Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2014) Volume 3, Issue 4
Four large groups of antibiotics were developed for different targets of bacterial cells which differ from those of human cells. Uncontrolled use of antibiotics, resistance soon followed. Today, the majority of bacterial infections are caused by multidrug resistant bacterial isolates and result in serious treatment difficulties. The main cause for antibiotic resistance is the acquisition of plasmids which carry antibiotic resistant genes. These extra-chromosomal genetic elements can be eliminated by phenothiazines from antibiotic resistant bacteria by the selective inhibition of plasmid replication over that of the plasmid carrying bacterium at three different levels or steps as follows: 1. replication of plasmid DNA is destabilized due to the drug induced relaxation of super-helical structure of replicative form of plasmid DNA; 2. inhibition of partition of plasmid DNA during the cell division blocks the rolling circle type of distribution of plasmid into the two daughter cells; 3. inhibition of re-infection of plasmidless bacteria by blocking conjugation. The medical significance of plasmid elimination in vitro provides a method to isolate plasmid-free bacteria for biotechnology without any risks of mutation and opens a new perspective in rational drug design against multidrug resistant bacterial infections.
<Keywords: In vitro elimination of plasmids; Antibiotic resistance; Metabolic; Colicin; Virulence plasmids; Promethazine; Plasmid elimination in vivo
Elimination of plasmids that carry antibiotic resistant genes renders the once infected bacterium susceptible to antibiotics to which the plasmid containing bacterium expressed resistance. Once the plasmid is eliminated, the infection can be treated with commonly available antibiotics which are inexpensive and safe. This new perspective makes “old” antibiotics useful for treatment of antibiotic resistant infections currently resistant due the presence of plasmids that carry antibiotic resistant genes.
Although the phenothiazine promethazine was initially selected and much studied for its in vitro antiplasmid effects because this compound is widely used in clinical practice and there are no contraindications when used in combination with other therapeutic compounds, other phenothiazines have been studied for potential anti-plasmid properties and some have been shown to express these properties in vivo. This review will discuss the development of phenothiazines for plasmid elimination for clinical use. During the last four decades the frequency of antibiotic resistant infections has continued to escalate. Due to this, many antibacterial chemotherapeutics are deemed to be ineffective globally and this, according to the recent WHO report [1] and presents a serious health problem that affects all countries of the world regardless of economic status (add new reference from the WHO published two weeks ago) There is thus a recognised urgent need for effective treatment alternatives for antibiotic resistant and multidrug resistant bacterial pathogens. In the herein review, the focus is on one of the mechanisms involved in multi-drug resistance of bacteria, namely, the presence of plasmids within the bacterial pathogen which carry antibiotic resistant genes that result in the resistance phenotype of bacteria. The mechanisms involved in plasmid replication and opportunity to block the plasmid replication in vitro and in vivo are mainstays of this review. Discussion of different resistance mechanism resulting from antibiotic resistance genes carried by the plasmids, e.g., antibiotic inactivating enzymes, membrane permeability or drug efflux, etc., are excluded from this short review.
Plasmids are extra-chromosomal self-transmissible DNA elements in prokaryotes and some eukaryotic cells. The most important functions of plasmids are related to antibiotic resistance, virulence, metabolic functions of pathogenic and commensal bacteria. The intracellular replication of bacterial chromosome and plasmids are basically different as it was first shown by Jacob et al. [2].
Plasmids are responsible for the self-transmissible antibiotic resistance of bacteria in nature and some of the bacterial plasmids are responsible for encoding virulence factors (from Bacillus anthracis to Mycobacteria) including tumorigenesis mediated by IV secretion systems, TGF etc in humans e.g Bartonella henselae, B. quintana, Helicobacter pylori and plants e.g. Agrobacterium tumefaciens [3]. A minireview was published by Nóra Gyémánt et al. [3].
