Journal of Glycobiology
Open Access
ISSN: 2168-958X
ISSN: 2168-958X
Research Article - (2014) Volume 3, Issue 1
Keywords: Protein glycosylation; Mammalian cell culture; Product quality; Cobalt
2-AB: 2-Aminobenzamide; ADCC: Antibody Dependent Cell-Mediated Cytotoxicity; CDBM: Chemically-Defined Basal Media; CDFM: Chemically-Defined Feed Media; CHO: Chinese Hamster Ovary cells; Fuc: Fucose; FucT-Glycoprotein 6-α-L-fucosyltransferase; Gal: Galactose; GalT: β-N-acetylglucosaminyl glycopeptide β-1,4-Galactosyltransferase; GlcNAc: N-acetylglucosamine; GnTIII: β-1,4-Mannosyl-glycoprotein 4-β-N-acetylglucosaminyltransferase; Man: Mannose; pCO2: Dissolved Carbon Dioxide; PK: Pharmacokinetics; SiaT: β-Galactoside α-2,3/6 sialyltransferase; VCD: Viable Cell Density.
Protein glycosylation has been proven to be important towards the physiochemical characteristics of proteins and has been studied in considerable detail. Protein glycosylation is common to both mammalian and some non-mammalian species (e.g., yeast). However, it is the use of mammalian cells in culture which has enabled the generation of recombinant proteins that more closely resemble those present within humans. Therefore, mammalian cells are more frequently utilized for therapeutic protein generation. Amongst the various post-translational modifications, glycosylation has had one of the well-documented histories of having a direct impact on the resulting characteristics of the molecules they are attached to. Some of the noteworthy impacts glycosylation has been shown to be implicated in; include protein folding [1], solubility [2], immunogenicity [3-5], binding [6], stability [7] and pharmacokinetics (PK) [8,9]. For biopharmaceutical companies, the control of this is very important post-translational modification, thus paramount towards both ensuring acceptable product quality, as well as overall batch-to-batch comparability. The capability to control the addition of oligosaccharides onto a recombinant protein of interest is an important and sought-after endeavor, and will likely remain so for the considerable future.
There are two main types of protein glycosylation corresponding to the amino acid that the oligosaccharides are attached to Asn-linked (N-linked) glycosylation corresponds to the oligosaccharide attached to Asn residues. Ser/Thr-linked (O-linked) glycosylation corresponds to the oligosaccharide attached to Ser/Thr residues. In the case of protein N-glycosylation, the metabolic pathway is characterized through an initial enblock transfer of a pre-formed oligosaccharide onto select Asn residues of an antibody in the endoplasmic reticulum (ER) of mammalian cells. After a series of step-wise monosaccharide trimming reactions, a variety of Golgi and ER enzymes react upon the nascent N-glycan to generate a diverse range of potential N-glycan structures. The sum total of these various N-glycan structures is called the glycol form profile.
The biological pathway of N-glycan biosynthesis is associated with a series of converging and diverging reactions that has been described elsewhere [10]. The entire N-glycan biosynthesis pathway can be described by a relatively small number of enzymes, but with a correspondingly large number of potential N-glycan structures. Although glycoform diversity is typically observed in traditional cell culture processes, frequently there is a predominance of a few N-glycan species, with trace levels of a handful of others. This suggests some level of cellular or metabolic control over the metabolic pathway. Indeed, there have been numerous reports in the literature about the control points for protein glycosylation. These control points have been demonstrated to be at the enzymatic or substrate levels for the various reactions within the pathway. In addition, there have been numerous reports of the dependency of the resulting glycoform profile on the particular cells used for protein expression [11] as well as the corresponding process parameters and media conditions used for the cell culture process [12-16].
Control of glycosylation gene expression has been particularly effective towards the overall fine-tuning of glycoform profiles. This has been demonstrated in multiple instances including the overexpression of key enzymes such as GalT [17], SiaT [18-20] and GnTIII [21,22]. Gene silencing and gene knockouts have been demonstrated for the FucT enzyme leading to an overall reduction in core fucose levels [23,24], which has been proven to lead to overall increased ADCC levels [3]. At the substrate level, there have also been numerous studies that select media supplementation can facilitate dramatic changes in the resulting oligosaccharide profile. Some of those include the availability of sugars [25], amino acids and trace metals [26], as well as nucleotide-sugar precursors [27,28]. The cumulative evidence in the literature suggests that the rational gene manipulation or media supplementation of the principal factors of the glycosylation pathway typically results in the desired effect of modifying the glycoform profile towards the targeted glycan structure(s). This sensitivity towards the associated cell culture conditions is a primary reason that glycosylation is strictly monitored during the production of recombinant glycoprotein therapeutics.
