Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2013) Volume 1, Issue 4
Development of dairy products containing bifido bacteria is one of the main focuses in the food industry, considering the health benefits attributed to its survival through gut intestinal tract and the liberation of biogenic compounds. In order to evaluate the effect of process conditions on survival of Bifidobacterium animalis subsp. Lactis HN019 and release of biogenic compounds during cold storage two technological processes were employed: (i) addition of Probiotic (unfermented bifido milk; UFBM) and (ii) fermentation (fermented bifido milk; FBM). B. Lactis HN019 counts were stable only in fermented milks during seven weeks of cold storage. Moreover, dairy matrices associated with cold storage appeared to protect bifidobacteria during exposition to gastric conditions, assuring the correct Probiotic counts at gut entrance. The distribution of fatty acids was affected by the different Probiotic products. In contrast, fermentation process increased the relative contents of some bioactive fatty acids, such as elaidic acid (+11%) and conjugated linoleic acids (+20%). It was also noted an increase in polyunsaturated fatty acids in the FBM and a higher relative content of monounsaturated fatty acids in the UFBM, as compared to control milk. In addition, fermentation process by bifido bacteria enhanced the long chain fatty acids fraction in bifido milks as compared to control and unfermented bifido milks. Control milk and unfermented bifido milk showed the same peptides even after seven days of storage in contrast with fermented milks that had increase bioactive peptides. It is possible to suggest that opioid peptides are formed during fermentation process, increasing the source of bioactive peptides. Finally, the cold conservation process modified peptides and could improve antibacterial activity.
Keywords: Bifidobacterium animalis subsp. Lactis HN019; Fermented milk; In vitro simulated digestion; Bioactive metabolites
Development of dairy Probiotic products is one of the main focuses in the food industry. Regarding the benefits of dairy functional foods, milk is known, beyond its nutritional properties, to contain bioactive compounds that may promote some beneficial effects on the host, such as anti-atherosclerotic and anti-carcinogenic activities [1,2].
Bifidobacteria are natural members of the human intestinal microbiota, in which they occur at concentrations of 109 to 1011 cells per mL of feces, and represent up to 91% of the total gut population during the early stages of life [3]. Interest of bifidobacteria for human health is related to their survival through gastrointestinal tract and to their role for stimulating the immune system and for preventing microbial gastroenteritis [4,5]. Furthermore, bile salts can also trigger physiological adaptive responses in bifidobacteria [6]. Biogenic compounds such as bioactive peptides and fatty acids, produced by bifidobacteria were shown to be a possible mechanism for their health enhancing properties [7,8].
There are many studies describing the effects of probiotics in humans and animals, from preventative and therapeutic points. The beneficial characteristics of probiotic strains include physiological, immunological, metabolic and genetic features. Besides that, the functional aspects depend on technological properties of bacteria chosen, as well as, the technologies applied to produce the functional food. Therefore, probiotic activity is not changed just by the strain specificity but also by the technological process used and the matrix in which it is delivered [3].
Based on pioneering studies by Metchnikoff in 1907, Tissier in 1906 [9,10] and the notion that ingested live bacteria could have a beneficial effect has been developed and pared down into the idea of “Probiotics”, a term that has generated several definitions over time. There currently appears to be a consensus concerning the definition published by an expert committee of the FAO and WHO, which states that Probiotics are “Live microorganisms which when administered in adequate amounts confer a health benefit on the host” [11].
Fermented milks are recognized as the main vehicles for Probiotics consumption [12,13], nevertheless, the inclusion of bifidobacteria in these products is a great challenge in the food industry. The development of fermented milks containing bifidobacteria has gained attention the last years as these probiotic bacteria grow slowly in milk due their lack of proteolytic activity [14,15]. According to Mercenier et al. [16] and others, some important characteristics are expected of probiotic strains as great survival, proliferation and metabolic action at the target site.
