Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2015) Volume 4, Issue 1
The biology of the macro-element sulfur (S) is attracting an ever growing attention concerning cell physiology and human health. Sulfur metabolism works at the interplay between genetics and epigenetic as well as in the maintain of cell redox homeostasis. Indeed, unbalanced levels of S compounds in the body are actually under investigation as vulnerability factors and/or indicators of impaired cell oxidation state in a variety of human diseases. The purpose of this article is to overview some main S metabolic pathways in humans and their relevance in cell physiology and pathology. Since S is an essential nutrient for life, we first present its distribution and significance in the biosphere, focusing then on S metabolic fluxes which encompass S-containing amino acids (S-AAs), as well as sulfoconjugation, the synthesis and release of H2S together the formation of iron-sulfur cluster proteins. Despite the vastness of the topic, we would like to emphasize herein that the study of S networks in human pathology, especially in complex, multi-factorial disorders, deserves greater impulsion and deepening.
<Keywords: Sulfur metabolism; Sulfur compounds; Methylation; Redox state; Detoxification; Human disease
Among chemical element essential for life, the mineral sulfur (S) presents an unexpected complexity of bioactive derivatives that makes difficult to fully define its tissue distribution, metabolic fates and requirements for humans. A major input to deepen S biology and its impact on human health comes from results obtained in these last decades by the scientific community showing the relevance of S-containing biomolecules, as S-amino acids (S-AAs), reduced γ-L-Glutamyl-L-cysteinylglycine (glutathione, GSH) or 3’-phosphoadenosine 5’-phosphosulfate (PAPS) in a variety of homeostatic processes in the body. Sulfur biomolecules exert important functions in all living organisms and their transformations are involved in metals’ transport, free radicals scavenging, tissue integrity protection, enzyme functionality, DNA methylation and repair, regulation of gene expression, protein synthesis, remodeling of extracellular matrix components, lipid metabolism and detoxification of xenobiotics/ signaling molecules in plants and animals [1,2]. If S biology is intricate, this area of investigation proves to be, at the same time, really fascinating: the widespread presence and diversity of organosulfur bioactive molecules in living organisms can be explained by the fact that this macro-element was one of the predominant atoms on Earth before life and the origin of life has been even attributed to S chemistry. It is indeed probable that former living organisms used S for energy metabolism and breathing under O2-free atmosphere conditions, a theory mirrored by the highly conserved S metabolism in bacteria as well as the evolutionary survival of species belonging to anion sulfide (S2-) producing bacteria as Archea. Moreover, the existence of “strict” anaerobic and sulfate (SO42-)-reducing Desulfovibrio species, which are able to grow under partial aerobiosis, attests the metabolic adaptation occurred during the first passage from a O2-free atmosphere to variably O2-enriched ecological niches [3]. The hypothesis of a common ancestor of plant or animal eukaryote cells deriving from the fusion of aerotolerant eubacteria and sulfidogenic Archea bacteria [4] could explain why most S chemical species play similar functions in plant and animal cells. The fact that the two Kingdoms express different S paths suggests instead that the coupling of oxidative reactions and S pathways for the maintain of chemical homeostasis could have contributed to the early divergence between plant and animal cells. Substantially, the study of S biochemistry provides further evidence to the hypothesis that cell evolution has been favored by mechanisms of interaction, exchange and transformation involving prokaryote and eukaryote genomes [5]. Hence, it follows that S species and S-containing biomolecules occupy a foremost position in metabolism, even from the evolutionary standpoint, an issue which deserves special attention for ecosystems and human health. Because of the complexity of S biochemistry, we have divided the review on S compounds and human health in two parts: the present, a first and more general section, which overviews some main S metabolism substrates, mentioning their involvement in human pathology, especially in complex, multi-factorial disorders; a second one which will deal with data obtained from the literature in clinical research showing S metabolism variation in specific complex diseases as autism spectrum disorders (ASD), schizophrenia and fibromyalgia, by outlining their significance and future perspectives.