Control of the protein N-glycosylation profile becomes particularly important towards the end of the pathway at the reaction steps that are frequently called terminal glycosylation (Figure 1). The addition of galactose through the activity of GalT helps extend and ensure for a more fully processed N-glycan. The enzymatic mechanism for this enzyme has been previously shown to be sensitive towards the presence of metal cofactors in purified enzyme studies, as well as in cell culture systems. Manganese in particular has been shown to be very effective towards catalyzing the reaction [29]. Cobalt has been shown to be partially effective in purified enzyme studies, but less effective than manganese. Cobalt was found to activate expression of the GalT enzyme in mung beans, but not as effective as manganese [30]. In studies with bovine derived enzyme, it was found that cobalt was significantly less active towards enzyme activation compared to manganese [31,32]. However, no prior precedent has been established for the selective use of cobalt as a mammalian cell culture media supplement for the specific enhancement of the GalT enzyme, and the resulting increase in a more fully galactosylated, and more fully processed N-glycoform profile on a recombinant protein therapeutic.
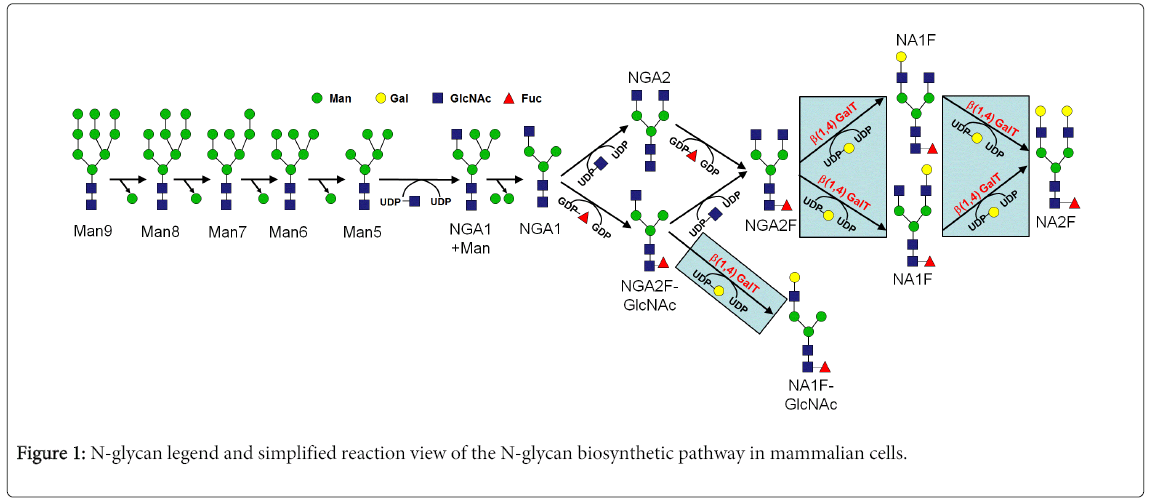
Figure 1: N-glycan legend and simplified reaction view of the N-glycan biosynthetic pathway in mammalian cells.
In the present work it has been found that the media supplementation of cobalt has resulted in a significant increase in N-glycan galactosylation. This observation is consistent with the enhancement of the N-glycan biosynthetic pathway at the GalT enzymatic reaction step, which results in the accumulation of these particular N-glycans. For some particular recombinant glycoproteins, a more fully galactosylated glycoform profile is a desired product quality attribute. This is especially the case when it is a precursor towards the formation of a more fully sialylated glycoform profile, which has been shown to facilitate a longer in vivo PK profile [33]. Thus, cell culture process conditions which bestow an increased N-glycan galactose content provide a clear benefit towards the generation of recombinant antibodies and other proteins where having a more fully processed N-glycan content is beneficial. The supplementation of cobalt in cell culture media also provides an effective and straight-forward means to target individual protein glycosylation profiles that might bestow a benefit towards some protein therapeutics.
Cell culture
Two recombinant Chinese Hamster Ovary (CHO) cell lines expressing two different recombinant glycoproteins were evaluated in 2 different culture vessels (shaker flasks and 3 L laboratory scale bioreactors). Cell Line 1 expressed monoclonal Antibody 1, and Cell Line 2 expressed Dual Variable Domain Immunoglobulin 1 (DVD1). Antibody 1 was an IgG1 glycoprotein, and DVD1 was an immunoglobulin glycoprotein with two variable domains as documented previously [34].