However, to promote the health effects probiotic bacteria must be viable and in high number during storage period and have to persist, even for short periods, in gastrointestinal tract. After consumption, bacteria must resist to extreme conditions encountered in this environment. They must to overpass the stressful conditions such as strong acidity, oxygen, bile salts, cholesterol, pancreatine and compete with the resident microbiota. It has been demonstrated that the dietary intake of beneficial fatty acids and Probiotic bacteria may impact on the modulation of microbiota and consequently resulting in health benefits on the host [17]. Conjugated linoleic acid (CLA)in fermented milk and dairy products should provide “functional” aspects such as inhibition of initiation of carcinogenesis process, effects on anti-atherogenic, anti-adipogenic, anti-diabetogenic and anti-inflammatory activities, beneficial regulatory effects on immune function, and alteration in the low-density lipoprotein/high-density lipoprotein cholesterol ratio in humans [18]. In addition, other bioactive substances of food origin can be considered as dietary components that exert a regulatory activity in human organism, beyond basic nutrition [19]. Some peptides show multifunctional properties such as opioid, antihypertensive, antimicrobial, antithrombotic, immune modulation, and metalbinding activities [20]. Until now, few data is available in literature concerning the effect of technological process, fermentation or addition of probiotic cultures in the production of bioactive compounds in functional products. From our knowledge just the study of Zareba et al. [21] has demonstrate the differences in volatile compounds in fermented and unfermented milks produced by Bifidobacterium animalis subsp. Lactis BB12.
Based on previous works, Bifidobacterium animalis subsp. Lactis HN019 was chosen by its immunogenic capability and by their technological properties, such as great tolerance to acid and bile salts stress [18,22]. Moreover, when compared to B. Lactis BB12, a widely employed probiotic strain and B. Lactis HN019 showed shorter fermentation time in co-culture [18,23,24] higher CLA and α-linolenic acid (ALA) expression [23,24].
Taking into account these considerations, this study aims at analyzing and comparing the probiotic resistance upon in vitro gastrointestinal digestion and the release in bioactive metabolites in unfermented and fermented milks containing bifidobacteria using the same matrix, probiotic strain and probiotic dose.
Experimental procedure
Bifidobacterium animalis subsp. Lactis HN019 (DuPont-Danisco, Madison, USA) was the probiotic strain used in this study. Commercial UHT skimmed milk (Molico, Nestlé, Araçatuba, SP, Brazil) from the same batch, was stored at 4°C until use.
Two technological processes were employed: addition and fermentation, resulting in two kinds of products, respectively, Unfermented Bifido Milk (UFBM) and Fermented Bifido Milk (FBM). Briefly, for addition procedure, at first a pre-culture was prepared inoculating 1.2 g of B. Lactis HN019 in 50 mL of sterilized milk (121°C/15 min). Afterwards, 49 mL of the pre-culture was added to 450 mL of skimmed milk, mixed during 5 min, than was dispensed in 60 mL polyethylene terephthalate cups (PET 60 mL) and immediately cold stored at 4°C. In parallel, for the fermentation process, 500 mL of skimmed milk was inoculated with 1 mL of the same pre-culture, and tempered at 37°C. Fermentation was conducted until pH 4.7 controlled by CINAC system (Ysebaert, Frépillon, France) [25,26]. After achievement of pH 4.7, the fermentation was stopped by rapid cooling in an ice bath during 30 min until reach 10°C, than was dispensed in 60 mL polyethylene terephthalate cups (PET 60 mL) and immediately cold stored at 4°C.
Probiotics counts as well biogenic compounds - fatty acids and peptides were determined before fermentation (D0), after 24 hours (D1) and 7 days (D7) of product´s storage at 4°C. Products were submitted to in vitro simulated gastrointestinal conditions, and survival of Probiotic bacteria was investigated. Samples were prepared at least in duplicate, and the experimental procedure was replicated twice on different days.
