Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2016) Volume 4, Issue 1
A diverse, symbiotic ecosystem of microbes resides in our gut, contributing to the complexity of human health. As the most microbe rich area of the human body, the gut microbiota provides a number of important physiological functions including metabolism, immunity, and protection from pathogens. Environmental factors, especially nutrition and dietary-components, can influence or even completely alter the microbial landscape and its functions. Currently, it is thought that under certain, but unknown, genetic and environmental contexts these changes can cause or exacerbate chronic inflammatory diseases. While using probiotics to treat disease seems like an easy solution, both basic and clinical data have demonstrated mixed results. Thus, it is imperative to re-examine probiotics in the complex context of both a healthy and diseased microbiome along with associated factors such as diet.
Keywords: Gut microbiota; Nutrition; Probiotics; Targeted therapy
The human body is home to trillions of microorganisms, each with functions affecting the microenvironment of our bodies. From the different areas of the skin, to the start and finish of gastrointestinal (GI) tract, robust and vastly diverse communities of microbes thrive. While the human skin alone harbors microbes from 19 out of 100 different bacterial phyla [1,2], the human gut contains only 7 [3], yet is home to 100 trillion microbes collectively referred to as our microbiota [4]. With these microbes, our GI tract represents a complex, multifunctional organ that is not only associated with digestion, but also with immunity, metabolism and resistance to pathogenic infection. The intestine contains multiple and unique physical, chemical, and structural features that define its anatomical landscape. These dramatically different, yet adjacent, geographies allow the colonization of microbes expressing certain genetic or virulence traits that allow them to survive. For those microbes not already expressing the necessary genes for colonization they will not survive or they will mutate and adapt to the microenvironment. Thus, the 9 million genes expressed by our microbiota [5] play important roles in helping to define the microbial diversity of our GI tract as well as contributing an enormous amount of to our own genetic diversity [5].
Early microbiota
The gut microbiota interacting with the human host is a unified super-organism with stable yet evolving features. In utero we begin life as a sterile entity. Microbial colonization begins during birth, with one profound initial colonization event occurring during delivery (Figure 1). Studies have compared the microbial environments resulting from vaginal births to those resulting from Cesarean births, finding that infants born via a vaginal birth acquire a microbial phenotype dominated by probiotic Lactobacillus species but have less overall diversity, or fewer bacterial types [6]. In contrast infants born via Cesarean section have a greater overall diversity as their intestinal colonization much more resembles the skin microbiome which also often has lower levels of lactobacilli [6]. These early colonization events from a direct transfer of microbiota from the mother has important consequences for the overall health of the child [7].
Complicating these studies is the nutritional status of the infant and whether they received a formula-fed diet or were nursed on their mothers’ breast milk. In the latter case, passive transfer of secretory immunoglobulin A (sIgA) not only helps to protect an infant from potentially infectious organisms, since the development of its own immune system takes many months, but sIgA also controls the interaction of the infants’ cells with its newly acquired microorganisms [8]. Additionally, human breast milk contains essential ingredients for a healthy gut microbiota including carbohydrates, short chain fatty acids (SCFAs), and lactoferrin [9]. Breast milk may also contain viable microbes capable of influencing the colonization of the infant gut microbiome. In a study comparing the microbiota of 20 vaginally delivered infants and the composition of their mothers’ breast milk, Solis et al. [10] report viable lactobacilli and Bifidobacteria species capable of vertical transfer to the infant gut microbiota. A longitudinal study by Fallani et al. [11] further highlights the influences of infant feeding patterns on the intestinal microbiota. Breast-fed infants have a Bifidobacteria dominated microbiota while formula-fed infants have a more diverse microbiota including lower counts of Bifidobacterium and higher counts of Bacteroides and Clostridium [11]. Likewise, in a review by Thum et al. [12] the benefits and possibility of modifying infant microbiota and overall health through maternal diet are discussed.
While these studies demonstrate the influences of maternal probiotic bacteria and immunity on the infant microbiota, there remains a lack of understanding of the interplay between these factors in the short and long term health of infants. For instance, comparing the evolution of the infant microbiota with the interaction of delivery mode with feeding conditions have not been well described as most studies compare only mode of birth or feeding practice with vaginallydelivered infants. By analyzing the changes in the microbiota of a child delivered vaginally and formula-fed to a child born by Cesarean fed breast milk could provide critically important data regarding the influence of the microbiota by environmental factors. Perhaps breastfeeding has a stronger influence on the microbiota, leading to a convergence of microbiota patterns despite mode of birth. Or maybe the initial mode of birth results in a lasting variation of the microbiota that may later influence the immune responses. Thus, it is important to consider how environmental factors including nutritional conditions and initial exposure to skin or vaginal microbiota communicate to develop the infant microbiota.