Gene mediated metabolic effects of plasmids such as fermentation of sugars, nitrogen fixation in plants, haemolytic effects, etc., are also known for particular plasmids. Other important functions of specific plasmids are the coding for virulence factors (bacteriocins, haemolysins, toxins, etc.) that are responsible for the pathogenicity of Gram negative and Gram positive bacterial species. Tumor induction in man by Helicobacter pylori carrying genes that promote damage and inflammatory processes are well known and analogous to tumor induction in plants by plasmid carrying Agrobacterium or nitrogen fixation Rhizobium. Due to the spreading of antibiotic resistance, new compounds with antibiotic effects are urgently needed against common bacterial infections and tuberculosis as well As an alternative solution to combat plasmid mediated multidrug resistance of bacteria that cause serious infections and are therapeutically problematic, and often untreatable –we must find drugs that target plasmids. These we term antiplasmid compounds. The process by which antiplasmid compounds may effectively work is termed plasmid elimination.
At first, plasmids carrying antibiotic resistant genes were eliminated by acridines and ethidium bromide and phenothiazines in vitro. However, because these compounds are well known mutagens, they are excluded from in vivo use. In this review, the antiplasmid activity of phenothiazine drugs that are used in clinical practice every daybecause of their known property to intercalate to DNA much in the same manner as ethidium bromide and acridines- we have studied these compounds for potential antiplasmid effects [4-6]. During these studies, it was interesting to note that the growth of various Gram positive and Gram negative bacteria were also inhibited by several phenothiazines e.g chlorpromazine, levo-mepromazine, diethazine and promethazine both in vitro and in vivo [6]. The bactericidal effect was also shown on M. tuberculosis at 50 μg/ml; total destruction of the organism and loss of acid fastness in part of the cells were shown at 300 μg/ml phenothiazines [7]. Preliminary studies in mouse experiments had shown that the phenothiazine derivatives were moderately effective by comparing the mean survival time of M. tuberculosis infected mice. Mean survival times of the experimentally infected mice with M. tuberculosis No. 23296 were found to be between 45 ± 11 and 48 ± 12 days.
The plasmid elimination effect of ethidium bromide, acridine orange and widely used medicines such as the antihistaminic phenothiazine promethazine Pipolphen®) were compared [4-6]. In those experiments, chlorpromazine was found to be more effective than the acridines. For plasmid elimination we applied tricyclic compounds phenothiazines which were not mutagenic in vitro by the direct Ames test [8] and hence if effective, could be considered for in vivo human use. Of major importance is that these phenothiazine compounds were subsequently shown to possess antimutagenic properties. This was the beginning of a series of experiments in the direction of rational drug design applicable for elimination of antibiotic resistance- and other plasmids for possible use in the medical practice to treat antibiotic resistant infections or to produce plasmid free bacteria for basic biotechnology free of any mutagenic effects. Theoretically, antiplasmid compounds and antibiotics could have synergistic effects and could be recommended for the treatment of polyresistant infections without the risk of mutagenic effects.
Three phenothiazines such as promezthazine, chlorpromazine, levopromazine exerted a bacteriostatic, antiproliferative effect on Gram positive, Mycobacterium tuberculosis HRV37 bacteria at 10-60 μg/ml, and on Gram negative bacteria at 130-180 μg/ml concentration. Of the three compounds, chlorpromazine had a marked bactericidal effect on cultures of Bacillus anthracis growing in minimal medium. In addition, chlorpromazine had a significant bactericidal effect on resting cells of Escherichia coli suspended in saline. Pseudomonas aeruginosa however, was found to be resistant to phenothiazines. Experiments failed to derive phenothiazine resistant mutants from the highly sensitive B. anthracis.
An effective plasmid carrying R-factor elimination was observed at a chlorpromazine concentration of 50 μg/ml; the dose somewhat reduced the growth rate of multiple resistant E. coli in broth culture [5].
In these experiments we determine the effective phenothiazine structures responsible for F’lac plasmid elimination [9]. In another experiment, the inhibition of conjugation, the horizontal R-plasmid transfer from one cell to another was investigated in the presence of chlorpromazine derivatives. Plasmid-curing effect on different compatibility groups of the R-plasmids of E. coli was different [10-14].
