Journal of Fertilization: In Vitro - IVF-Worldwide, Reproductive Medicine, Genetics & Stem Cell Biol
Open Access
ISSN: 2375-4508
ISSN: 2375-4508
Research Article - (2016) Volume 4, Issue 1
Objectives: The current study aimed to investigate the in vitro effects of physiologic concentration of adiponectin in follicular fluid of pre-ovulatory bovine ovarian follicles on: 1) the bovine oocyte maturation and the early developmental rates of bovine embryos in vitro; 2) the expression profiles of some developmentally important marker genes in the bovine cumulus cells and oocytes.
Materials and Methods: The cumulus-oocyte complexes (COCs) were aspirated from abattoir ovaries and cultured in vitro in the presence/absence of the recombinant human (rh) adiponectin (5 μg/mL) and used for real time RT-QPCR and in vitro embryo production assays.
Results: Adiponection had no significant effects on nuclear maturation and cumulus expansion rates (p > 0.05). The expression levels of hyaluronic synthase 2 gene (Has2) was significantly higher in the adiponection group compared to the control group (p = 0.014). However, the early embryonic development was not affected significantly in the presence of adiponectin. Adiponectin did not change the expression rates of adiponectin receptors or other markers of oocyte competence. Also, the expression rates of two important oocyte secreted factors (OSFs), bone morphogenetic protein 15 (Bmp15) and growth/differentiation factor 9 (Gdf9) were not affected (p > 0.05).
Conclusion: Adiponection did not affect the bovine oocyte developmental competence and early embryo development in spite of a significant increase in the expression level of Has2 gene, a biomarker of the future oocyte developmental competence.
<Keywords: Adiponectin, Bovine, Gene expression, Oocyte competence, Embryo development
It has been well accepted that reproductive performance is strongly associated with nutrition in different farm animals [1]. Both chronic over- or under-nutrition has been demonstrated to result in reproductive failure [1]. Researchers have shown that several hormones, metabolites and neuropeptides are involved in regulation of reproductive functions. Adipose tissues synthesize and secret several factors (adipokines) which act at various levels to control male and female reproductive functions [2]. Adiponectin, as one of the adipokines, is the most abundant protein secreted from adipose tissues [3,4]. Metabolic and anti-inflammatory properties of adiponectin have been previously confirmed [2]. Furthermore, recent evidence suggests that adiponectin has a direct peripheral role in regulation of reproductive functions [2]. Two receptors (Adipor1 and Adipor2) have been identified for adiponectin in different species including humans, rodents, pigs, chicken, and cows [5]. Previous studies have clearly shown that adiponectin and its receptors are expressed by follicular cells (granulosa and theca cells), the oocyte and the corpus luteum in several species. Moreover, the adiponectin accumulates in the follicular fluid from a local source and/or from circulation [5].
The stage of follicular and luteal development influenced the expression pattern of adiponectin and its receptors in the bovine ovaries [6]. Adiponectin receptors were expressed significantly higher in granulosa and theca cells of the bovine large follicles. Furthermore, the adiponectin expression increased in the bovine granulosa cells and in cumulus-oocyte complex during follicular development from small to large follicles [6]. However, adiponectin is expressed at undetectable or very low levels in mouse or human granulosa cells [7,8]. Tabandeh et al. [9] showed that the theca/ cumulus cells or high quality oocytes recovered from dominant follicles expressed higher amounts of adiponectin and its receptors compared to atretic follicles or lower quality oocytes during both the follicular and luteal phases in the bovine. However, results from studies in other species did not follow the same patterns. For example, in chickens and pigs, there were no variations in the levels of adiponectin receptors in granulosa cells compared to theca cells or among different sizes of follicles [10-12]. It has also been shown that adiponectin differentially alters the expression of genes involved in steroidogenesis, ovulation and apoptosis in cumulus cells and enhances oocyte maturation and early embryo development in the mouse, human and the swine [6,13]. No significant effect were observed on the bovine in vitro oocyte maturation, cleavage and blastocyst formation rates in the presence of adiponectin [14]. However, adiponectin decreased insulin-induced steroidogenesis and increased IGF-1-induced proliferation of cultured granulosa cells through a potential involvement of ERK1/2 MAPK pathway [14]. Also, previous studies have confirmed that the changes in the expression levels of some genes in cumulus cells are highly correlated with the future developmental competence of the enclosing oocytes so that they are considered as markers of the oocyte quality [15,16]. Therefore, the current research was designed to investigate the in vitro effects of adiponectin at physiologic concentrations found in the follicular fluid of the pre-ovulatory bovine ovarian follicle on 1) the bovine in vitro oocyte nuclear maturation and cumulus expansion rates, 2) the early development of bovine embryos in vitro, 3) the expression profiles of adiponectin receptors and some developmentally important marker genes in the cumulus cells and 4) the expression rates of the two most important oocyte secreted factors: Bmp15 and Gdf9.