Adult microbiota
During early colonization, the unique microbial composition of each child converges into the core microbiome of an adult [13]. Once established the microbial community is stable, but not fixed, influenced by environmental and genetic factors (Figure 1) [13]. These influences provide a variety of phylum proportions throughout the human body and variability among humans [14]. Similar to the influence of nutrition on an infants’ microbiome, diet and antibiotic consumption are essential in shaping the adult microbiome. For example, mouse models with high fat diets and fecal microbiota transplants show clear evidence of the gut microbiota being indicative and causal of a physical phenotype such as obesity and leanness [15-18]. Among humans, one controlled-feeding study showed a strong microbiota profile, or enterotype, associated with long-term diets [18,19]. Mouse models clearly demonstrate that genetically obese individuals develop a unique gut microbiota [15]. In a pioneer study by Ley et al. [15] in which obese mice were shown to have a microbiota highly dominant in bacteria of the Firmicutes phylum, a more balanced ratio between Firmicutes and Bacteroidetes was observed in lean mice [15]. At the genus level, Bacteroidetes species have been positively associated with lean individuals, [20] while Lactobacillus has been negatively associated with lean individuals [20]. Even species-specific benefits have been shown to influence obesity. In a study analyzing Lactobacillus and Bifidobacterium species in obese individuals, Million et al. [21] identified an association of B. animalis , L. paracasei , and L. plantarum with normal weight and L. reuteri with obesity. These studies clearly outline a definitive microbiota in obese individuals capable of altering phenotypic, and possibly health, condition.
In addition to the environmental factors constantly contributing to shifts of the microbiome, the human host affects the microbiome through one’s own genetic background. Genetic studies of both human twin subjects and mouse models reveal genes associated with specific bacterial abundances and composition. Goodrich et al. (22) establishes this connection using 416 twin-pairs and over 1,000 fecal samples; monozygotic twins had greater microbiota similarities than dizygotic twins [22]. In another study, Davenport et al. [23] incorporates genome-wide association studies with microbiota sequencing using a founder population to limit variation in environmental exposure. Despite a small sample size, at least 8 bacterial taxa were significantly related to human genetic variation [23]. The mechanism of hostgenetic modification of the microbiome remains unclear [24], however proposed models include alterations in levels of hormone production, energy availability, and immune system interaction [23]. Studying the genetic predisposition is crucial for understanding the evolving microbiota. While studies using adult twins or related family members provide a human model, these studies cannot control the variable environmental factors. Therefore, genetic influences are easily confounded by environmental influences. The use of genetically engineered mouse models manipulated under controlled environmental conditions can help us begin to understand the interplay between environment and genetics, but the translatability of these studies to humans still remains an issue.
Microbiota: Initiator of disease and target of treatment
Most notably, changes within the gut microbiota have been associated with a number of chronic inflammatory diseases. These changes can occur in the composition, location, and/or function of the microbiota and are referred to as dysbiosis [25]. With over 60% of immune cells residing in the intestinal mucosa, the GI system and the microbiome is one of the leading influences on the immune system [26]. As the initial colonization of an infant’s GI tract begins, so does the development of the immune system [27]. Furthermore, the gut microbiota is in constant contact with the human host and will continue to influence the immune system throughout life. Due to an association between microbiota shifts and diseases, the gut microbiota is connected to a range of conditions such as obesity, cardiovascular disease, and inflammatory bowel disease [13]. In animal studies, the gut microbiota is shown to have important effects on metabolic diseases with causal effects being reported on glucose and lipid metabolism, adipose development, and insulin signaling [1,28].
The dynamic nature of the microbiome makes it a desirable target for potential therapies to prevent or treat disease. An obvious choice for such development is the administration of probiotics. Probiotics, as defined by the FDA and WHO, are “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [29]. As the body is already populated with several millions of microbes, the idea of probiotic therapy would be to supplement the existing microbiota with “beneficial” bacteria. The health benefits of these bacteria would interact directly with the microbiota via the competition with potential opportunistic commensals (“pathobionts”) or infectious microorganisms for nutrients and/or anatomical niches. In fact, studies of the physicochemical and adhesive properties of Bifidobacterium and Lactobacillus species have been instrumental in revealing possible strains for probiotic use due to their ability to inhibit adherence of enteric pathogens such as Shigella, Staphylococcus, Salmonella, and Listeria [30,31].