Both cells lines were of CHO DUX-B11 origin based on a dhfr (Dihydrofolate Reductase) expression system. All cultures utilized the same chemically defined basal media (CDBM), and chemically-defined feed media (CDFM), except Cell Line1 utilized a CDFM that was enriched at a 50% higher relative concentration. Each of the experimental conditions was supplemented with either cobalt chloride or manganese chloride (Sigma-Aldrich, St.Louis, MO) to evaluate their potential impact on the resulting N-glycan oligosaccharide profile. Concentrated glucose solutions were fed to the cultures as needed, when glucose levels dropped to sufficiently low levels. In preparation of the cultures, the cell lines were expanded through separate seed train inoculums to generate enough cell biomass for inoculation of multiple cultures. Process conditions utilized during the cultures were similar between each experimental condition and the respective non-cobalt supplemented control condition (Table 1).
| Cell Line 1 | Cell Line 2 | ||
|---|---|---|---|
| Culture Vessel | 250 mLshaker flask | 3L lab-scale bioreactors | 250 mL shaker flask |
| Culture Mode | Fedbatch | Fedbatch | Fedbatch |
| Initial Culture Temperature (°C) | 36 | 36 | 35 |
| Dissolved Oxygen (%) | N/Aa | 30-40 | N/Aa |
| pH Setpoint | N/Aa | 6.9 | N/Aa |
| CoCl2Concentrations (µM)b | 0, 1, 5, 25, 50, 100 | 0, 50 | 0, 1, 5, 25, 50, 100 |
Table 1: Summary of cell culture process conditions and cobalt supplementation details.
aCultures run in CO2 incubators at 5% CO2 in air; pH and DO parameters were not controlled, and thus did not have set point values.
bCoCl2 added to both chemically-defined basal and feed media at the same listed concentration.
Viable cell density (VCD) and cell viability values were measured through trypan blue exclusion via Cedex automated cell counters (Roche Applied Science, Indianapolis, IN), glucose and lactate values were measured with a ABL-805 (Radiometer Medical, Denmark) blood gas analyzer. Offline pH, dissolved oxygen (DO), and pCO2 measurements were also performed with an ABL-805 (Radiometer Medical, Denmark) blood gas analyzer. Osmolality was measured on a Multi-Osmette 2430 osmometer (Precision Systems, Natick, MA).
Protein A Affinity Chromatography
Antibody titers were measured from crude cell culture harvests on a Poros ATM (Life Technologies, Carlsbad, CA) affinity column using an HPLC system operating with a low pH, step elution gradient with detection at 280 nm. Absolute concentrations were assigned with respect to reference standard calibration curves.
Purified antibodies subjected to additional analytical characterization were purified using MabSelectTM Protein A (GE Healthcare, Piscataway, NJ) using a low pH, step elution gradient, followed by buffer exchange using Spin Concentrator X UF columns (Corning Lifesciences, Tewksbury, MA), or equivalent, according to the manufacturer’s recommended procedure.
N-glycan Oligosaccharide Profiling
Approximately 200 µg of Protein A purified samples from Cell Lines 1 and 2 were treated with N-glycanase at 37°C for an overnight duration to remove the N-glycans from the protein. The protein was precipitated and the supernatant was taken for subsequent chemical derivatization of the reducing end of the released glycans with 2-aminobenzamide (2-AB) dye. Following the derivatization step, the excess dye was removed using clean up cartridges and the samples were analyzed using normal phase HPLC with fluorescent detection. Mobile phase A was 100% acetonitrile and mobile phase B was 50 mM ammonium formate at pH 4.4. The glycans were eluted from a polyamide column (Prozyme, Hayward, CA) using a shallow gradient. The labeled glycans were detected using a fluorescence detector with an excitation wavelength of 330 nm and an emission wavelength of 420 nm.
Experimental results are expressed as mean ± 1 SD for those results generated from at least 3 independent cultures. Experimental results are expressed as the mean value for those results generated from less than 3 independent cultures. Results were evaluated for statistical significance (when needed) through 2-sided t-tests, with a requirement of p<0.05 relative to the unsupplemented control conditions.
Impact of Cobalt on the Protein Oligosaccharide Profile of a Recombinant Antibody
Cell Line 1 was cultured in shake flasks in fed batch mode after an abbreviated seed train. Various concentrations of cobalt chloride were supplemented into both the CDBM and CDFM at the same concentration in each media and evaluated for the resulting impact on cell culture process performance (Figure 2). Viable cell density profiles for the cobalt supplemented cultures trended similarly compared to the unsupplemented control, with the 1, 5, and 25 μM concentrations facilitating an approximately similar peak VCD compared to the control. The culture flasks supplemented with cobalt to 50 μM and 100 μM concentrations facilitated a lower cell growth achieving a peak VCD of approximately 9.4x106 cells/mL and 7.0x106 cells/mL, respectively. The largest statistically significant differences in cell growth compared to the unsupplemented control were realized at the highest cobalt tested concentration of 100 μM. Cell viability profiles across all conditions, either with cobalt, or without, were also comparable up to the point of culture harvest on Day 14. In each of the cobalt concentrations evaluated, they all supported slightly higher cell viability at harvest compared to the unsupplemented control. These results suggest that cobalt alone does not have a very significant impact on cell growth over the lower end of the range of concentrations evaluated, but at concentrations higher than 50 μM it is anticipated that larger drops in VCD should be apparent.