Two metabolic derivatives of chlorpromazine, the chlorpromazine sulphoxide and 7,8-dioxochlorpromazine had no plasmid curing effect nor did methylene blue. However amitriptyline, 7,8-dioxochlorpromazine and acridine orange were inhibitors of the conjugal transfer of the resistance plasmid, R-144, whilst methylene blue, chlorpromazine sulphoxide, and imipramine had only slight effects on the horizontal plasmid transfer in vitro. We were therefore unable to demonstrate a simple correlation between curing ability and. inhibition of plasmid transfer amongst the psychoactive drugs tested. A mechanism of plasmid curing by surface action of the drugs was suggested as an alternative to the direct intercalation of the drugs into plasmid DNA. Furthermore it is known that acridines and chlorpromazine are able to intercalate into DNA in vitro. Consequently, as previously suggested [11-13], the mechanism of plasmid curing by phenothiazines might be based on an acridine-like intercalation into DNA and retransfer of the eliminated plasmids into the plasmidless daughter cells both in vitro and in vivo. The partitioning of plasmid into daughter cells was simultaneously blocked by inhibiting the septum formation in the dividing bacteria forming filaments in the culture and the intercalation were essential for the effective plasmid elimination at “sub growth inhibitory” doses. The inhibition of plasmid replication, partitioning and conjugal transfer of plasmid replication are essential requirements for in vitro and in vivo plasmid elimination. The procedure did not affect chromosomal replication; however, inhibition of DNAgyrase was shown. The curing effect was correlation of to the superdelocalizational ability of pi-electrons on the C-8, C-9b and N-atoms of phenothiazines [14-18].
Different kinds of plasmids were eliminated form different bacteria with phenothiazines and coordination complexes e.g., multiple antibiotic resistance, haemolysin B, colicine, and F’lac plasmid (Table 1) of E. coli, calcium -binding protein of Y. enterocolitica, tumor inducing plasmid of A. tumefaciens, and nitrogen fixation encoding plasmid from Rhizobium [6].
| Bacterial species | Elimination of |
|---|---|
| Acinetobacter anitratus | antibiotic resistance |
| Agrobacterium tumefaciens | tumor inducing ability in plants |
| Escherichia coli | antibiotic resistance and haemolysin B |
| Escherichia coli | lactose fermentation |
| Staphylococcus aureus | antibiotic resistance |
| Yersinia pseudotuberculosis | antibiotic resistance |
| Yersinia enterocolitica | virulence |
| Rhizobium meliloti | nodule formation, N-fixation in plants |
Table 1: Results of plasmid elimination in some bacterial species.
An example of the plasmid curing effect by a phenothiazine is presented by (Figure 1). Bacterial cells that produce dark blue colonies on EMB agar are cells that contain the F’lac plasmid. However when the F’lac plasmid carrying bacterial cells were cultured in the presence of a promethazine at sub- inhibitory concentrations for 20-24 hrs, the F’lac plasmid was eliminated from the fraction of the initial cell population and these formed pink colonies on the EMB agar plate [9].
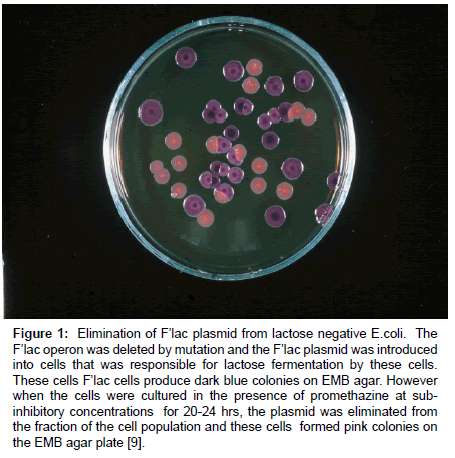
Figure 1: Elimination of F’lac plasmid from lactose negative E.coli. The F’lac operon was deleted by mutation and the F’lac plasmid was introduced into cells that was responsible for lactose fermentation by these cells. These cells F’lac cells produce dark blue colonies on EMB agar. However when the cells were cultured in the presence of promethazine at subinhibitory concentrations for 20-24 hrs, the plasmid was eliminated from the fraction of the cell population and these cells formed pink colonies on the EMB agar plate [9].