In vitro evaluation of gastrointestinal survival
In vitro evaluation upon simulated gastrointestinal conditions of B. Lactis HN019 – gastric and enteric, was conducted according to Baruzzi et al. [26] with adaptations. Briefly, products obtained from different technological processes - FBM and UFBM, at D1 and D7 of storage at 4°C were submitted to three phases of in vitro digestion (gastric, enteric I and enteric II). Initially, for gastric digestion, the samples were suspended in 200 mM pH 1.93 orthophosphoric acid (H3PO4) at room temperature. Porcine pepsine (Sigma-Aldrich, São Paulo, Brasil) was added to buffer at 16.000 U.mL-1 for two hours at 37°C under 150 rpm agitation. Subsequently, before the start of enteric phase I, samples were washed twice and centrifuged at 5500xg for 10 min at 4°C (Centrifuge Eppendorf 5810/5810R, Hanppange, NY, USA). The pellet was dissolved in 6 g.L-1 porcine pancreatine (Sigma pr. num. P7545, 8× USP specification activity), diluted in 0.1 M TRIS buffer, pH 8.39 at ambient temperature (corresponding to pH 8.00 a 37°C) and maintained during two hours at 37°C under 150 rpm agitation. At the end of enteric phase I, samples were washed twice (5500xg, 10 min, and 4°C). Finally, enteric phase II was initiated suspending the pellet in sterile saline 0.09% solution containing 0.1% of porcine bile salts (Sigma-Aldrich, São Paulo, Brazil) and 0.3% bovine bile salts (Sigma-Aldrich, São Paulo, Brazil) diluted in the ratio 1:10 in de Man Rogosa Sharpe Broth culture media, incubated for one hour under 50 rpm stirring at 37°C. Samples were collected at the end of each phase, and enumeration of probiotic viable cells conducted. Survival was expressed as concentrations attained at different intestinal sites. Samples were prepared in duplicate, and the experimental procedure was replicated twice on different days.
Enumeration of viable bacteria
Bifidobacterium animalis subsp. Lactis HN019 was enumerated by pour plate technique in Reinforced Clostridia Agar (Oxoid, Basingstoke, UK) added with 2 μg.mL-1 of dicloxacillin (pH 7.1) and 0.3 g.L-1 aniline blue (InLab, SãoPaulo, Brazil) incubated at 37°C for 72 h under anaerobic conditions (AnaeroGen, Oxoid, Basingstoke, UK) [27]. Results were done based on at least four replicates measurements in each experiment.
Biogenic compounds determination
Fatty acids: The fermented and unfermented bifido milks were submitted to lipid extraction according to ISO method 14156 [28]. Fatty acids methyl esters (FAME) of milk lipids were prepared by transesterification according to ISO method 15884 [29], that consists of a base-catalyzed methanolysis of the glycerides, followed by a neutralization with crystalline sodium hydrogen sulfate to avoid saponification of esters. Analyses of FAME were carried out in a gas chromatograph (model 430 GC, Varian Chromatograph Systems, Walnut Creek, California, USA) equipped with a CP 8412 auto injector. Injections were performed in a 100 m fused silica capillary column (ID = 0.25 mm) coated with 0.2 μm of polyethylene glycol (SP- 2560, Supelco, USA) and analysis were carried out under the following conditions: using helium as carrier gas; isobaric pressure of 37 psi;
linear velocity of 20 cm.s-1; make- up gas: helium at 29 mL.min-1 at split ratio of 1:50; volume injected: 1.0 μL. The injector temperature was set at 250°C and the detector temperature at 280°C. The oven temperature was initially set at 75°C for 3 min, then programmed to increase to 150°C at a rate of 37.5°C.min-1, and then to 215°C at a rate of 3°C.min-1 [30]. Qualitative fatty acid composition of the samples was determined by comparing the retention times of the peaks with those of standards 05632 and 189-19 (Sigma, Chemical Co., St Louis, MO, USA). The Galaxie Chromatography Data System (Varian Chromatograph Systems) was used for identification and quantification of peaks. Fatty acid composition of the samples was determined by comparing the retention times of the peaks with those of standards 05632 and 189- 19 (Sigma, Chemical Co., St Louis, MO, USA). The relative content of each FAME was calculated from the area of each peak, and expressed as a percentage, according to the official method CE 1-62 (AOCS, 1997). Results were grouped and expressed as percentages of short chain fatty acids (SCFA - C4:0 and C6:0), medium chain fatty acids (MCFA -C8:0 to C15:0), long chain fatty acids (LCFA - C16:0 to C20:4), saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), and polyunsaturated fatty acids (PUFA), according to Ackman [31]. Control milk (M), FBM and UFBM at D1preparations were analyzed in sextuplicate.