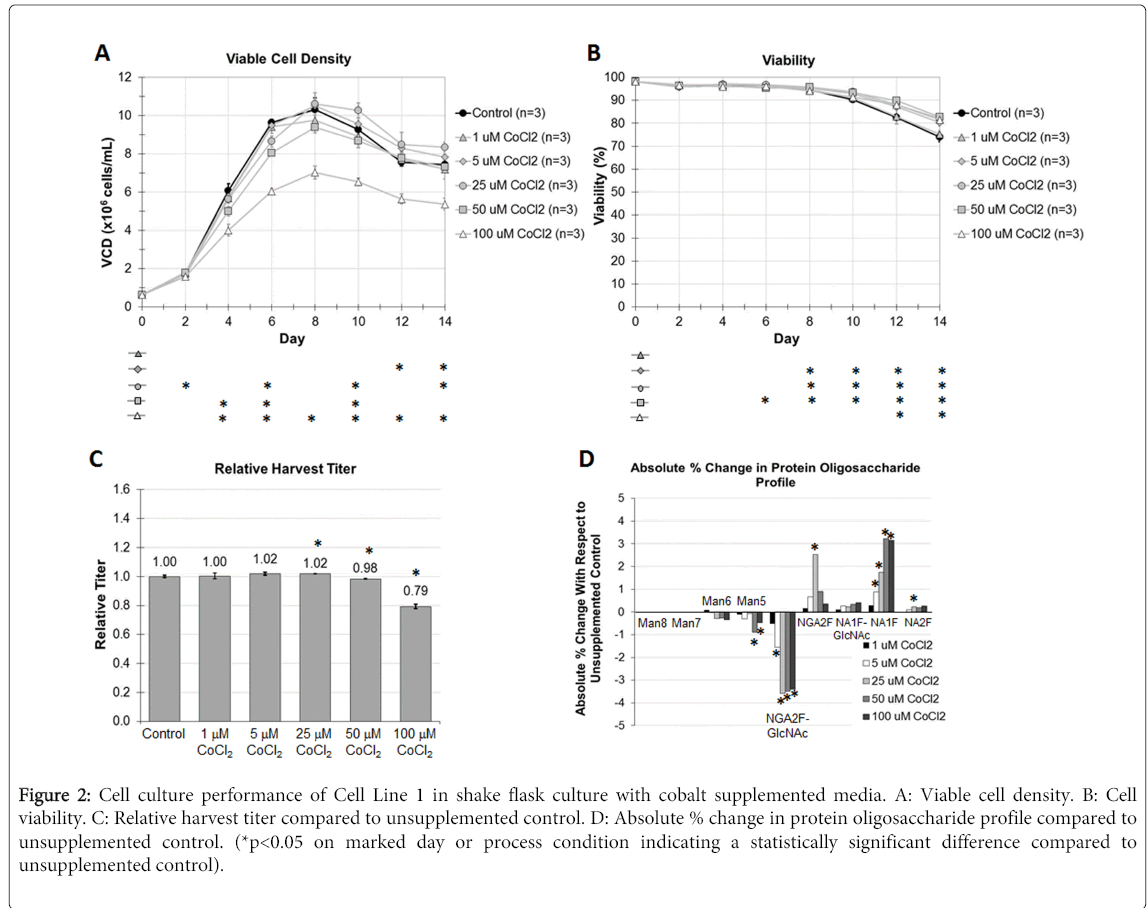
Figure 2: Cell culture performance of Cell Line 1 in shake flask culture with cobalt supplemented media. A: Viable cell density. B: Cell viability. C: Relative harvest titer compared to unsupplemented control. D: Absolute % change in protein oligosaccharide profile compared to unsupplemented control. (*p<0.05 on marked day or process condition indicating a statistically significant difference compared to unsupplemented control).
Harvest titers of Antibody 1 mirrored the cell growth results. For the most part there was a negligible impact on recombinant antibody productivity across the 1, 5, 25, and 50 μM cobalt concentrations evaluated. However, there was a 21% drop in productivity in the 100 μM cobalt concentration condition, which was also statistically significant compared to the control.
The N-glycan oligosaccharide profile demonstrated a clear trend which was also concentration-dependent on the amount of cobalt supplemented into the media. The higher the CoCl2 concentration, the larger the absolute percent increase in NA1F species. NA2F species also increased although to a much lower relative amount. This increase came at the expense of the lesser processed N-glycan species including NGA2F-GlcNAc, which saw statistically significant decreases in relative abundance. These measured results of higher G1/G2 N-glycan species is consistent with the purported mechanism of cobalt being a cofactor for the GalT enzyme, and although it is not the preferred cofactor in terms of reactivity, it still is capable of facilitating enzymatic activity. This is the first demonstration of this effect in a mammalian cell culture system expressing a recombinant biotherapeutic. Adding cobalt into cell culture media for the targeted enrichment of galactosylated N-glycans is a quick and easy means for fine tuning N-glycoform profiles, and creates a means towards ensuring biologic comparability through process change.