Sulfur in nature
Sulfur is a non-metal element, the second of the oxygen (O)-headed VI group in the periodic table. Despite both O and S share many chemical properties and belong to the same group of elements, the S atom, because of its higher radius, is less electronegative and supplies to biomolecules a greater hydrophobic behavior than O. Sulfur chemistry would thus reflect what introduced before: the ancient S metabolism has been preserved under aerobic conditions to restrain the consequences of oxidative metabolism and possible tissue damage by reactive oxygen byproducts [6]. On Earth, S occurs as a mixture of 4 stable isotopes, the most abundant being the 32S species (about 95%). It possesses oxidation states ranging from +6 (SO4 2-, oxyanion sulfate) to -2 (S2-, anion sulfide). Sulfur shows a mean concentration in the biosphere of about 0.05%, being meanly concentrated in oceans where it reaches about the 0.09%. The S biogeochemical cycle assimilates inorganic S species from the outer Earth crust into biomolecules of living organisms through a constant flux of S oxidization states [7]. Soil organic-S/SO4 2- species, reduced by sulfur-reducing bacteria to S2- (S dissimilatory reductions), as well as other S species and elementary S coming from volcanic and ocean sources, pass into the atmosphere and, after meteoric precipitations, into ground waters. Chemolithotrophic bacteria oxidize again them to SO4 2- by enzyme activities as sulfur oxygenase or sulfite oxidase. Sulfate, in turn, is absorbed by fungi and by plant roots through specific transporter proteins as SULTR [8], entering into the alimentary chain. Essentially, S is assumed by autotrophic organisms in the inorganic SO4 2- form, whereas heterotrophic organisms take S as both inorganic and organic S species. Sulfate is activated in all living organisms by a key enzyme which permits to assimilate the anion into cell metabolism: ATP sulfurylase. This enzyme integrates the mineral S in the biochemical machinery of cells from all living organisms, giving then origin to divergent pathways in plant or animal cells [9,10]. The reaction mediated by ATP sulfurylase consists in the formation of adenosine-5’-phosphosulfate (APS) by transferring SO4 2- to ATP and liberating pyrophosphate (PPi). The product of this reaction, APS is thus the activated form of SO4 2- [11]. In plants, S metabolism starts with soil SO4 2- assimilatory reduction pathways forming organic species, firstly the S-AA Cysteine (Cys), which then produces Methionine (Met) and all other bioorganic S derivatives. The reaction catalyzed by the enzyme ATP-sulfurylase represents the first step of Cys formation from SO4 2- in plant cells, through the precursor APS. In plants, two other enzymes, APS-reductase, a flavoprotein, and sulfite-reductase are able to produce a SH- group which is transferred to O-acetylserine generating Cys [1,7]. These metabolic reactions make plants the primary producers of organic S compounds in the alimentary chain. In the animal Kingdom, S as SO4 2- and/or S-containing biomolecules are introduced with beverages and food undergoing several metabolic fates, such as, for instance, Met transmethylation and the Cys-producing reactions of transsulfuration [2]. The requirement of S-compounds is thus provided by the diet, and, for that, S content is among the most abundant macro-elements in animal tissues, just after C,H, O, N, Ca and P [7]. Despite this, Met tissue amounts can be critical owing to the AA use in a variety of fundamental reactions. Indeed, Met is utilized to form S-adenosyl-methionine (SAM), the main methyl donor substrate, and Cys via the intermediates Homocysteine (HCys) and Cystathionine (Cyst). Cysteine subsequently promotes Taurine (Tau), GSH, H2S or SO4 2- formation. Figure 1 depicts the chemical structures of some among the main substrates of human S metabolism, S-containing AAs, some of their derivatives at two or three AAs, as well as S-adenosyl compounds.
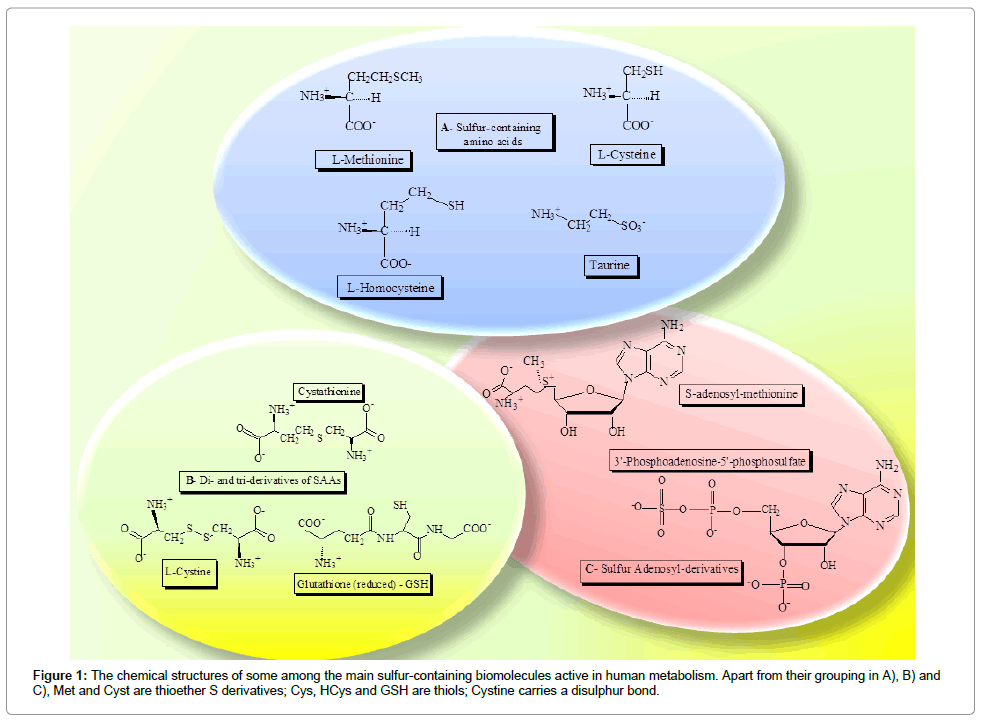
Figure 1: The chemical structures of some among the main sulfur-containing biomolecules active in human metabolism. Apart from their grouping in A), B) and C), Met and Cyst are thioether S derivatives; Cys, HCys and GSH are thiols; Cystine carries a disulphur bond.