Peptides: To analyze the potentially bioactive peptides present in milk, UFBM and FBM the samples were microfiltered (0.20 μm, Milippore, Billerica, MA, USA). Skimmed milk was used as control (Molico, Nestlé, São Paulo, Brazil). One mL of the filtrate was stored in a freezer at -80°C until being analyzed. Analytical Central, Chemistry Institute of São Paulo University performed the analysis. Peptides were analyzed by HPLC coupled to mass spectrometry analyzer type ion trap LC-MS (Bruker Daltonics MicroTOF, Billerica, Massachusetts, USA) with time-of-flight analyzer Esquire ESI-MS (n) low resolution. For analysis, the samples are thawed at room temperature to be injected into the C18 column at flow rate of 1 mL.min-1. The peptides were eluted thought 0.01 min gradient 5% B 30 min 60% B, 35 min 100% B, 40 min 100% B, 5 min 45% B, where B is 100% acetonitrile, adjusted to a wavelength of 280 nm. All analyses were conducted in quadruplicate.
Statistical analysis: Multifactor analyses of variance and multiple comparison tests were done in order to determine statistical significance of differences among samples. Mean values were compared using Tukey test at P ≤ 0.01 and P ≤ 0.05. Allstatistics analyzes were performed using Statistical version 8.0 (Stat Soft Inc., Tulsa, USA). All results of cell counts, peptides and fatty acid composition were expressed as the mean of four replicates.
B. Lactis HN019 survival in the product and after in vitro simulation of gastrointestinal digestion B. Lactis HN019 cell counts were measured after 24 hours (D1) of storage at 4°C; in unfermented bifido milk (UFBM) and fermented bifido milk (FBM) are shown in Figure 1A as initial counts. These data have shown that counts of viable bifidobacteria in each product – UFBM and FBM were not significant different after one day of storage. In contrast, after seven days of storage, the initial counts were significant lower (Figure 1B) in UFBM with a decrease of 0.6log CFU.mL-1 (P ≤ 0.05).
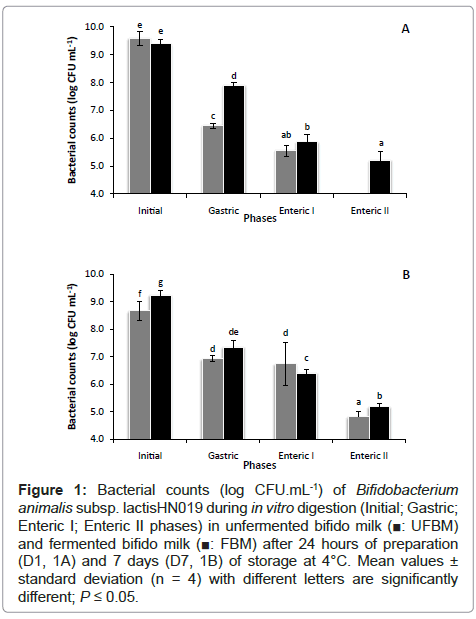
Figure 1: Bacterial counts (log CFU.mL-1) of Bifidobacterium animalis subsp. lactisHN019 during in vitro digestion (Initial; Gastric; Enteric I; Enteric II phases) in unfermented bifido milk (■: UFBM) and fermented bifido milk (■: FBM) after 24 hours of preparation (D1, 1A) and 7 days (D7, 1B) of storage at 4°C. Mean values ± standard deviation (n = 4) with different letters are significantly different; P ≤ 0.05.
During the gastrointestinal transit, Probiotic bacteria undergo drastic physiological stresses, such as strong acidity, presence of digestive enzymes and bile salts, which significantly affect their survival. It could be observed a reduction in survival of B. Lactis HN019, reaching 6.4 log CFU.mL-1 and 7.9 log CFU.mL-1 respectively for UFBM and FBM after the end of gastric conditions when simulated digestion was realized with products sampled at D1 (Figure 1A). After enteric phase I counts decreased but no significant differences were observed for counts between UFBM and FBM. However, after enteric phase II, B. Lactis HN019 do not survive in UFBM and different behavior was noted in FBM in which counts remained at 5.2 log CFU. mL-1 (Figure 1A).