The plasmid curing effect of various compounds was dependent on the applied concentrations of curing compounds; pH and the incubation times.
It was interesting for us to note that different psychopharmacons with tricyclic structures had similar antibacterial effects on bacteria and plasmid replication. Some modification in chemical structures of phenothiazines (e.g.sulphoxide) resulted in the loss of plasmid curing effects. However when some naturally occurring “psychopharmacons or pain killers” e.g cannabis derivatives were studied, the hallucinogenic cannabinols did not affect plasmid replication and bacterial growth. Only the non-hallucinogenic terahydrocannabiolic acid was able to eliminate the bacterial plasmids. This compound had no effect on the CNS, and had very slight antibacterial effects. Tetrahydrocannabidiolic acid was the only agent that expressed plasmid curing effects [15]. Some charge transfer complex formation with the sub-molecular interactions were shown to be responsible for inhibition of plasmid replication [16-18]. As a result of our in vitro studies, acridines, ethidium, phenothiazines compounds, proton pump inhibitor trifluoromethylketones were shown to be effective in plasmid elimination of bacteria in vitro. But because acridines and ethidium are well known mutagens, these compounds would exert harmful side-effects on the host, and therefore these are excluded for their application in vivo. Conversely, certain medicines, e.g., tranquilisers and antidepressants used in everyday medical practice exert a similar antiplasmid effect similar to that of acridine in vitro. However the phenothiazines studied were not mutagenic and their copper, palladium and gold coordination complexes had a remarkable anti-mutagenic effectiveness [8] and antiplasmid effects [13] by shielding DNA base pairs.
The structure activity correlation was evaluated in a series of in vitro experiments. The pi-electron structure of each compound was determined by the Hückel method, and its correlation with the antiplasmid effect was studied by quantitative structure activity relationship (QSAR) multiregression analysis [11,13,17]. We found that the antiplasmid effect of different ring substituted phenothiazines correlates with the super-delocalization of the pi-electrons on atoms C-8, C-9b and N-10 of the phenothiazines [18,19].
On the basis of the found correlations, some phenothiazine and new anthryl- and phenanthril- compounds were synthetised with antiplasmid activity [19,20]. Phenothiazines and structurally related drugs were found effective in eliminating various bacterial plasmids [9,21]. Charge transfer complex formation between chlorpromazine and xanthine dyes with different electron acceptor activities (e.g. fluorescein, eosin, erythrosine and rose-bengal) was detected by differential spectrophotometry. Charge transfer complexes were formed between the strong electron acceptor rose-bengal and various tricyclic drugs as well. On the basis of the wavelength shift, the binding energies of drugs and dyes were estimated. Drugs with binding energies in the range of 0.23-2.31 kcal/mol can inactivate plasmid replication systems [15]. A serendipitous discovery led to studies on drugs affecting the central nervous system which also affect the expression of extra-chromosomal plasmids in bacteria. The drugs have a well-defined chemical structure, related electrochemical structures and tranquillizing effect in humans. Although most of the tricyclic drugs were unable to bind covalently to target cells in living systems, promazine was found to from complexes with various compounds, e.g., membranes and nucleic acids [2,3,11].
Tricyclic drugs interfere with energy dependent gyrase activity and promote the relaxation of plasmid DNA, resulting in a cessation in the R plasmid-gene expression and block plasmid replication. These results provide insight into the chemical structures connected with significant specific antimicrobial, so called antiplasmid effects [8,14].
A special complex formation was found with the covalently closed circular form of plasmid DNA, where GC rich regions of plasmid DNA have higher affinity for antiplasmid phenothiazines than adeninethymine rich regions of plasmid DNA. Because the G-C rich regions have a special role in the DNA stability via three hydrogen bonds, it was supposed that these regions could have a key role in the plasmid stability and replication [17,18].
