Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2015) Volume 6, Issue 4
Surface topography of anterior adhesive apparatus was studied of the gill monogenean parasite Diplectanum sp. Diesing, 1858 inhabiting the marine water fish Dicentrachus sp. using SEM. The study revealed that the parasite has three head lobes on each anterolateral region of the head; each lobe accommodates a single, ventrally located adhesive sac. These sacs receive secretion from glands openings that located on the lumen of these sacs. Rodshaped secretory bodies and granular irregularly shaped bodies which may be secretory bodies of the adhesive gland cells were seen open into the adhesive sacs. The tegument of the anterior adhesive area of Diplectanum sp. was characterized by many microvillous like structure. The possible functions of these distinctive features in the temporary attachment are discussed. Numerous adhesive papillae presumed sensory structures were found associated with the tegument of the anterior adhesive area and ventral surface of the body. A single ciliary structure supported by a collar of tegument occurs singly close to the mouth opening. It is suspected to be ciliated sensory ending serve to locate feeding sites. The possible functions of these presumed sensory structures were discussed. The parasite adaptation in its microhabitat in facing the strong water current inside the host fish gills was discussed
Keywords: Monogeneans anterior adhesive apparatus, Gill monogenean parasite iplectanum sp, Sensory structures of Diplectanum sp
The anterior body region of monogeneans plays a crucial role in attachment, movement, feeding and reproduction [1,2].
Most adult monopisthocotylean monogeneans use an anterior adhesive region for temporary attachment to the host when posterior haptoral attachment is severed during leech-like movement [3]. This adhesive enables instant attachment and detachment of the anterior region. Preliminary chemical characterization of adhesive has been achieved for a variety of monogenean parasites [4,5] and the adhesive mechanism has been studied for the adults of two species from different families: the capsalid Entobdella soleae [6] the monocotylid Merizocotyleicopae [7].
The present SEM study was undertaken to investigate the Surface topography of the gill monogenean parasite Diplectanum sp., particularly those of the anterior adhesive areas in order to elucidate questions on maintenance and survival of the parasite on their microhabitat.
The monogenean parasites Diplectanum sp. Diesing, 1858, were recovered from the gills of Dicentrachus sp. fish (Dicentrachus labrax and Dicentrachus punctatus) that collected from the Mediterranean coast at Ras El-Bar at Damietta Province, Egypt. For electron microscopy technique, the monogenean gill parasites were washed several times with distilled water, then fixed for about 1/2 h at 4°C in 1% osmium tetroxide in 0.1% sodium cacodylate-HCI. They were washed for about 1 h in several changes of cold buffer (0.1 M sodium cacodylate containing 3% sucrose and 1% M CaCl). Post-fixation was carried out for about 2 h using 2.5% glutaraldehyde buffered to pH 7.3 with 0.1 M sodium cacodylate-HCI at 4°C. The specimens were left in the washing buffer overnight and were then dehydrated using an ascending series of acetone solutions. They were then critical point dried in carbon dioxide using a Polaron Critical Point Dryer, coated in gold and thus, a coated specimen become ready for mounting on the stub and examination using JEOL JSM 5300 at 20 kV at Alexandria University.
Scanning electron examination revealed that the body of Diplectanum sp. usually appears bending towards the ventral surface. The body of the presented parasite is elongated, with an anterior elongated prohaptor and a posterior attachment opisthaptor (Figure 1). The mouth opens subterminally on the ventral side (Figures 1-3) and having a lower lip-like structure, the tegument of which forms an irregularly arranged series of many thick tegumental folds (Figure 4).
The anterior body region has three head lobes on each anterolateral region of the head (Figures 1-3 and 5). Each lobe bears a single adhesive sac (Figures 2 and 3). These sacs receive secretion from glands openings that located on the lumen of these sacs (Figures 3 and 6). Rod-shaped secretory bodies and granular irregularly shaped bodies which may be secretory bodies of the adhesive gland cells were seen open into the adhesive sacs (Figure 6). Mucus-like substances were also detected in this region (Figure 6). There are many small openings lying in the anterior-most region of head (Figure 7). Considerably, large openings were also found in this area (Figure 7). The tegument covering the anterior adhesive areas is microvillus (Figure 7).
Numerous surface ciliary dome-shaped papillae were distinguishable on the head lobes close to the adhesive areas (Figure 5). Each papilla is elevated above the tegument and has a relatively long cilium-like structure (Figure 8). Ciliated structure occurs singly close to the mouth opening supported by a collar of tegument (Figures 4 and 9).