Probiotic bacteria may also modulate the microbiota indirectly through inhibiting commensal translocation by decreasing intestinal permeability [26] or promoting anti-inflammatory activity of the immune system [25]. For example, Yan and Polk [32] observed decreased apoptosis and increased survival of colonic cells cultured with L. rhamnosus GG. L. rhamnosus GG produces two soluble proteins capable of regulating apoptosis and proliferation in intestinal epithelial cells [33]. The latter is accomplished through the production of SCFAs [34] and anti-inflammatory cytokines such as IL-10 and TGF-β [35]. In addition, dietary constituents influence whether the microenvironment of the GI tract is permissive for the establishment of pathobionts or the promotion of probiotic bacteria. For example, diets high in fiber have been shown to increase bacterial production of acetate and butyrate [36] which is beneficial for the host because butyrate produced through the fermentation by probiotic bacteria induces colonic regulatory T cell differentiation and enhances immune-suppression and tolerance [36]. On the other hand, diets high in fat create low grade inflammation [37] that promotes the expansion of pathobionts, reduces probiotic species [37], and has been suggested to affect commensal translocation [38]. While the presence of probiotic bacteria may inhibit the adherence of pathogenic bacteria, studies examining the relationship between nutritional status, pathogenic infection, and probiotics have not been well studied. These results demonstrate the ability of probiotic species of the microbiota and their byproducts to directly and indirectly affect the intestinal microenvironment but currently lack mechanistic studies and translation to human disease.
Microbiota: Probiotic interventions
The interaction between nutritional status and probiotics is highlighted in research of metabolic diseases and associated syndromes. Several studies focusing on metabolic diseases have positive outcomes using probiotic therapy. One mouse study using high fat diet-induced diabetes found probiotic treatment highly effective in improving glucose tolerance, restoring insulin sensitivity, and reducing inflammatory cytokines in adipose tissue [38]. These results demonstrate clear evidence that a probiotic, capable of preventing bacterial adherence and translocation, can reverse a dietinduced intestinal disturbance and adipose inflammation [38]. In a double-blind human study of males with type 2 diabetes, use of a probiotic improved insulin sensitivity while baseline inflammatory markers remained the same [39]. Probiotic therapy has also shown success in a number of studies when used to treat chronic inflammatory diseases. For instance, clinical trials using B. infantis either mixed with a malted milk drink or in an encapsulated form both had promising results in relieving irritable bowel syndrome (IBS) symptoms, including decreased pain/discomfort, bloating, and bowel movement difficulty [40,41]. Molecularly, Pathmakanthan et al. [42] identified increases in IL-10 production and increases in the numbers of T cells and macrophage from the blood of healthy patients and colonic mucosa of active ulcerative colitis patients treated with L. plantarum . Another in vitro study using human monocyte-derived dendritic cells found that L. rhamnosus inhibits proliferation of T-cells and decreases IL-2, IL-4, and IL-10 production [43], results that were replicated in human subjects with Crohn’s disease, indicating a potential L. rhamnosus derived treatment [43]. Probiotic therapy has also been used to improve both infectious disease and behavioral issues, indicating the potential breadth and therapeutic potential of probiotics.
On the other hand, the potential claims of probiotics in treating disease have been called into question by a number of reviews, citing the lack of evidence or controversial results [28]. Two clinical reviews found insufficient evidence supporting probiotic use for treating IBS, Crohn’s disease, and necrotizing enterocolitis and sepsis in preterm infants [26,44]. Furthermore, studies have shown adverse effects when using probiotics in humans or effects in animal models that do not translate well in humanized trials [45,46]. There is also a lack of scientific evidence supporting the widely-advertised functional foods containing probiotics [47]. Companies with products like yogurt widely advertise to the general public using attractive claims such as improving digestion or strengthening the body’s natural defenses but cite questionable studies [47]. In a systematic review analyzing significance of studies containing L. casei and B. lactis , several of the studies were found linked to companies selling products containing those species through funding, company-supplied products, or a company-associated author [47].
Future of Probiotic Design
Given the new technological advances, cost effectiveness, and scale of the data generated by next-generation sequencing, we can analyze the relationship between the microbiome and probiotic-derived therapies. It is likely that given the proper knowledge of each individual’s microbiome, diet, and lifestyle, accurate prescription of probiotics can be designed. In a human study of obese individuals, Korpela et al. [48] demonstrated that the microbiome, rather than weight status, can help determine the efficacy of treatments. Using fecal microbiota analysis at baseline and after dietary interventions, they were able to predict responsiveness of the host microbiota using microbial biomarkers [48]. Another human study profiling geriatric individuals revealed potential microbial biomarkers between probiotic respondents and non-respondents [49]. It was previously suggested that level of response depended on change in cholesterol; instead, the fecal microbiota provided a more clinically useful biomarker [49]. It is also important to note that the geriatric study identified a microbiota shift in non-respondents [49]; thus, it is pertinent to distinguish individuals that respond or do not respond to dietary interventions such as probiotics.