Sulfur amino acids and their derivatives in humans
As afore mentioned, in humans, S inorganic species as SO4 2- and S-biomolecules, including S-containing AAs, their derivatives, S-vitamins as biotin or S-coenzymes and cofactors as coenzyme A and α-lipoid acid are obtained from a normal and equilibrated diet. As concerns S-AAs, Met and Cys are proteinogenic S-AAs, whereas HCys and Tau are not contained in proteins. Once introduced with diet, S-AAs enter in cell metabolic paths interlaced with several homeostatic processes in which Met and Cys exert the role of the main players. In the light of all reactions and processes which entail Met, this AA is probably the essential one which influences the greatest number of physiological processes. For this reason, cells are able to recover and maintain diet Met, to avoid its loss. As an example, among other metabolic uses, it is worth mentioning that considerable Met amounts are recruited for mRNA translation: Met is the initiating AA in the synthesis of eukaryotic proteins, while formyl-Met plays the same role in prokaryotes. In addition, Met maintains cell Cys reserve promoting its formation and for that Cys is considered a semi-essential AA which in turn origins many other fundamental S paths. The fact that Met and Cys are both protein AAs implies that their tissue pool is alimented not only by diet but also by hormone-regulated protein turn-over and degradation, as for all other protein AAs. As components of proteins, both Met and Cys are primary intermediates for spatial conformation, assembly and structure stability of peptides and polypeptide chains, regulating their folding features and functionality. Methionine is a thioether compound, a hydrophobic AA involved in interactions within and between proteins and peptides: protein domains rich in Met residues have been identified in active sites of proteins implicated in transport, trafficking or enzymes to prevent oxidation [6,12]. The AA Cys is a thiol, which contains the nucleophilic group –SH able to form S-S bridges. For instance, S-S bridges regulate protein shape and are highly contained in proteins linked to defense against bacteria and viruses (e.g., immunoglobulins and T-cell receptor) or to transport and structural function (e.g., metallothioneins, ferredoxin or keratin); additionally, S-S bridges occupy relevant positions in the catalytic site of several enzymes to yield their active conformations. Extracellular proteins present a higher Cys content vs. intracellular ones (ratio of about 2:1) [13], implying a main role of Cys in connective tissues. Cysteine also contributes to 3D spatial conformation and shape of peptides: the nonapeptides oxytocin and vasopressin, modulators of social behavior in mammalians, contain in their AA sequences two Cys residues bonded through a disulfide bridge. Figure 2 schematically presents the main reactions of Met metabolism in humans, divided into: transmethylation, remethylation, originating the so called “Met cycle”, and the irreversible Cys formation step of the transsulfuration pathway. The Met cycle starts from dietary or protein degradation Met, and then it continues with S-adenosyl-Met (SAM) and HCys formation (transmethylation) or comes back to Met by HCys remethylation [14]. Through irreversible transsulfuration reactions, Met can be instead transformed into Cys, giving origin to Cys metabolism as GSH or Tau formation. The pivotal intermediate of these two Met destinies is HCys. Thus, Met/HCys homeostasis depends upon the subtle balance between three metabolic fluxes: 1) transmethylation to HCys, 2) HCys remethylation to Met (Met cycle), and 3) transsulfuration of HCys to Cys. Methionine, by transmethylation reactions, is the precursor of SAM, the most studied methyl donor substrate in cells which acts in key, SAM-sensitive methyltransferase reactions involving a variety of acceptors and co-substrates, participating, for instance, to the formation of neurotransmitters, the pineal hormone melatonin (MLT), methylglycine, as well as to the processes of methylation of DNA (cytosine adjacent to guanosine residues, CpG sites), histones and phospholipids [15-17]. For this last function, SAM plays main actions in DNA replication and repair whereas its derivative S-adenosyl-HCys (SAH) promotes a powerful negative feed-back on methyltransferase activities [6], favoring HCys remethylation to Met. In more detail, SAM is formed from Met and ATP by Met adenosyltransferase; then, SAM loses a methyl group by a x-co-substrate methyltransferase producing SAH which is, in turn, converted into HCys by the enzyme S-Adenosyl hydrolase (Figure 2, 1-3). Production of HCys can generate transsulfuration reactions to Cys or remethylation to Met through the intersection with folate metabolism or the choline-betaine (trimethylglycine) way (Figure 2- 3,4). Transsulfuration reactions are irreversible steps so that HCys is not converted again to Met. Transsulfuration involves the activity of Cystathionine β-synthase (CBS) which leads to Cyst formation from HCys and subsequent Cystathionine-γ-liase (CGL) action which transforms Cyst in Cys by the release of α-ketobutyrate and ammonia (Figure 2-8,9]. The synthesis of Cys by transsulfuration can produce other S-containing substances as the intra-cell free radical scavenger GSH, synthesized from the 3 AAs glycine, glutamate and Cys, and the non-protein AA taurine (Tau) (Figure 2-18,19]. In summary, S-biomolecules can increase in tissues and bloodstream from their absorption, from the diet or protein degradation (Met and Cys) under hormonal control but their constant levels are guaranteed by the biochemical S “fuel” represented by the Met cycle and transsulfuration, as verified through studies using in vivo Met tracers [18]. The Met cycle is regulated by the gene expression of enzymes: a tissue-specific gene expression/regulation occur. For instance, liver shows the highest expression of Met cycle enzymes, but these last or their isoforms can be also highly expressed in pancreas, kidneys, brain, skeletal muscle and heart [14,19,20]. Transsulfuration enzymes are all equally expressed only in few tissues [20]; in the human brain, a relatively higher CBS/CGL ratio is observed indicating that HCys is prevalently transformed into Cyst in this tissue; in the human kidney and cardiovascular system, a lower CBS/CGL ratio is present and HCys remethylation to Met is predominant [20]. This would imply that Cys production undergoes a regulation on the basis of tissuespecific redox necessities. Moreover, the tissue-dependent gene expression of different enzyme isoforms of the Met cycle changes with age and gender; for instance, HCys remethylation is greater in males than females on normal diet. Hormones as insulin, glucagone or thyroxine orchestrate Met metabolism balance by regulating enzyme expression and activities [21]. After feeding, insulin allows AAs uptake inside cells for protein synthesis, while favoring Met remethylation; glucagon decreases Met levels in the bloodstream while increasing transmethylation and transsulfuration enzyme activities which bring to Cys synthesis to provide reserves of GSH, Tau or other Cys derivatives which can exert their scavenger and anti-oxidant activities during the fasting period. Also, thyroxine promotes SAM formation and HCys transformation into Cys; this explains why unbalanced HCys metabolism is linked to diabetes and hypothyroidism [21]. A pivotal regulation of the Met cycle is given by SAM itself. The role of SAM consists in positive allosteric feedback activation: SAM production in transmethylation path enhances the whole process. The liver is the only district which expresses MAT-I, a high Km Met adenosyltransferase isoform which subtracts the excess of dietary Met [14]. Elevated SAM promotes Met oxidation by transsulfuration, inhibits HCys remethylation to Met, bringing the cycle towards Cys synthesis [6]. A reduced Met availability to transmethylation and transsulfuration is obtained by Met oxidation and conversion to Met sulfone, which is reversed to Met through the action of Met sulfone reductases (MsrAs), a family of enzymes which decrease with aging and neurodegenerative processes [22] (Figure 2, 23). As afore mentioned, also Cys follows a variety of fates, comprising incorporation into protein, Tau or GSH production and oxidation to SO4 2-. High protein diet stimulates both transmethylation and transsulfuration enzyme activities, increasing endogenous production of HCys and Cys [23]. The contribute of Cys synthesis from HCys