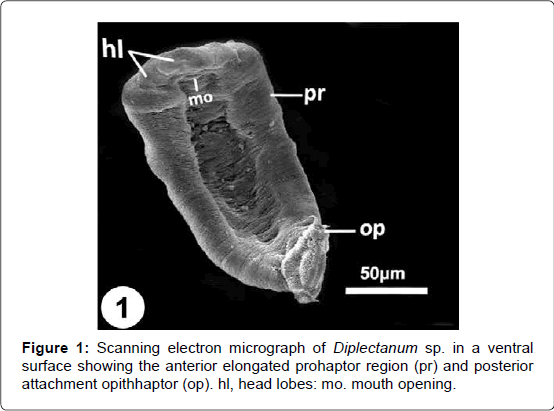
Figure 1: Scanning electron micrograph of Diplectanum sp. in a ventral surface showing the anterior elongated prohaptor region (pr) and posterior attachment opithhaptor (op). hl, head lobes: mo. mouth opening.
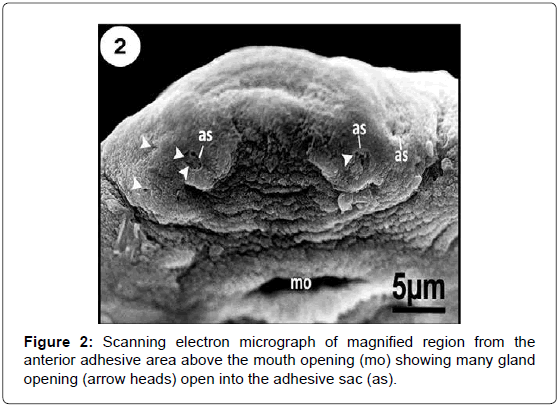
Figure 2: Scanning electron micrograph of magnified region from the anterior adhesive area above the mouth opening (mo) showing many gland opening (arrow heads) open into the adhesive sac (as).

Figure 3: Scanning electron micrograph of magnified region from the adhesive sac (as) showing mucus-like material (mu) and rod-shaped secretory bodies (rd) and granular irregularly shaped bodies (sb).

Figure 4: Scanning electron micrograph of the microvillous tegument (mv) of adhesive area showing large gland openings (lo) and many small gland openings (so).
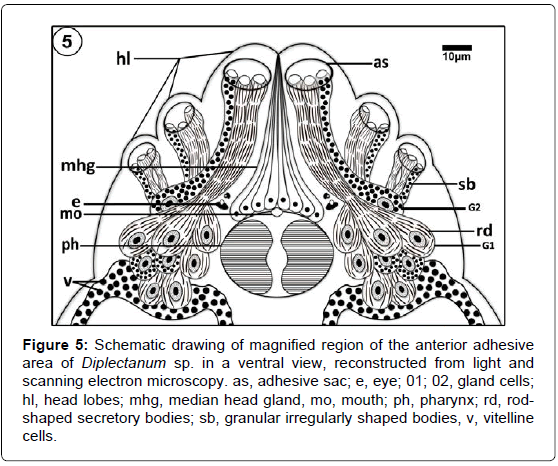
Figure 5: Schematic drawing of magnified region of the anterior adhesive area of Diplectanum sp. in a ventral view, reconstructed from light and scanning electron microscopy. as, adhesive sac; e, eye; 01; 02, gland cells; hl, head lobes; mhg, median head gland, mo, mouth; ph, pharynx; rd, rodshaped secretory bodies; sb, granular irregularly shaped bodies, v, vitelline cells.

Figure 6: Scanning electron micrograph showing magni?ed dome-shaped adhesive papillae (sc) elevated above the tegument bearing long cilium (arrow head).
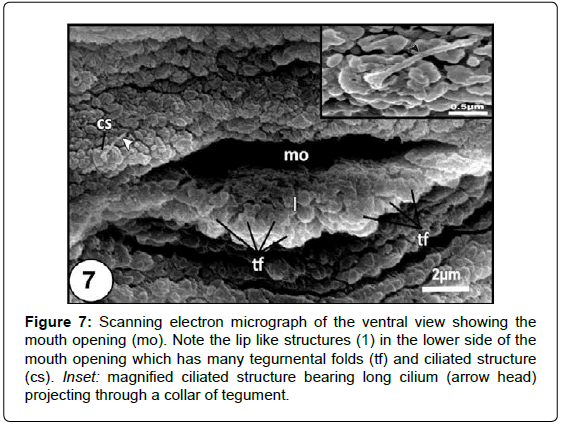
Figure 7: Scanning electron micrograph of the ventral view showing the mouth opening (mo). Note the lip like structures (1) in the lower side of the mouth opening which has many tegurnental folds (tf) and ciliated structure (cs). Inset: magni?ed ciliated structure bearing long cilium (arrow head) projecting through a collar of tegument.