Plasmid DNA becomes more rigid due to the intercalated phenothiazine molecules and strand breaks occur that lead to the formation of non-replicative forms open, circular or linear derivatives of CCC form (called covalently closed circular DNA). It is known that open circular forms or linear forms of CCC form of plasmid DNA cannot start to replicate from the origin of the promoter region due to the relaxed form of plasmid DNA [8,16]. Based on these observations new anthril compounds were synthetized [19].
It was surprising that some natural compounds isolated from essential oils of the peppermint oil exhibited antimicrobial and antiplasmid activities. The antiplasmid compounds were isolated and identified as menthol [22]. Their effect was comparable to the promethazine in vitro.
There are a number of pharmacologically active tricyclic compounds, many of which are in regular clinical use. Some of them have been demonstrated to exhibit various antimicrobial activities. They inhibit the growth of pathogenic bacteria, causing a general increase in membrane permeability, have antibacterial effects on Salmonella, Staphylococcus Escherichia coli and Yersinia enterocolitica cells [6,18,23,24]. It was shown that phenothiazines exhibited antibacterial affect in vitro and in vivo as well [5,6].
The role of charge distribution on the phenothiazines in the antiplasmid and antiviral effects was compared. The main aim was to design drugs without effects on the central nervous system – and this may open up a new way in the experimental chemotherapy.
The inhibition of plasmid replication and transfer can be the theoretical basis for decrease of bacterial plasmid replication in nature as well, where the gram negative bacteria can mediate the transmission of various plasmids in the nature. In this aspect there is a certain similarity between plasmids and viruses [18].
Before the introduction of plasmid curing in vivo, the phenothiazine compounds were selected and basic clinical investigations were performed e.g., the antibiotic sensitivity of individual bacteria from the urine of non-treated and treated patients were compared. In these experiments the urine or other biological samples (pus) were serially diluted and 100 μl of each dilutions were plated to nutrient agar and the plates were then incubated at 37°C to allow the growth a visible colony from each bacterial cell. The colonies grown on the master plates were transferred to antibiotic containing nutrient agar by using the Lederberg replica plating transfer method [25] and plasmid elimination from specific colonies readily demonstrated with clinical isolates from non-treated patients and patients treated with antibiotics plus phenothiazines [25-27]. These results gave rise to the possibility of a new type of combination therapy for antibiotic resistant bacterial infections mediated by then presence of plasmids carrying antibiotic resistant genes; the combination of traditional antibiotics with antiplasmid phenothiazines.
The antiplasmid action of phenothiazine and structurally related compounds was shown by elimination on the model F’lac plasmid of E. coli in model experiments. The first studies of successful plasmid elimination by promethazine in human patients were described in 1981-1982 [26-28].
In our former experiments, the conditions for plasmid eliminations were defined. For studies investigating the in vivo antiplasmid effects of a phenothiazine, the phenothiazine promethazine was selected because of its wide and safe use in everyday medical practice. This phenothiazine antihistamine and antiemetic has no remarkable side effects on the central nervous system as a consequence of symmetric pi-electron distribution in the skeleton of the phenothiazine. This phenothiazine was used to potentiate the antimicrobial effects of different antibiotics in various urogenital clinical infections in the Clinic of General Surgery at the Department of Nephrology, at University of Szeged [28]. Promethazine, - a phenothiazine-derivative – was given to patients presenting with a urogenital infection. This treatment resulted in the pathogenic bacteria in the urine to become sensitive to gentamicin and some other antibiotics to which they had been resistant before the administration of promethazine.
According to the phage-type, the bacteria before and after treatment with Pipolphen belonged to the same strain [26-28]. In addition to the R plasmid elimination, synergism between promethazine and gentamicin was also shown. Due to these results, we supposed that the membrane permeability of gentamicin could be enhanced by the presence of promethazine and Promethazine (Pipolphen®) was successfully applied in the clinical practice with 22 patients presenting with urogenital bacterial infections.
The pathogen was isolated from the first urine sample of the patients and the antibiotic sensitivity was determined. From the patients, 22 samples yielded resistant pathogens to 7 different antibiotics.