Determination of adiponectin concentration in the follicular fluid of bovine pre-ovulatory follicles
The concentration of the follicular fluid adiponectin was determined in the pre-ovulatory follicle in dairy cows as previously described by Kafi et al. [17]. Briefly, the estrous cycle of lactating cows (n = 8) was synchronized using two injections of prostaglandin F2α 13 days apart. Six to 12 hours after detection of the estrus, ovarian ultrasonography was performed to assess the presence of a pre-ovulatory follicle (10–17 mm diameter). Then, a sample of follicular fluid was aspirated transrectally using a long fine-needle covered by a hard plastic tube under the caudal epidural anesthesia (2% Lidocaine hydrochloride; 0.2 mg/kg). All samples of follicular fluid used for hormone assay were free of blood contamination. Then, adiponectin concentrations were determined in the follicular fluid of the pre- ovulatory follicles. Validated commercial ELISA kit (Bovine Adiponectin, CUSABIO Biotech Co. Ltd., China) was used for adiponectin measurement. The analytical sensitivity of the test was typically less than 1.562 mg/mL. The interassay coefficient of variability for low and high bovine adiponectin (3.125 and 50 mg/ mL, respectively) were 6.2% and 5.9%, respectively. The intra-assay variations for the two above samples were 3.4% and 4.7%, respectively.
Collection of cumulus-oocyte complexes and in vitro maturation
Bovine ovaries (Bos taurus) collected at a local abattoir were transported to the laboratory within 2-3 hours in sterile normal saline maintained at 35-37ºC. Upon arrival at the laboratory, ovaries were washed five times in pre-warmed (37ºC) sterile phosphate buffered saline. Cumulus-oocyte complexes (COCs) were recovered from healthy antral follicles (3-8 mm in diameter with smooth, transparent outer walls) using the aspiration method with a 10 mL hypodermic syringe fitted to a 20G needle. The aspirated COCs were evaluated for the morphological appearance of the cumulus cell investments and homogeneity of ooplasm under a stereo-microscope and only COCs with at least 3 complete layers of cumulus cells and finely granulated homogeneous ooplasm were selected as suitable for in vitro maturation [18]. The selected COCs were rapidly washed five times in drops of equilibrated oocyte washing medium consisting of HEPES-buffered TCM-199 (Sigma, M4530-500ML, St. Louis, MO, USA) supplemented with 10% FBS. The maturation medium consisted of TCM-199 supplemented with 0.1 IU/mL recombinant human FSH (Follitrope, LG Life Sciences, South Korea), 5 IU/mL highly purified hCG (Karma, Pharmatech GmbH, Germany) and 3 mg/mL Bovine Serum Albumin (BSA, Sigma, A3675-5G, St. Louis, MO, USA) in the control group and all of the control medium components plus recombinant human (rh) adiponectin at final concentration of 5 μg/mL (BioVendor Research and Diagnostic Products, RD172023010, Germany) in the treatment group. Groups of 30-50 COCs were incubated in 500 μL equilibrated maturation medium in four-well culture dishes (NuncTM) for 24 h at 38.5ºC in 5% CO2 under maximum relative humidity. Totally, 554 excellent and good COCs were randomly assigned to treatment (220 COCs) and control (334 COCs) groups in five independent repeats.