transsulfuration is important, even in the presence of dietary Cys: this is mainly due to the important amount of Cys consumed for generating GSH, Tau and other cofactors and biomolecules. Reactive oxygen species (ROS) and peroxide species, formed under stress-related conditions, promote transsulfuration, while the antioxidant ones reduce it: this indicates that in the presence of oxidant species, transsulfuration provides Cys for GSH request, while increased antioxidants slow down Cys synthesis, less demanded for GSH synthesis. Thus, stress, ROS formation and apoptosis signals can be counterbalanced by pro-oxidant activities, whose efficiency and availability is critical for cell health. For GSH synthesis, Cys enters together glutamate into the γ-glutamyl cycle (Figure 2) [18,19) forming the dipeptide γ-glutamyl-L-cysteine by γ-glutamyl-L-cysteine synthetase; Gly is then added to the dipeptide by glutathione synthetase. This tripeptide, more concentrated inside cells rather than body fluids, represents a storage form of Cys and its disulfide derivative Cystine. The structural GSH feature is the γ-glutamyl linkage, which facilitates its role in a remarkable variety of reactions and functions such as protection of sulfidryl groups of coenzyme A, other cofactors, proteins and enzymes. In addition to being one of the major protective molecules for hepatocytes, GSH exerts multiple roles in all cells, as presented in the next paragraph. Next to GSH, another product of Cys metabolism is Tau, a sulfonated β-amino acid (2-aminoethanesulfonic acid). This non protein S-AA is synthesized in the animal Kingdom and results from the activity of Cys dioxygenase to produce the intermediate cysteine sulfuric acid (3-sulfinoalanine), which is then decarboxylated by sulfino-alanine decarboxylase into hypotaurine. Hypotaurine is then oxidized to Tau [23] (Figure 2, 10-12)
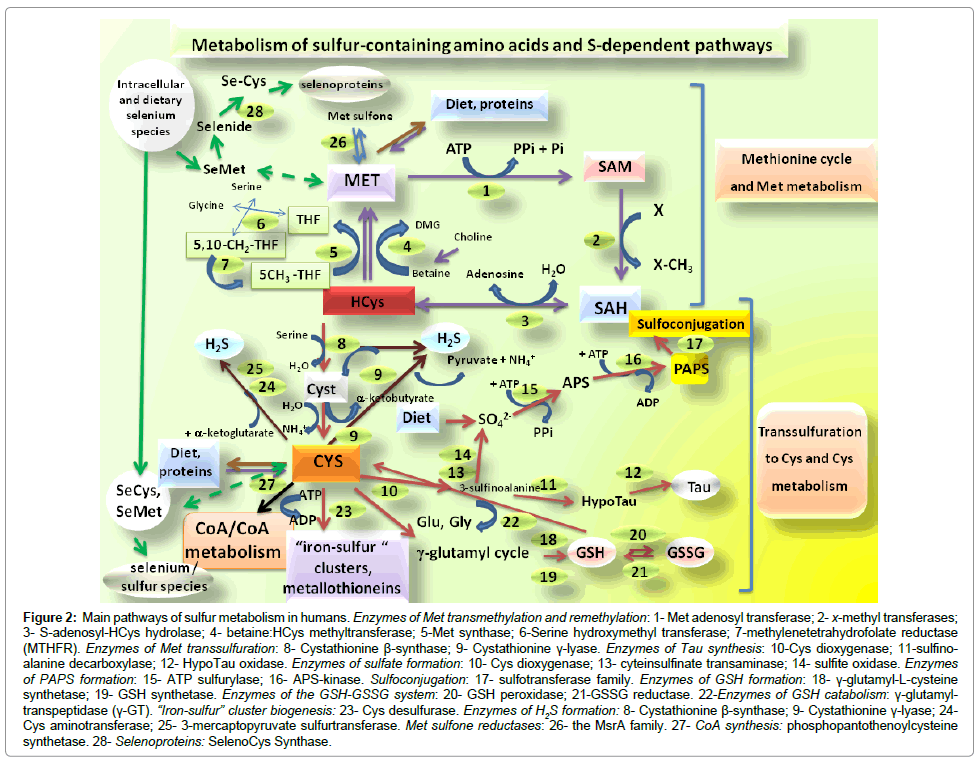
Figure 2: Main pathways of sulfur metabolism in humans. Enzymes of Met transmethylation and remethylation: 1- Met adenosyl transferase; 2- x-methyl transferases; 3- S-adenosyl-HCys hydrolase; 4- betaine:HCys methyltransferase; 5-Met synthase; 6-Serine hydroxymethyl transferase; 7-methylenetetrahydrofolate reductase (MTHFR). Enzymes of Met transsulfuration: 8- Cystathionine β-synthase; 9- Cystathionine γ-lyase. Enzymes of Tau synthesis: 10-Cys dioxygenase; 11-sulfinoalanine decarboxylase; 12- HypoTau oxidase. Enzymes of sulfate formation: 10- Cys dioxygenase; 13- cyteinsulfinate transaminase; 14- sulfite oxidase. Enzymes of PAPS formation: 15- ATP sulfurylase; 16- APS-kinase. Sulfoconjugation: 17- sulfotransferase family. Enzymes of GSH formation: 18- γ-glutamyl-L-cysteine synthetase; 19- GSH synthetase. Enzymes of the GSH-GSSG system: 20- GSH peroxidase; 21-GSSG reductase. 22-Enzymes of GSH catabolism: γ-glutamyltranspeptidase (γ-GT). “Iron-sulfur” cluster biogenesis: 23- Cys desulfurase. Enzymes of H2S formation: 8- Cystathionine β-synthase; 9- Cystathionine γ-lyase; 24- Cys aminotransferase; 25- 3-mercaptopyruvate sulfurtransferase. Met sulfone reductases: 26- the MsrA family. 27- CoA synthesis: phosphopantothenoylcysteine synthetase. 28- Selenoproteins: SelenoCys Synthase.
Taurine has been found highly concentrated in animal and human cells, especially in excitable tissues as brain, retina, muscle and hearth; it is also present in the liver and kidney, while it’s circulating levels being, in comparison, much lower. Taurine function has been coupled to antioxidant activities, osmoregulation and modulation of Ca2+ levels [24]. In the CNS, where Tau is highly concentrated, this β-AA acts as a neuromodulator and neuroprotector, also preserving the homeostasis of retinal functions. Taurine plays a role also in bile salts formation. Nevertheless, the full comprehension of its tissue distribution and tissue-specific physiological role deserves additional investigation. Taurine, by its chemical structure, is supposed to stabilize membranes interacting with phospholipids, contributing to their fluidity and excitability. Taurine physiology has been linked to the classical neurotransmitter γ-aminobutyric acid (GABA): Tau can mimic some effects provoked by GABA release, including the interaction with the GABAA Cl- ionophore receptor [25].