Figure 1B shows counts of B. Lactis HN019 after in vitro gastrointestinal digestion of fermented milks sampled after 7 days of chilled storage (D7). After gastric phase, counts decreased significantly in both fermented milks. Counts in UFBM remained stable after enteric phase I but decreased after enteric phase II attaining 4.8 log CFU.mL-1 at the end of in vitro digestion. In contrast, counts in FBM after enteric phase I decreased as well as after enteric phase II reaching 5.2 log CFU.mL-1.
B. Lactis HN019 in UFBM at one day did not resist to in vitro gastric conditions. However, after seven days of storage, Probiotic bacteria in unfermented and fermented milk submitted to cold storage adapted itself, then, impacting on further resistance upon the in vitro digestion conditions in enteric phases I and II of viable cells respectively. These data confirm the findings of Madureira et al. [32], who reported that dairy matrix protects B. Lactis HN019 throughout stomach acidity, assuring the correct Probiotic counts at gut entrance. Finally, it is considered that survival investigation of Probiotic bacteria within the gastrointestinal tract in vivo is necessary in order to confirm its efficacy in the host.
Biogenic compounds released in the products
Bioactive fatty acids: The fatty acid composition of lipids extracted from control milk (CM), UFBM and FBM is shown in Table 1. It could be seen that the main fatty acids in the products corresponded to saturated fatty acids, like palmitic (C16:0) and stearic acid (C18:0). Afterwards, the monounsaturated (MUFA) fraction was mainly represented by oleic acid (C18:1cis 11) and elaidic acid (C18:1 trans 9) (Figure 2A). Finally, polyunsaturated fatty acids were composed predominantly by linoleic (C18:2 cis 9, cis 12) and conjugated linoleic acid (C18:2 cis 9, trans 11) (Figure 2B and 2C, respectively). This distribution according to saturation degree was within the range reported by Rodríguez- Alcalá et al. [33] with cow milk samples and by Estrada et al. [34] with strawberry yoghurts. Considering the products analyzed in this study, control milk showed the higher relative contents in SFA as compared with the probiotic products (P ≤ 0.01). Consequently, MUFA and PUFA fractions were lower in control milk. Fermentation process contributed to the increase in PUFA fraction, due to the bacterial metabolism that provided isomerization and desaturation reactions in some long chain fatty acids (LCFA) [7].
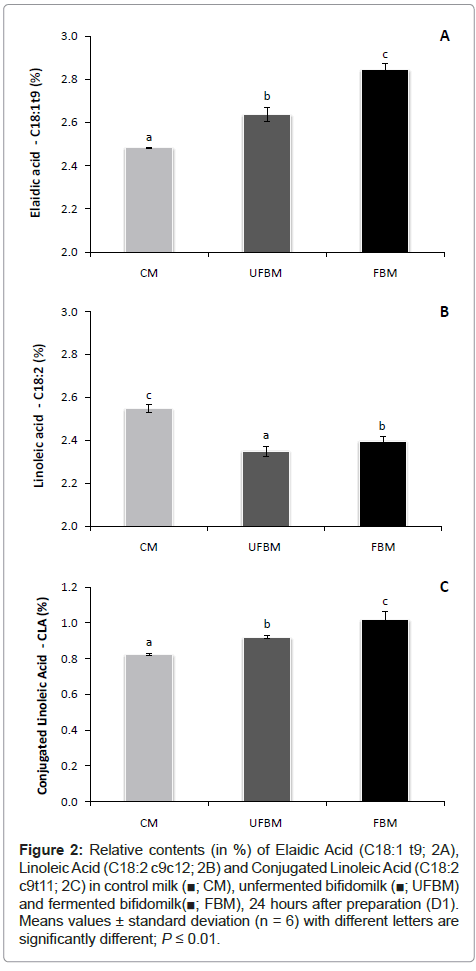
Figure 2: Relative contents (in %) of Elaidic Acid (C18:1 t9; 2A), Linoleic Acid (C18:2 c9c12; 2B) and Conjugated Linoleic Acid (C18:2 c9t11; 2C) in control milk (■; CM), unfermented bifidomilk (■; UFBM) and fermented bifidomilk(■; FBM), 24 hours after preparation (D1). Means values ± standard deviation (n = 6) with different letters are significantly different; P ≤ 0.01.