Impact of Cobalt on the Protein Oligosaccharide Profile of a Dual Variable Domain Immunoglobulin
Cell Line 2 was cultured in shake flasks in fed batch mode after an abbreviated seed train. Various concentrations of cobalt chloride were supplemented into both the CDBM and CDFM at the same concentration in each media and evaluated for the resulting impact on cell culture process performance (Figure 3). Viable cell density profiles for the cobalt supplemented cultures trended similarly compared to the unsupplemented control, with the 1, 5, and 25 μM concentrations facilitating an approximately similar peak VCD compared to the control. The culture flasks supplemented with cobalt to 50 μM and 100 μM concentrations facilitated a lower cell growth achieving a peak VCD of approximately 10.6x106 cells/mL and 6.7x106 cells/mL, respectively. The largest statistically significant differences in cell growth compared to the unsupplemented control were realized at the highest cobalt tested concentration of 100 μM. Cell viability profiles across all conditions, either with cobalt, or without were also comparable up to Day 6. After this, the cell viability profiles began to diverge, with the higher cobalt media concentrations supporting higher cell viability over time, as well as at harvest. These results suggest that cobalt alone does not have a very significant impact on cell growth over the range of concentrations evaluated, but at concentrations higher than 50 μM it is anticipated that larger drops in cell growth performance should be apparent.
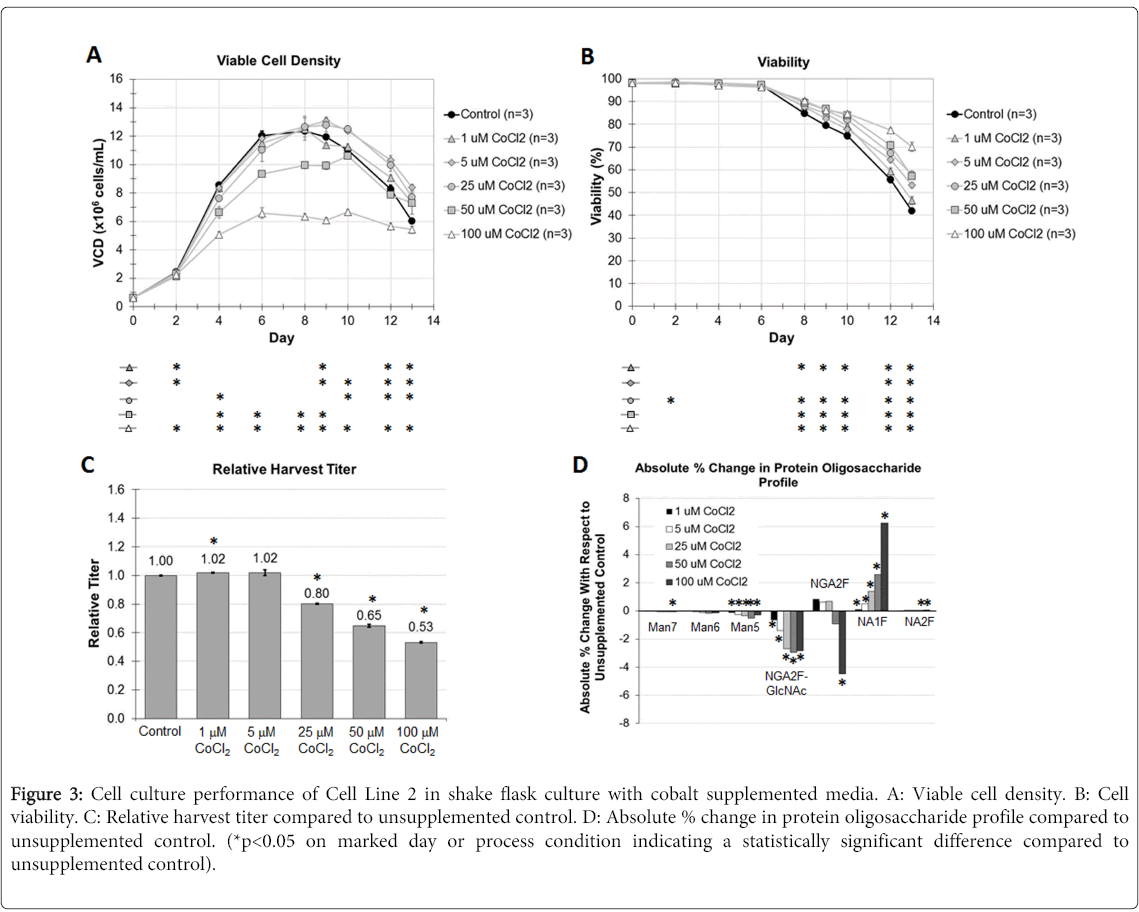
Figure 3: Cell culture performance of Cell Line 2 in shake flask culture with cobalt supplemented media. A: Viable cell density. B: Cell viability. C: Relative harvest titer compared to unsupplemented control. D: Absolute % change in protein oligosaccharide profile compared to unsupplemented control. (*p<0.05 on marked day or process condition indicating a statistically significant difference compared to unsupplemented control).