Also coenzymes and vitamins contribute at upholding Met metabolism homeostasis. First of all, the vitamin folic acid and its derivative tetrahydrofolate (THF) which generates the so called “one C metabolism” and HCys remethylation to Met (Figure 2- 4,5). Folic acid is synthesized by dihydrofolate reductase requiring NADPH. Glycine and Ser inter-conversion produces THF and 5,10 methenyl- THF, this last deriving from Gly decarboxylation. Vitamin B-12 is required for Met synthase function and FAD is the prosthetic group of Methylenetetrahydrofolate Reductase (MTHFR). Impaired activities of reductases and transulfuration enzymes CBS and CGL have been observed in case of loss of vitamin B-6 (pyridoxal phosphate) necessary for their catalytic function. This explains why deficiency in one of these vitamins, cofactors and groups provokes high HCys circulating levels [6,26]. Finally, it should be pointed out that, beside gender and aging, the different development stages can affect the activity of enzymes participating to the Met cycle [27]. Even if we do not treat this topic, the interaction between Met, Cys and selenium (Se) species coming from diet or intracellular metabolism, represents another big chapter in S biology: seleno-Met leads to the formation of selenide species, the co-substrate of the enzyme seleno-Cys synthase (Figure 2- 27, 28) , an enzyme able to transform Ser-tRNA into seleno-Cys-tRNA, this last incorporated into selenoproteins. The Se-S species and selenoproteins possess a strong antioxidant activity in the body, especially the compound seleno-Cys [28]. Figure 2 (23) also refers to other paths linked to Cys, as CoA formation or iron-sulfur cluster biogenesis and synthesis of metallothioneins, small Cys-rich proteins, which also plays a fundamental action in the metabolic anti-oxidant defense machinery against oxidative stress and metal/heavy metal toxicity.
The reduced and oxidized glutathione system
The GSH anti-oxidant system deserves a separate paragraph. Glutathione is in fact the most abundant thiol compound in cells and its function is critical for the maintain of the cell redox status. Moreover, this compound binds to xenobiotics and several endogenous compounds for detoxification purposes [29]. Several reviews, as the paper of Wu and coauthors [30], exhaustively describe the GSH relevance in cell physiology. In particular, once formed from Cys in tissues and cells, GSH fates are multiple: it can exert “per se” a feedback regulation on its own synthesis and, concomitantly, it undergoes homeostatic redox reactions involving the activities of two main enzymes, glutathione peroxidase and glutathione reductase (Figure 2). These two enzymes are part of the cell anti-oxidant defenses constituting the GSH-Px/GSSG-Rd system which maintains the suitable redox state of cells and tissues [31]. The selenoenzyme Glutathione peroxidase transforms GSH into GSSG (glutathione disulfide or oxidized form) to counteract hydroxyperoxides’ formation avoiding cell damage [32]. Several isoforms of GSH peroxidase are expressed, each capable to counteract the generation of diverse hydroxyperoxide chemical species. Glutathione disulfide is transformed back to GSH by the GSSG reductase enzyme activity, a NADPH-dependent activity, as in thioredoxin system [33]. The extra/intracell levels of GSH are also regulated by the enzyme gamma-glutamyl transpeptidases (γ-GT, Figure 2, 22) in cells and tissues expressing this activity: γ-GT is able to break down the tripeptide originating again Cys interacting with AA metabolism. This enzyme is acquiring further interest in human health since it is able to maintain the homeostasis of both GSH or Cys and can regulate cell proliferation/apoptosis [34,35].