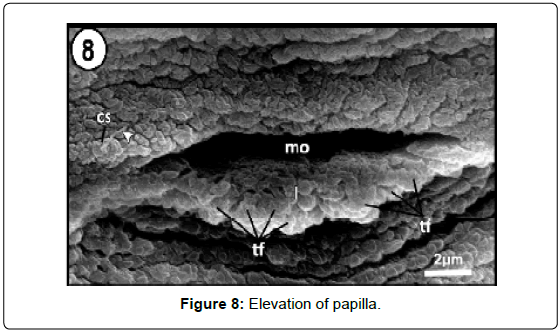
Figure 8: Elevation of papilla.

Figure 9: Occurrence of ciliated structure.
Scanning electron observations in the present study revealed that the anterior adhesive apparatus of Diplectanum sp. generally resemble those of gill-infesting Dactylogyrids [8] and the skin-infesting Entobdellids [9] in being composed of three distinct adhesive sacs encloses three head lobes in each side of the head. Demonstrations said that the capsalid Entobdella soleae has two ventral adhesive pads, one on each side of the head region. Each of these pads is subdivided into three areas [6]. In the monocotylid monogeneans Empruthotrema raiae and E. torpedinis, which inhabit the nasal fossae of their elasmobranch hosts, the gland ducts open into three eversible adhesive sacs on each side of the head region [10]. The adhesive sacs may indirectly assist feeding by firmly attaching the head region to the host and by holding the mouth in the feeding position, a function for the anterior adhesive apparatus which has been suggested by [11,12]. However, [13] noted that the anterior adhesive areas of Entobdella soleae appear not to be attached to the host while feeding is taking place. It seems likely that such an arrangement with six points of adhesion to the host surface is to provide strong resistance to shear forces exerted on the Diplectanum sp. by water current from any direction [14].
Two kinds of gland cell (G1 and G2), one producing rod-shaped bodies (rd) and other producing granular secretory bodies (sb) on both sides of the head associated with the anterior adhesive apparatus of Diplectanum sp. were described by [15] at the light microscope level. The ducts of these gland cells terminate into three pairs of adhesive sacs located on the distal extremity of the ventro-lateral margins of the head region. Scanning electron microscopy of the present study confirms the presence of two kinds of secretory bodies (rod-shaped and granular-shaped). So, it is possible that the large openings open into the adhesive sacs in the anterior head region of Diplectanum sp. represent the opening of the gland ducts (G1 and G2). In this respect, the anterior attachment apparatus of Diplectanum sp. resembles that of other gill-infesting monogeneans, Cichlidogyridsv [16] and diplectanids [17]. The apparatus was also described in skin-infesting monogenean, Entobdellids [9] and Acanthocotyle [14]. However, one kind of gland cells producing roughly spherical secretory bodies was reported by to supply this apparatus in the stomach-infesting monogenean Enterogyrus cichlidarum. Previous observations on Entobdellids have been interpreted to mean that the rods provide the adhesive material and that the granular secretion may be responsible for rapid detachment sticky areas [9]. Another possibly is that the granular secretion serves to create stickness by interacting in some way with rods [14].
Moreover, many small openings were detected on the anteriormost region of the head. During the present study some considerably large openings were also found on the anterior median region between the large openings of adhesive sacs. The small openings may represent openings of tegumental glands, while the large openings on the anterior-most median region of the head may be the openings of the anterior median head glands that observed at the light microscope level by [15] to open in this region. The multiple openings have also been recorded in other monogeneans [18].
