expression of important signaling receptors, TLRs, which is important for the development of protection against viral infections and hypersensitivity, as some of the ligands of these receptors regulate the function of adhesion molecules such as CD11b and L-selectin [5].
Derivatives of microbial origin, including LPS, MDP, and CpG DNA, are identified by immunocompetent cells with TLR receptors [12]. Thus, LPS E. coli stimulates mainly monocytes and macrophages [31]. CD14, the LPS receptor, first interact with microbial components ahead of TLR activation and recognition and signal transduction by the TLR4/MD2-complex [32]. It can be found on monocytes, macrophages, neutrophils, lymphocytes, and bowel epithelial cells. Fixation of microbial derivatives with receptors results in a signal change in the given biological system, which stimulates the synthesis and release of different immunity mediators, or cytokines. It should be noted that gram-positive bacteria, including lactobacilli, activate the major class II histocompatibility complex, which induces IFN-γ and IL-12, which are necessary for Th1 differentiation of T-helpers. Gram-negative bacteria and LPS (a major component of the cell wall of gram-negative bacteria; lipopolysaccharides are endotoxins and important antigens) induce monocytic production of IL-10, inhibiting cytotoxicity activation of IFN-γ and secretion by T- and NK-cells [33,34]. Since clinical applications of LPS and gram-negative bacteria are limited because of high toxicity, finding selective immunomodulators is one of the main conditions for improving the efficacy of immunostimulating therapy.
The synergistic activity of cytokine production induced by Del-Immune V® and Bifidim helps to demonstrate some therapeutic effects of these formulations. The comparative study of Del-Immune V® and Bifidim demonstrated that both formulations had a stimulating effect on cytokine secretion activity of the splenocytes and macrophages necessary for production of IFN and TNF. Bifidim contains living cells of bifidus bacteria, while active substances of Del-Immune V® are MP (muramyl peptides) and nucleoproteids of the probiotic strain Lactobacillus rhamnosus V. This TNF production in vivo call for further studies, since TNF mobilizes leukocytes, terminates inflammatory processes, and plays an important role in the effector and regulatory networks of body immune response. Enhanced TNF production leads to activation of neutrophils, macrophages, and lymphocytes, thus strengthening anti-infection immunity [11,27,35]. A TNF-induced cascade of induction signals results in gradual production of IL-1 and IL-2, activation of T-lymphocytes, and generation of anti-tumor effector cells-lymphokine-activated killers lysing different tumor target cells. TNF intensifies the proliferative response in mixed culture lymphocytes and tumor cells, and demonstrates adjuvant activity for T- and B-lymphocytes. It should be noted that circulating TNF was quickly eliminated from the body.
In this study, higher dosages did not always result in higher efficacy. The success of immune active therapy can be enhanced not only by new medications but also by their rational use. The choice of probiotic formulation (live probiotics cells or structural derivatives of probiotic cells) depends on a large number of factors, including potential, mechanism, mode of administration, and desired immune response. The mechanisms of action of this group of formulations are most likely multifactorial and include a number of signals, cell types, and receptors. One characteristic of probiotic activity is selective effects on the immune system of the macro-organism, whereby only those parts of the natural immune response that require correction are altered [36,37].
Probiotics demonstrate a variety of influences on immunological processes, depending on the type and strain of the bacteria. In contrast, lactobacillus strains, which have strain depended response for induction of cytokines, but have a tendency to activate Th1 [38]. Bifid bacteria and immunogenic derivatives of lactobacillus tended to produce a more anti-inflammatory profile. It is important to identify and characterize the immunomodulatory activity of probiotic preparations. For example, bacteria L. fermentum and L. plantarum stimulate B-cell proliferation, while L. acidophilus mainly causes induction of T-cell immune response [39]. In mouse peritoneal macrophages, Lactobacillus plantarum potently induced IL-10 but weakly induced IL-12 production, whereas Lactobacillus casei potently induced IL-12, but weakly induced IL-10 production [40]. Certain structural components of lactobacilli, including peptidoglycans and DNA fragments, can also influence the secretion activity of human monocytes in vitro through intensified production of IL-1, IL- 6 and TNF-α; in vivo they can activate synthesis of E2 prostaglandin and activate the system of complement and maturation of T-cell precursors [41]. The Lactobacillus casei peptidoglycan inhibits production of inflammatory cytokines [42]. Probiotic components that act as TLR2 agonists are capable of influencing cytokine production signal cascades mediated by common adaptor proteins [12,31,43]. Inhibition of TLR2-induced signaling, influenced by lactobacilli via adapter Mal/MyD88 [34,44], can lead to partial inhibition of TLR4 signal, accompanied by decreased production of proinflammatory cytokines [45,46]. Ability to influence signaling pathways of cytokine production opens new opportunities for the development of probiotic preparations with immunoregulatory properties. In this study, the dry enzymatic lysate powder of a special lactic acid bacteria Lactobacillus rhamnosus (DV Strain) Del-Immune V®, in a dose of 50 μg/mouse, was shown to actively induce IFN and moderately stimulate the production of TNF, showing significant promise as an immunomodulating preparation. Its natural origin, interferonogenous activity, safety, usability, and the possibility of oral administration allow us to consider Del-Immune V® as a supplemental immunomodulator. The dose-dependent responses observed in mice should be further assessed in humanclinical trials to determine whether similar immunomodulation occurs
Medical editor Michael Linde assisted with the preparation of this manuscript.