Harvest titers of DVD 1 mirrored the cell growth results. For the most part there was a negligible impact on recombinant antibody productivity across the 1 and 5 μM cobalt concentrations evaluated. However, there was a 20%, 35%, and 47% decrease in productivity across the 25 μM, 50 μM, and 100 μM cobalt supplemented cultures, which was statistically significant compared to the control.
The N-glycan oligosaccharide profile demonstrated a clear trend which was also concentration-dependent on the amount of cobalt supplemented into the media. The higher the CoCl2 concentration, the larger the percent increase in NA1F species. The 100 μM cobalt condition facilitated a 6.2% increase in NA1F glycans, for example. NA2F species also increased although to a much lower relative amount. This increase came at the expense of the lesser processed N-glycan species including the high mannose glycans, as well as NGA2F-GlcNAc, which saw statistically significant decreases in relative abundance. These measured results of higher G1/G2 N-glycan species are consistent with the purported mechanism of cobalt being a cofactor for the GalT enzyme, and although it is not the preferred cofactor in terms of reactivity, it still is capable of facilitating enzymatic activity. Adding cobalt into cell culture media for the targeted enrichment of galactosylated N-glycans is a quick and easy means for fine tuning N-glycoform profiles, and affects a means towards ensuring biologic comparability. In the demonstrated examples it is also readily apparent that the impact of media supplementation of cobalt is very consistent across a variety of different recombinant protein types.
Laboratory-Scale Bioreactor Confirmation of the Targeted Modulation of Protein Glycosylation Profiles through Cobalt
3 L scale down model bioreactors were utilized to verify the impact of CoCl2 on the resulting protein glycosylation profiles in Cell Line 1. For comparison purposes, MnCl2 was also evaluated since this particular trace metal has already been established to impact the oligosaccharide profile towards higher galactosylated species. Cell culture process performance indicators were monitored and measured throughout the respective cultures. Viable cell density, cell viability, harvest titer, and harvest N-glycan oligosaccharide data was measured and reported. Figure 4 highlights the cell culture performance results observed through the use of 50 μM CoCl2 and 1 μM MnCl2 targeted supplementation into the CDBM and CDFM.
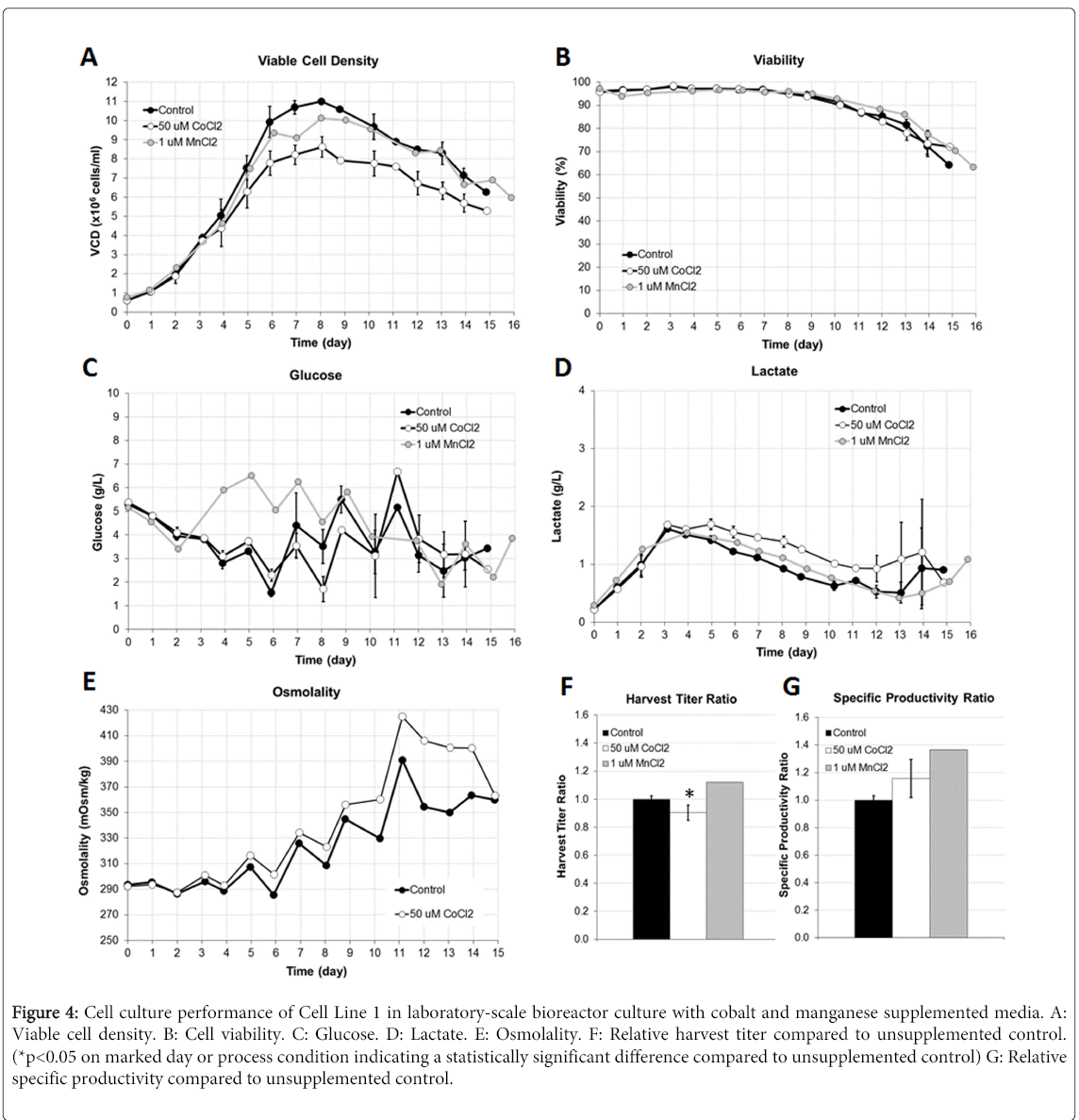
Figure 4: Cell culture performance of Cell Line 1 in laboratory-scale bioreactor culture with cobalt and manganese supplemented media. A: Viable cell density. B: Cell viability. C: Glucose. D: Lactate. E: Osmolality. F: Relative harvest titer compared to unsupplemented control. (*p<0.05 on marked day or process condition indicating a statistically significant difference compared to unsupplemented control) G: Relative specific productivity compared to unsupplemented control.