Taking into account the distribution according to the chain length, it could be seen that short chain fatty acids (SCFA) increased (P ≤ 0.05) with the addition of Probiotic culture, independently of the technological process used, as compared to control milk, and this data agree with those of Ekinci et al. [35], who observed higher amounts of SCFA in products fermented by Probiotic, propionic bacteria and lactic acid bacteria. Moreover, the relative contents in medium chain fatty acids (MCFA) concentration decreased (P ≤ 0.05) while LCFA increased (P ≤ 0.05) during fermentation process, as previously reported by Florence et al. [23] with yoghurts and yoghurts fermented by bifidobacteria species.
In addition to these results, significant differences in individual fatty acids were observed between control milk and probiotic products. Higher amounts (P ≤ 0.01) of capric (C6:0), caprylic acids (C8:0) were found in probiotic fermented or unfermented milks, while caproic (C10:0) and myristic acid (C14:0) were considered as lower (P ≤ 0.01) as compared to control milk (Table 1). Moreover, milk fermentation by B. Lactis provided higher relative levels of palmitic acid, elaidic (C18:1 trans 9) and conjugated linoleic acid (CLA) while it decreased stearic, oleic, linoleic and α-linolenic acids (ALA; C18:3) (Table 1 and Figure 2).
| Fattyacids (%) | Control milk | Unfermented milk | Fermented milk |
|---|---|---|---|
| SCFA* | 4.15 ± 0.04a | 4.44 ± 0.02ab | 4.40 ± 0.13b |
| C4:0 | 2.61 ± 0.01a | 2.65 ± 0.02a | 2.61 ± 0.14a |
| C6:0** | 1.54 ± 0.04a | 1.79 ± 0.01b | 1.79 ± 0.01b |
| MCFA* | 11.61 ± 0.03b | 11.38 ± 0.11a | 11.26 ± 0.21a |
| C8:0** | 0.84 ± 0.01a | 1.17 ± 0.00b | 1.18 ± 0.01b |
| C10:0** | 3.12 ± 0.01b | 2.86 ± 0.02a | 2.90 ± 0.02a |
| C12:0 | 3.75 ± 0.01b | 3.62 ± 0.06a | 3.68 ± 0.04ab |
| C14:0** | 0.69 ± 0.02b | 0.59 ± 0.00b | 0.29 ± 0.13a |
| C14:1 | 1.03 ± 0.05a | 0.94 ± 0.00a | 0.99 ± 0.07a |
| C15:0 | 1.72 ± 0.00b | 1.65 ± 0.02a | 1.66 ± 0.04ab |
| LCFA* | 84.14 ± 0.08a | 84.18 ± 0.10a | 84.34 ± 0.21b |
| C16:0* | 37.21 ± 0.06ab | 37.00 ± 0.10a | 37.40 ± 0.22b |
| C16:1 | 0.61 ± 0.05a | 0.62 ± 0.00a | 0.63 ± 0.03a |
| C17:0* | 0.90 ± 0.00a | 0.92 ± 0.01ab | 0.95 ± 0.02b |
| C18:0* | 11.86 ± 0.05c | 11.52 ± 0.06a | 11.65 ± 0.07b |
| C18:1* | 26.90 ± 0.01b | 26.97 ± 0.01b | 26.46 ± 0.26a |
| C18:2* | 2.55 ± 0.02c | 2.35 ± 0.02a | 2.40 ± 0.02b |
| C18:3** | 0.21 ± 0.02b | 0.12 ± 0.00a | 0.11 ± 0.01a |
| SFA** | 65.00 ± 0.03b | 64.52 ± 0.13a | 64.84 ± 0.07ab |
| MUFA* | 31.10 ± 0.01a | 31.74 ± 0.13b | 31.27 ± 0.21a |
| PUFA* | 3.70 ± 0.03b | 3.75 ± 0.02a | 3.89 ± 0.05b |
Table 1: Relative contents of the main fatty acids (%) in control milk, unfermented bifido milk and fermented bifido milk, 24 hours after preparation (D1).