Due to the diverse results of probiotic studies, gut microbiota profiling could help tailor probiotic therapies per individual. With conditions highly dependent on glucose or insulin metabolism, personalized nutritional treatments have the greatest prospects. For example, probiotic therapy may be especially useful in cases where the identification of a “pre-diseased” microbiome is compared to a profile during disease or flare and specific probiotic bacterial species once lost are restored. This targeted microbiota therapy (TMT) would then be restoring species that have already been able to previously colonize and survive in an individual’s particular intestinal landscape (Figure 1). However, this therapy relies on understanding the various healthy prediseased and diseased microbiome and, while the advent of stool banks and sequencing of the microbiome makes this information accessible to the general population, this knowledge will still be a limiting factor in such designs. A second and more intriguing notion is the idea of probiotic sensitization in which the intestinal microenvironment is somehow manipulated through drugs, prebiotics, or antibiotics in an effort to alter the topography of the intestine, making it more permissive for colonization and establishment of the specific probiotic.
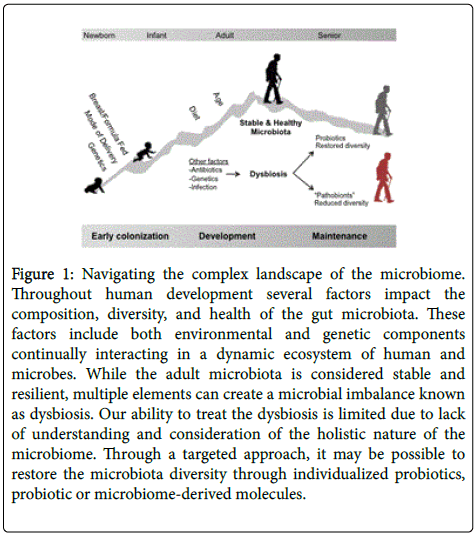
Figure 1: Navigating the complex landscape of the microbiome. Throughout human development several factors impact the composition, diversity, and health of the gut microbiota. These factors include both environmental and genetic components continually interacting in a dynamic ecosystem of human and microbes. While the adult microbiota is considered stable and resilient, multiple elements can create a microbial imbalance known as dysbiosis. Our ability to treat the dysbiosis is limited due to lack of understanding and consideration of the holistic nature of the microbiome. Through a targeted approach, it may be possible to restore the microbiota diversity through individualized probiotics, probiotic or microbiome-derived molecules.
The anatomical, immunological, and microbiological complexity of the GI tract has likely contributed to the variable results seen in clinical trials and human studies regarding probiotic therapy. Utilizing the technological and intellectual advances regarding the microbiome, we need to have a plan for the design and execution of a new generation of probiotics using TMT. We must spend more time and effort understanding traits that underlie the probiotic function. We need to identify the genes responsible for this given function through mutagenesis studies that will help to derive mutants, isolate genes, and/or identify proteins responsible for the probiotic function. Once identified, we will need to either design genetically modified microorganisms (GMM) or synthesize and test the probiotic-derived protein to establish a treatment not requiring the delivery of a live bacteria. Lastly, we must establish more standardized treatments and standardized strains in order to compare efficacy across clinical trials. Only when we start rigorous, cross-disciplinary, and holistic studies that consider the analysis of the microbiome, dietary, and lifestyle choices both pre- and post-probiotic treatment will we be able to create significant therapies utilizing probiotics or microbiome-derived products.
Humans are known for their diversity and complexity as a species and the identification of the microbiome, with its associated 9 million genes, have further increased our diversity and individuality. Here we discussed the numerous ways the uniquely tailored microbiome is formed, shaped, and altered throughout the human lifespan by genetic and environmental factors. As our knowledge of the microbiome continues to expand, the need to include microbiota analysis in conjunction with disease and treatment studies is crucial as the microbiome could confound our understanding of host and disease. Our current lack of understanding, especially in regards to probiotics, could be the cause of the contradicting results at the research and clinical trial level with probiotic treatments. Before proposing probiotic and nutritional interventions that induce microbiome changes, we first need to acknowledge and incorporate our findings of the microbiota landscape. Only through the comprehensive approach of integrating our microbial ecosystem with nutritional and medical knowledge will we be able to advance the field of probiotics.