Similar to the shake flask results, there was a modest drop in the viable cell density profile, but no change in the viability profile in the 50 μM CoCl2 supplemented culture. The 1 μM MnCl2 supplemented culture also demonstrated a modest, but measurable drop in VCD, with no apparent impact on cell viabilities over time. Glucose and lactate levels in the culture media were approximately similar to each other over time, with the only differences observed with the MnCl2 supplemented culture which had an earlier glucose addition, and in the CoCl2 supplemented culture which demonstrated a slightly higher overall lactate with at most a 0.5 g/L higher concentration, on average. Osmolality levels were also slightly elevated in the CoCl2 supplemented culture compared to the unsupplemented control.
Similar to the shake flask results using 50 μM CoCl2, there was a nominal drop in the final harvest titer with the cobalt supplemented culture achieving a final harvest titer 10% below that of the unsupplemented control. The decrease in harvest titer of Antibody 1 was likely caused by the drop in viable cell density through the use of this relatively high concentration of cobalt. The MnCl2 culture did not demonstrate as large a drop in cell growth, and its final harvest titer was actually higher than the unsupplemented control by a nominal amount. As a result of the aforementioned data, the calculated cell specific productivity of the CoCl2 and MnCl2 supplemented cultures was higher than the unsupplemented control.
The N-glycan oligosaccharide species were measured in these samples and the results shown in Figure 5. The results suggest that through the use of CoCl2 there was an approximate 8% increase in NA1F species, which came at the expense of the less fully processed NGA2F-GlcNAc, NGA2F, and Man5 species, with all results being statistically significant. In the MnCl2 supplemented culture there was an even higher increase in NA1F species of 25% and NA2F species of 3%, which also came at the expense of the less fully processed N-glycans. These results outpaced that of the CoCl2 cultures, and highlight that Mn is without question a potent and primary cofactor towards the GalT enzyme, but Co is also effective, and its supplementation in cell culture media provides another means with which to effect significant changes in the protein glycosylation profile. It can be reasonably concluded that the higher levels of NA1F and NA2F measured in these trace metal supplemented cultures is the result of the trace metals themselves, and not the result of a lower cell specific productivity, since this calculated parameter was actually higher in the trace metal supplemented cultures. It can also be concluded based on the above data that this effect of cobalt on NA1F and NA2F species can be considered to be process scale-independent.