Sulfoconjugation and detoxifying function
This paragraph presents the relationships between S, S biotransformations and detoxification processes. The metabolic inactivation of endogenous signals and xenobiotics represents a pivotal step of cell response regulation, avoiding their noxious accumulation in the adult body and during development. Inactivation is mediated by phases I and II detoxifying enzymes, which promote, as the final passage, the excretion in urines of bioactive compounds transformed into water-soluble derivatives. One of these activities is sulfoconjugation or sulfation, mediated by the families of sulfotransferases (STs) [36]. Sulfotransferases, in the presence of the S compound PAPS as the co-substrate and sulfate donor, inactivate a variety of bioactive compounds. These last include drugs, hormones, and neurotransmitters as catecholamines or end products of catabolism producing hydrophilic sulfo-derivatives excreted in urines. Sulfation is an essential cell reaction implicated also in other processes as the remodeling of glycosaminoglycans, polysaccharide components of the extracellular matrix and proteoglycans, implying a main role in structural function and formation of connective tissues. Sulfation and ST activity also participate to post-translational modifications of tyrosine residues in secretory proteins [37]. The S molecule PAPS is produced by 2 ATP molecules, SO4 2- and the activity of two enzymes, ATP sulfurylase and APS-kinase (Figure 2 – 15, 16). Bifunctional forms of ATP sulfurylase/APS-kinase have been also identified in humans. Anyway, in substance, ATP and SO4 2- availability significantly affect the sulfation capacity of STs. Diet and water are the main sources of SO4 2-. After assumption with diet, both intestinal absorption and renal re-absorption of SO4 2- contribute to maintain the homeostasis of the body amount of this anion as well as its availability to STs action [38]. Sulfate can be generated also by Cys after its transformation into Cys sulfinate by Cys dioxigenase, an enzyme in common with the Tau synthesis path, subsequent formation of sulfinylpyruvate by cyteinsulfinate transaminase, release of sulfite and oxidation to SO4 2- through the action of a sulfite oxidase (Figure 2 - 10,13,14] [38]. As a consequence, increased amounts of Met and Cys can raise SO4 2- levels. Other enzyme activities which boost the SO4 2- intracellular pool are lysosomial sulfatases involved in cell turn-over, whose activity leads to macromolecule degradation [38]. Sulfotransferases forms a heterogeneous class of enzymes: actually, 5 families of STs have been identified and each can recognize a specific group of substances. These families are also differentially expressed in anatomical districts and brain. It has been suggested that brain STs play a crucial role in the CNS development. Research on ST families is an open field and the most studied ST1 one is actually divided into 5 subtypes [36,38]. The ST1A is highly expressed in brain, platelets, liver, breast and gastrointestinal tract. Its preferential substrates are catecholamines or phenols and can be also distinguished in thermo-stable or thermo-sensitive forms [39]. The ST1B family produces sulfation of thyroid hormones whereas ST1C overall recognizes xenobiotic compounds and ST1E promotes the sulfoconjugation of estrogens. The ST2 family targets cholesterol, pregnenolone, DHEA and neurosteroids. Intriguingly, there is also evidence that STs contributes to fetal development and mother-fetus exchanges through sulfate transporter molecules [40,41]. In our lab, we have previously demonstrated that platelet ST shows seasonal rhythms [42], depending on gender [43], underlining that cells can adapt detoxifying activities or neurotransmitter signaling to photoperiod as well as to the different necessities during the lifespan in men and women separately.
Hydrogen sulfide as a gasotransmitter in humans
A promising new interest concerning investigation on the biology of S chemical species in mammalians and humans relates to the preserved capacity of eukaryotic cells at producing the sulfide acid H2S, the final product of dissimilatory reactions in SO4 2--reducing bacteria. In particular, H2S, considered highly toxic, a poisonous gas with a typical odor of rotten eggs, is now classified among the endogenously produced gasotransmitters involved in many cell homeostatic processes as nitric oxide (NO) and carbon monoxide (CO), each other interacting [44,45]. This acidic molecule, is present in cells as HS- or as a persulfide bound form in proteins, and is an example of the relative concept of toxic, noxious compounds for life functions. H2S body levels are maintained very low by pO2 which enhances its clearance into thiosulfate and sulfate. Enzymes implicated in H2S formation are the transsulfuration enzymes CBS and CGL, which catalyze Cyst and Cys formation from HCys. The first produces serine, H2S, pyruvate and ammonia starting from Cys, the second induces Cys dimerization to Cystine, forms thiocysteine and its rupture to H2S. The enzyme CGL has been found to catalyze H2S formation through several pathways [46] (Figure 2- 8,9]. CGL can produce Cyst and H2S starting from Cys and HCys. A third minor synthesis pathway is H2S formation by 3-mercaptopyruvate sulfur transferase (3MST) and Cys aminotransferase using Cys and α-ketoglutarate as the substrates [44- 47]. These 2 enzymes are expressed both in the brain and periphery (Figure 2 – 24, 25). As aforementioned, CBS and CGL are differentially regulated in the body, thus CGS produce H2S overall in the CNS, while CGL forms it in peripheral districts, cardiovascular system and smooth muscle cells. Among the main homeostatic roles played by H2S in cells, there are cardiovascular functions, respiration and muscle relaxation; moreover, H2S release has been implicated in the regulation of blood pressure through the opening of ATP-dependent K+ or voltage-gated channels of vascular tissues producing a vasorelaxant action [48- 51]. Other H2S roles include apoptosis regulation, acting as a kind of metabolic “sensor” to prevent tissue damage and to counteract inflammation [52]. More, as most S chemical species in the body, H2S promotes scavenging of ROS. The discovery of H2S endogenous production has paved the way towards the comprehension of its regulation in human disease: defective H2S production has been related to arterosclerosis and impairment of the cardiovascular function [45,52]. H2S production is inhibited by the other two gasotransmitters via CBS inactivation. Insulin and glucocorticoids regulate CBS/CGL and can exert opposite effects, inhibitory or stimulatory, respectively, on H2S release. Interestingly, some authors have even observed a sex hormone modulation of the gasotransmitter release [53]. Another district where H2S acts as a modulator is the gut, where the gas is also produced by S-metabolizing microbiome strains [54], Gram-negative species or aerotolerant Proteobacteria [55]. In the brain, H2S has been linked to hippocampal long-term potentiation, to the negative feedback of glucocorticoids release and neuroprotection [56].