It is noticeable that the tegument of the adhesive apparatus of Diplectanum sp. is amplified by microvilli, feature which is also characteristic of the tegument covering the adhesive areas of Diplectanum aequanus [19]. These microvilli may increase the surface area of the worm, suggesting that, on morphological grounds, the tegument has potential as a site for metabolic exchanges into and out of the worm [1,20-22] and absorption of \ from the surrounding environment [23] also, might support a mat of mucus that might afforded osmoregulatory or antibacterial protection [22,24]. In this way, experimental studies by [25] demonstrated that Diclidophora is capable of trans-tegumental absorption of certain neutral amino acids. The significance of this is as unclear since the worm is sanguinivorous and undoubtedly derives most of its food from blood which is ingested and degraded in a well differentiated gut [26]. Suggestion about the fact that a monogenean tegument is microvillous and has an absorptive potential provide evidences of a close phylogenetic relationship between Monogenea and Cestoda, moreover, adds support to [27] argument for a close affinity between ancestral monogeneans and ancestral cestodes [22]. Postulated that the occurrence of microvilli and the absorptive function of the tegument provide evidence of a close phylogenetic relationship between Monogenea and Cestoda. The function of theses microvilli in Diplectanum sp. is not known. However, it was suggested by [8,28] these microvilli may assist in spreading and mix the secretory materials of different gland cells and increase the surface area available for binding the adhesive secretion of the head region during locomotion. In this respect, absence of microvilli from the rest of the head region may complete the action of the presence of microvilli of the adhesive sacs. Moreover, the thin thickness of the head lobes tegument compared to that of the other body tegument is thought to. The present study supports [28] hypothesis, in that the stubby microvilli of the adhesive apparatus helping Diplectanum sp. to mix and spread sticky different secretions of gland cells as a tacky film over the host during locomotion.
In the present study, a number of distinctly different types of presumed sensory receptors were mapped on the surface of the investigated species. Those ciliated sensory endings structures present on the body surface may be rheo-or tango or chemoreceptors aiding in orientation of the parasite’s body with respect to gill architecture and in relation to the direction and flow of ventilation water currents.
Monogeneans are richly equipped with numerous receptors [4,29- 32]. Presumed sense organs are not only numerous in the tegument but each species has several different types to which a variety of functions have been ascribed. Uniciliated, non-ciliated and various other types of presumed sensory receptors have been described on the body surface and haptor of monogeneans and other Platyhelminthes [19,23,33,34].
There is little experimental evidence to suggest the functions of the various types of receptors found in Platyhelminthes but all functional interpretations of presumed sensory structures remain uncertain and are based solely on their distribution and ultrastructural anatomy and are therefore speculative.
Sensory endings like those found on the tegument of some monogeneans [35] have been observed in other invertebrates, and were considered to be touch and rheoreceptors. Some of these sensilla (rheo- , chemo-, photo- and mechanoreceptors) could possibly be used for host finding [4,32,36,37] particularly touch receptors, since the fluke depends on it in locating and maintaining contact with the host. The ciliary structure presumed sensory receptors may be rheoreceptors/ tangoreceptors involved in the orientation of the fluke’s body in relation to the flow of the ventilation water currents. Non ciliated receptors may have a chemo-mechanoreceptor involvement in site selection for feeding and, possibly, in mating during cross-fertilization. [38] recorded two new sensory receptors in Gyrodactylus sp. the authors suggested that the first type may have a pressure/contact or photoreceptoral function, while the second type closely resembles presumed photoreceptors found in several other parasitic and freeliving Platyhelminthes.
An additional suggestion concerning the function of receptors is that they may play an important role in the survival of monogeneans when the worm occupies a sheltered position inside the branchial chamber of the host and not subjected to full force of the water current [35]. Also these papillae equipped with and penetrated by nerves, serving in the attachment to the host [2]. It is evident that they respond actively to both mechanical and chemical stimuli.
The possible functions of the ciliary sensilla and dome-shaped structures of Diplectanum sp. in the anterior areas may act as tangoreceptors or chemoreceptors. The dome-shaped structures may be concerned with the control of the attachment and detachment mechanism of the anterior adhesive sacs to help in maintenance of the parasite’s adhesive attitude between gills.
The present study revealed that the presence of ciliary structure close to the mouth opening in Diplectanum sp. may be mechanoreceptors involved in the orientation of the parasite in relation to the flow of the ventilation water currents for feeding [39-42]. Mentioned that such sensory structures were already encountered in monogenea, and serve to locate feeding sites. In this respect, Diplectanum sp. support suggestion made by [13] that the majority of monopithocotylean living on skin and gills of their hosts are epidermis feeders.
The present parasite under discussion is highly adapted in its parasitizing site. All the former structures (adhesive sacs, glands, microvilli and sensory papilla) help the parasite in facing the strong water current inside the fish gills.
Consequently, the present study investigate that the anterior adhesive apparatus of the gill monogenean parasite Diplectanum sp. masks many wonderful adaptations to parasitism in facing the strong water current inside the fish gills. Furthermore, the associated structures of the anterior adhesive apparatus (adhesive sacs, glands, microvilli and sensory papilla) can elucidate the maintenance and survival of the parasite on their microhabitat.