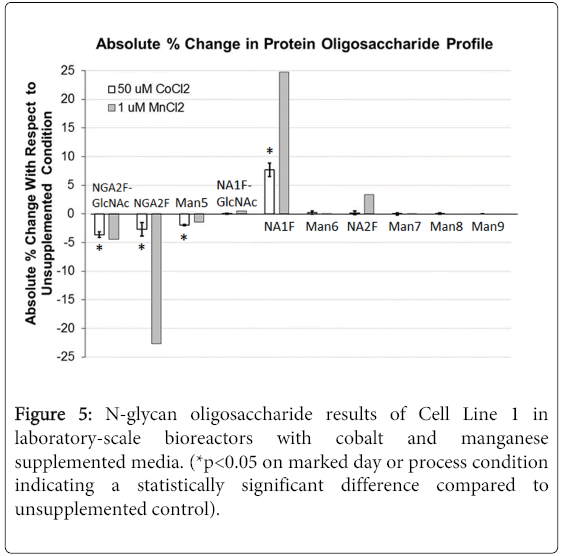
Figure 5: N-glycan oligosaccharide results of Cell Line 1 in laboratory-scale bioreactors with cobalt and manganese supplemented media. (*p<0.05 on marked day or process condition indicating a statistically significant difference compared to unsupplemented control).
It is interesting that with both trace metals there is an increase observed in NA1F species, but not much with the NA2F species, regardless of the magnitude of the increase in the NA1F species. A likely reason for this is the branch specificity of the GalT enzyme and the ungalactosylated N-glycan substrate. That is, it has been shown previously using purified enzyme from rat liver that the GlcNAcβ(1,2)-Manα(1,3) branch of biantennary glycans is a more preferred N-glycan substrate for the GalT enzyme compared to the GlcNAcβ(1,2)-Manα(1,6) branch [35]. The resulting effect of this substrate specificity is an enrichment of one type of NA1F type glycan that predominates the total galactosylated N-glycan oligosaccharide profile.
In the present work it has been found that the selective use of cobalt is an effective approach in mammalian cell culture for the targeted shifting of protein glycosylation profiles. Cobalt was found to facilitate an increase in NA1F species in a statistically meaningful way, increasing the overall levels of galactosylation, and thus the extent of terminal N-glycan processing. It is highly likely that this effect of cobalt is enabled by elevating the activity of the GalT enzyme through its role as a cofactor for the enzymatic reaction shown in Figure 1. This behavior was observed across multiple concentrations in a concentration-dependent manner. It was further found that up to a particular concentration there was a negligible impact on cell growth, at which point an adverse impact on cell growth began to be apparent at relatively higher concentrations. For Cell Line 1 this concentration was 50 μM and for Cell Line 2 this concentration was 25 μM. In addition, it was found that up to a particular concentration there was a negligible impact on recombinant protein productivity. For Cell Line 1 this concentration was 50 μM and for Cell Line 2 this concentration was 5 μM. At higher concentrations than these, the harvest titers began to drop for those particular cultures. It is apparent that individual mammalian cell lines have different sensitivities towards cobalt and how much they can or cannot tolerate without being impacted in a negative way.
However, what appears to be common is that in each instance cobalt is able to increase the relative levels of galactosylated N-glycans. For Cell Line 1, the maximum absolute percentage increase was observed to be 8% relative to the unsupplemented control in 3 L scale bioreactors. For Cell Line 2, the maximum absolute percentage increase was observed to be 6% relative to the unsupplemented control in shaker flask cultures. These afore mentioned results of cobalt facilitating an increase in galactosylated N-glycans were consistent across the volumetric scales of the cultures, as well as the protein expressed. Since cobalt is typically in most cell culture media at trace levels, its supplementation is a quick and easy way to improve a cell culture process where the extent of N-glycan galactosylation concerns are apparent. The targeted increase in galactosylation is indeed an important capability since the addition of galactose must proceed before the addition of sialic acid can occur which has been well documented towards increasing in vivo PK. In addition, cobalt supplementation provides another tool towards ensuring biologic comparability, the targeted optimization of product quality, and the empowerment towards ensuring the expressed protein will meet pre-defined release specifications.
Analytical testing from Yu Zhou and Haly Raharimampionona from the Protein Analytics group within Process Sciences at AbbVie Bioresearch Center, Worcester, MA.
All authors and acknowledged contributors were employees of Abbott Laboratories until 2012, and were employees of AbbVie Inc. starting in 2013, and were employed at these companies during the data generation for this work. Financial support for performing this work was provided by Abbott (until 2012), and AbbVie (starting in 2013). The authors and acknowledged contributors were responsible for experimental design, data collection, analysis, and interpretation, as well as the writing of this work. AbbVie was responsible for this work’s approval. All authors and acknowledged contributors have a financial interest in Abbott and/or AbbVie, and have no professional or personal conflict of interest to declare.