From Figure 2, it can be observed that significant changes (P ≤ 0.01) occurred when Probiotic culture was added or grown in milk and as also compared to control milk. Addition and fermentation process enhanced, at different levels, the relative contents of elaidic (Figure 2A) and CLA (Figure 2C). On the one hand the simple addition of Probiotic culture increased by 6% and 11% the relative contents of elaidic and CLA contents, respectively. On the other hand, fermentation process, contributed to an increase of 11% and 20% of elaidic and CLA fatty acids relative contents, respectively. As a consequence, both technological processes also ascribed a reduction of 8% (UFBM) and 6% (FBM) in linoleic acid relative levels (Figure 2B).
These changes in the profile of bioactive fatty acids could be related to the activation of bio hydrogenation pathway [7]. As previously reported by some authors [18,22,36-38], the enhancement of CLA using lactic acid bacteria (LAB) and bifidobacteria in fermented products can occur. Some reported criteria are responsible for the rate of rise in CLA relative levels: bacterial species and strains [37], type of milk and fermentation time [18,22], media supplementation with growth factors like Probiotics and diet fibers [38-40] and finally the rate of enzymatic reactions in biohydrogenation pathway during manufacture of Probiotic products [41].
Finally, from the data obtained in this study, it was possible to observe that the addition and the fermentation of bifido bacteria affected the distribution of fatty acids in the products. The main differences were ascribed between fermented bifido milk and control milk. Nevertheless, lipids extracted from unfermented milk showed some intermediate characteristics in fatty acids profile as compared to the control and fermented products that can corroborate the observations of Kim and Liu [40] who indicated that enzymatic reactions in the biohydrogenation pathway requires a specific rate to occur and maybe the simple addition of Probiotic in the product is not enough to activate this metabolic process.
Bioactive peptides: Bioactive peptides derived from milk protein are described to promote a lot of physiologic responses. Mostly, the milk proteins are latent until released and activated after digestive enzymes’ hydrolysis or through action of Proteolytic microorganisms in food processing [20]. These peptides have 3-20 amino acids (a.a) residues per molecule and its activity is related to a.a composition and sequence. Moreover, milk contains Angiotensin I-Converting enzyme (ACE-I) and cell modulation peptides that are important to anti-inflammatory actions [42].The Proteolytic system of lactic acid bacteria is well characterized. This system consists of a cell wall-bound proteinase and a number of distinct intracellular peptidases, including endopeptidases, aminopeptidases, tripeptidases and dipeptidases. Extracellular proteinases cause degradation of casein into oligopeptides. The long chain oligopeptides may be a source of bioactive peptides when further degraded by intracellular peptidases of lysed-lactic acid bacteria [20].
Table 2 shows the biopeptides profile presented in CM, UFBM and FBM during storage determined by HPLC LC-MS. The possible sequences and activities were compared with the literature [42].
| Product | MW of peptides detected by HPLC (Da) | MW of reported peptides (Da) | Probable sequence1 | Probable Function1 |
|---|---|---|---|---|
| CM | 391.1 803.5 |
391.1 802.5 |
Phe-Leu-Leu Lys-Ala-Val-Pro-Tyr-Pro-Gln |
ACE-I Cell Modulation |
| UFBM - D1 | 113.0 803.5 |
391.1 802.5 |
Phe-Leu-Leu Lys-Ala-Val-Pro-Tyr-Pro-Gln |
ACE-I Cell Modulation |
| UFBM - D7 | 391.1 803.5 |
391.1 802.5 |
Phe-Leu-Leu -Lys-Ala-Val-Pro-Tyr-Pro-Gln |
ACE-I Cell Modulation |
| FBM - D1 | 391.1 628.4 770.5 803.5 |
391.1 627.3 771.3 802.5 |
Phe-Leu-Leu Tyr-Leu-Gly-Tyr-Leu Ser-Arg-Tyr-Pro-Ser-Tyr Lys-Ala-Val-Pro-Tyr-Pro-Gln |
ACE-I Opioid agonist Opioid antagonist Cell Modulation |
| FBM - D7 | 391.1 451.7 628.4 770.5 803.5 |
391.1 904.4 627.3 771.3 802.5 |
Phe-Leu-Leu Thr-Val-Gln-Val-Thr-Ser-Thr-Ala-Val Tyr-Leu-Gly-Tyr-Leu Ser-Arg-Tyr-Pro-Ser-Tyr Lys-Ala-Val-Pro-Tyr-Pro-Gln |
ACE-I Antibacterial Opioid agonist Opioid antagonist Cell Modulation |
Table 2: Peptides detected by LC-MS HPLC in milk, fermented and unfermented milk, probable sequence and function according to literature.