Iron-Sulfur cluster proteins and human metabolism
Another key section of S biology embraces iron-sulfur (Fe-S) cluster proteins and their regulatory action at the level of the respiratory chain and mitochondrial function. Iron-sulfur clusters are prosthetic groups contained in metalloproteins, as NADH dehydrogenase, xantine deydrogenase, coenzyme Q reductases, Krebs cycle enzymes (aconitase and succinate dehydrogenase), flavoproteins and ferredoxins involved in the mitochondrial respiratory chain, proteins of DNA metabolism and repair or iron-regulatory proteins as IRP1 [57]. Among these clusters there are the [2Fe-2S] or the [4Fe-4S] forms, composed by tetrahedral iron atoms bounded to S and to proteins through sulfidryl -SH Cys groups. The biogenesis of these Fe-S groups in mammalians cells has been elucidated and consists in a series of reactions concerning the enzyme Cys desulfurase (NFS1) [57] (Figure 2, 23). In mammalians, these reactions start with the formation of a protein complex including the enzyme NFS1 and a bound scaffold protein (ISCU). Once again, this S pathway, precisely a Cys pathway, needs vitamin B6 pyridoxal phosphate as the cofactor together an additional monomeric protein, the ISD11. The complex transforms 2 molecules of Cys into 2 molecules of Alanine (Ala) in the presence of iron (2 Fe2+). The frataxin protein, together a ferredoxin e- donor, also participates to the enzyme complex formation and the first steps of “iron-sulfur” cluster biogenesis. Once formed, Fe-S clusters are transferred to specific recipient proteins, providing to these ones a versatile capacity to accept or donate electrons depending on redox potential, oxidation state and conformational changes. The Fe-S clusters are highly conserved in bacteria, yeasts, plants, animals and high vertebrates. First living organisms may have used early formed Fe-S clusters to capture and release electrons under primeval conditions. The importance of Fe-S clusters has been evidenced by molecular biology studies and genetic defects of Fe-S cluster containing proteins. A variety of inherited mutations of genes codifying for these proteins has been identified: these mutations are linked to a wide spectrum of severe human diseases, encompassing the neurodegenerative disease Friedreich ataxia, a sideroblastic anemia and a form of myopathy which derives from mutations of the Fe-S cluster assembly scaffold factor ISCU [58]. The research on proteins containing these prosthetic groups is relatively recent and, for this reason, represents an open field of investigation [59]; it is thus conceivable to think that other human diseases and mitochondrial malfunctions could be related to an altered expression/regulation of this group of proteins within tissues.
Sulfur biology and human disease
From such considerations, it comes out that S biotransformations entail much awareness for human pathology. Sulfur metabolism is the heart of a variety of basic homeostasis functions inside cells and tissues. Sulfur compounds play a protective role during immune response [60] and in a variety of other cell responses [61]. The fact that genetic Mendelian defects at the levels of the Met cycle and transsulfuration enzymes lead to relevant syndromes and disorders linked to hypermethionemia, homocystinuria or cystathioninuria further underlines the importance of S metabolism in the body [14]. On the other hand, changes in S compounds’ formation and catabolism have been reported as significant metabolic indexes in complex diseases with a multifactorial and multifaceted pathogenesis. Polymorphisms or unbalanced S metabolism and S-AAs levels have been revealed in cardiovascular diseases, musculoskeletal disturbances, diabetes and neurodegenerative disorders as Alzheimer disease [62-67]. Moreover, S metabolism investigation needs to be deepened in the field of clinical research on complex diseases and overlapping conditions as obesity, neuropsychiatry and pain perception disorders, all at unclear etiology [68]: the second section of this review will specifically deal with the involvement of S metabolism in some specific human complex disorders and the possible use of valuable instruments to apprise the levels of S intermediates as useful parameters in this clinical research field.
The chief role of S metabolism in cells and its tight coupling to oxidative stress suggest the involvement of S compounds in basic functions as the homeostasis of chemical cell composition as well as in defense and detoxifying mechanisms. Sulfur inorganic/organic compounds work as main metabolic sensors in the modulation of environment-cell interplays. The development of S biology studies can pave the way toward new research fields in human disease and, potentially, in pharmacology. For instance, a better evaluation of the mechanisms of regulation, gene expression of involved enzymes in specific tissues during the lifespan, the formation and use of SO4 2-/H2S within cells is prospected together the investigation of S organic species which interact with metals/semi-metal in Fe-S clusters or Se-S species and their regulation, or the assessment of epigenetic DNA/protein SAM-dependent methylation patterns and the activity of the GSH/ GSSG system.