Of the 22 patients, 14 stayed in the clinic for 17 days and the patient’s urines were checked continuously every day for bacteria and the bacteria isolated were studies resistance toe gentamycin resistance. After 7 days of treatment all the patients were cured from the infection and recurrent infections were not observed. The mechanism of synergistic action of the drug interaction on cell membrane permeability was not studied in detail, but the disappearance of clinical symptoms and bacteriuria caused by gentamycin resistant Gram negative pathogens (E. coli, Klebsiella, Acinetobacter) was established [26-28].
Indeed tricyclic psychopharmacons contain not only a wide spectrum of agents that differ chemically and pharmacologically, but also in terms of interaction with various macromolecules of different cells. Among the well known phenothiazines, promethazine has been the most extensively studied. In a series of investigations promethazine has been administered to pregnant women for its antiemetic effect.
Finally, the effects of promethazine were studied in forty children with frequently recurring pyelonephritis aged 9.9 ± 4.2 years in the Pediatric Clinic at the Department of Nephrology at University of Szeged [29]. No significant difference in age and sex was between the 3 groups. None of the cases involved urological abnormalities. The patients were divided in three groups for the treatments. The results of three methods of treatment were compared:
For the in vivo studies we had 3 groups of patients.
In group 1 for combination therapy 10 children were given a combination of gentamycin and promethazine for 7 or 10 days. The daily dose of promethazine was 2 mg/kg orally, divided into two doses on days 1 and 2, and 1 mg/kg in one dose for the following 5 days. Gentamycin was administered i.m. in a daily dose of 4 mg/kg in two divided doses for the following (Group 1). The bacterial infections were successfully cured.
In the Gentamycin control group 11 patients received gentamycin treatment alone (i.m.). alone in the same dosage as in Group 1 for 10 days (Group 2).
In Group 3, 19 children received long term antibiotic prophylaxis with episodes of intensive treatments for recurrence for two weeks in a full dose of antibiotics according to the antibiotic susceptibility test (Group 3) were on long-term traditional oral antibiotic prophylaxis (5.6 ±2.1 years) with episodes of intensive treatment of recurrences. The number of recurrences in Group 1 was 3.
The number of recurrences in Group 2 was 27 during the 3 years: new scars were not observed in any of the patients from this group.
In a 3-year follow-up period, the number of pyelonephritis recurrences was significantly lower in Group 1 than in Groups 2 and 3. In the 3rd group, six out of 19 children had renal scarring. The authors suggest a synergistic effect between gentamycin and promethazine therapy. Promethazine increases antibiotic sensitivity, which could contribute to the elimination of recurring urinary tract infections [28,29].
In previous studies we reported that promethazine and other phenothiazines are able to eliminate the antibiotic resistance plasmid of E. coli in vitro and in vivo [23-26]. The antibiotic resistance plasmids allow the multiplication of bacteria in the presence of antibiotics which makes the antibiotic treatment insufficient. Plasmid curing was supposed to improve the opportunities of chemotherapy.
The practical importance of plasmid elimination in antimicrobial chemotherapy was supported by the studies where the plasmid curing effects of promethazine was induced in mixed bacterial cultures e.g. in the presence of S. epidermidis and B. cereus. The interactions between coexisting bacterial species did not reduce the frequency of plasmid curing by promethazine in mixed cultures with S. epidermidis or B. cereus in vitro [30].
In conclusion, we have demonstrated the role of plasmid elimination with combination chemotherapy by sensitizing the bacterial cells to antibiotics. Elevating the penetration of somle antibiotics into bacterial cells also can be a component to be considered in the rational drug design against multidrug resistant bacterial infections by the simultaneous application of antiplasmid compounds and antibiotics [31].
Antiplasmid effect of structurally similar maproptiline, amitriptilline to phenothiazines has also been shown in the patients. Based on on-going research in vivo studies, some new results will be reported in a forthcoming publication together with implication of combinations of antiplasmid compounds with antibiotics to improve the therapy of various acute and chronic bacterial infections.