The relatively easy combination of ESI with an ion-trap (IT) mass spectrometer has permitted many investigations of polar substances over a wide range of interests. This MS technique has permitted the characterization of a large number of peptides with antimicrobial, ACE-inhibitory and antioxidant activities, and phosphopeptides [43]. Several ACE-inhibitory peptides from milk proteins fermented with different lactic acid starters and hydrolyzed with digestive enzymes were identified by amino acid and MS analysis. It could be seen from Table 2 that ACE-I and cell modulation peptides are maintained in all products as biopeptides, i.e., CM, UFBMand FBM showed the same peptides even after 7 days of storage, underscoring that neither the presence of bifidobacteria nor the technology employed promote changes in that milk biopeptides.
Opioid peptides are commonly found in dairy products, it is know that food-derived opioid peptides are comparatively much more stable than the majority of endogenous opioid peptides to enzymatic degradation and usually possess weaker activity than endogenous peptides [44]. The fermentation process by bifidobacteria allowed opioids bioactive peptides formation. The peak 628.8, an agonist to selective opioid delta receptor are found in FBM, this bioactive peptide were an alpha-caseine exorphine. Some studies, including [45], demonstrated that food derived exorphines could enhanced memory consolidation and improved mental status by decreasing anxiety in mice models. However no data are available on opioid effects of casein, casein hydrolysate or milk in adult humans, which could be reduced to the presence of milk protein-derived opioid receptor ligands [45,46].
Moreover, it could be observed that cold stress after fermentation process increased the antibacterial peptide Thr-Val-Gln-Val-Thr-Ser- Thr-Ala-Val formation derived from kappa-casein [42,45] in FBM. To our knowledge this is the first time that this substantiation is reported in literature. Apart from that, Håkansson et al. [47] associated the increase in elaidic acid amounts (Figure 2A) to cleavage, as a cofactor, the α-lacto albumin of milk to liberate other antibacterial peptides [45,46].
The survival capacity of bifidobacteria and the production of bioactive compounds is a promising area of research. The potential health benefits of milk protein-derived peptides and fatty acids have been a subject of highest commercial interest to functional foods health-promoting functional foods.
Cold storage affected counts of B. Lactis HN019 in unfermented milk after seven days of storage. In the contrary, cold storage promoted probiotic adaptability, favoring better resistance to in vitro enteric digestion and subsequently to be delivered in appropriate amounts to colon. These data suggests that the probiotic resistance to in vitro gastrointestinal digestion is not just related to the technological process applied during food production, but the protection and adaptability of the matrix in resisting the digestion process.
Different technological process affected the relative contents of elaidic and conjugated linoleic acid in fermented milk prepared with B. Lactis HN019, but fermentation contributed to increase of these bioactive fatty acids relative content. As a consequence, both technological processes also ascribed a reduction of relative levels in linoleic acid.
The simple addition of B. Lactis HN019 in milk was not able to change the peptides profile of the milk. In contrary, fermentation process using B. Lactis HN019 increased bioactive peptides. Moreover, according to the literature, peptides formation by bifido fermentation is a new subject and most reports concerns data about yoghurt starters’ bacteria. In this study, it was clearly demonstrated that opioids, antagonist and agonist peptides were formed due to fermentation process, increasing the source of bioactive peptides. Finally, some peptides were modified during storage and increased the antibacterial activity of fermented milk. More studies are necessary in determining bioactive peptides during simulated in vitro digestion.
None declared.
Thiswork was supported by FAPESP (Fundação de Amparo a Pesquisa do Estado de São Paulo) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).The authors’ wish to thank Ana Beatriz Perez doAmaral for technical assistance.