In vitro fertilization and embryo culture
After 24 hours of in vitro maturation with or without adiponectin (5 μg/ml), COCs were washed five times in oocyte washing medium (HEPES-buffered modified Tyrode’s medium supplemented with 0.168 mg/mL Na bicarbonate, 10 mM Na lactate, 0.009 mg/ml Na pyruvate, 6 mg/mL fatty acid free BSA and 50 μg/mL gentamicin). Next, they were transferred into 500 μL of fertilization medium (modified Tyrode’s medium supplemented with 0.27 mg/mL caffeine, 10 μg/mL Heparin, 50 μg/mL Hypotaurine, 6 mg/mL BSA, 1 mg/mL Na pyruvate, 10 mM Na lactate and 2.2 mg/mL Na bicarbonate) in four-well dishes (NuncTM) (50 COCs/well). In order to obtain motile spermatozoa, frozen/thawed semen from bulls with proved fertility incubated for 45 minute in sperm capacitation medium (modified Calcium-free Tyrode’s medium supplemented with 0.1 mg/mL caffeine, 6 mg/mL BSA, 1 mg/mL Na pyruvate, 10 mM Na lactate and 2 mg/mL Na bicarbonate) and then centrifuged for 10 minute at 200 × g. Then, the supernatant was discarded and the sperm pellet was re-suspended in fertilization medium (approximately 6 million spermatozoa/mL). COCs and spermatozoa were incubated together in 500 μL of the fertilization medium/well for 18 hours at 38.5ºC in a humidified atmosphere with 5% CO2 in 95% air. Semen from the same bull was used throughout the experiment. Day of fertilization was considered as Day 0. At the end of the fertilization period, presumptive zygotes were completely denuded by repeated pipetting and washed in serum-free modified synthetic oviduct fluid (mSOF) supplemented with 4 mg/mL fatty acid-free BSA and cultured in same medium in four-well dishes under paraffin oil at 38.5ºC for 8 days in a water-saturated atmosphere of 6% CO2. The embryo culture medium was renewed every 48 hours. Embryonic cleavage, morula and blastocyst rates were determined on Days 2, 4 and 8 of in vitro culture, respectively. Totally, 397 excellent and good quality COCs were randomly assigned to treatment (199 COCs) and control (198 COCs) groups in five independent repeats. All in vitro fertilization reagents were from Sigma-Aldrich™, Germany.
Assessment of oocyte nuclear maturation and the cumulus cell expansion
After 24 hours of in vitro maturation, the degree of cumulus expansion was scored under a stereo- microscope based on the subjective scale of 0 to 3 describe by Lorenzo et al. [19], in which 0 indicates no detectable response; 1 shows the minimum observable response; 2 indicates partial expansion, with more than half of the cumulus expanded; and 3 shows full expansion, where all layers of cumulus cells have expanded, including those closest to the oocyte. Then, the oocytes were mechanically denuded of the surrounding cumulus cells using repeated pipetting in 3% sodium citrate solution and were mounted on grease-free glass slides with coverslips hold in place with a mix of vaseline and paraffin wax (20:1). The slides were immersed in the oocyte fixation solution (ethanol/acetic acid, 3:1 v/v) for 24 hours. The oocytes were stained with 1% aceto-orcein (1% orcein in 45% glacial acetic acid) and examined for nuclear morphology with compound microscope at x100 and x400 magnifications. Based on their nuclear status, oocytes were classified as immature (germinal vesicle, germinal vesicle break down), in resumption of meiosis (metaphase I, anaphase/telophase I), mature (metaphase II) or degenerating (pyknotic chromatin at various meiotic stages or degenerated chromatin spread throughout oocyte) [20].
Collection of samples for expression analyses
Immature COCs: The collected bovine ovaries intended to be used for gene expression analysis of the immature COCs were transported to the laboratory in sterile 4ºC normal saline and washed three times with cold sterile PBS (5-10ºC) upon the arrival to the laboratory. The aspirated immature COCs meeting the selection criteria were washed three times in drops of cold molecular grade PBS (EUROIMMUN®) and transferred to the sterile RNase/DNase free 1.5 mL microtubes in groups of 50-70, repeated five times, independently. The microtubes were then centrifuged twice for 6 minute at 600 g at 4ºC, the supernatant was discarded and 350 μL of RNA extraction lysis buffer (RNeasy Plus Mini® Kit, Qiagen, Germany) was added to the cell pellets and mixed by vortexing for 1 minute. Samples were then snap-frozen in liquid nitrogen and stored at -80ºC until further processing.
In vitro maturation : After 22-24 hours of maturation, pools of 30-50 COCs (five repeats for control group and five repeats for treatment group) were selected and mechanically denuded by repeated pipetting in 3% sodium citrate. The completely denuded oocytes were washed five times in drops of cold molecular grade PBS and transferred to the sterile RNase/DNase free 1.5 mL microtubes. The cumulus cells were also collected and transferred to separate microtubes. Both oocyte and cumulus samples were then centrifuged twice for 6 minute at 600 × g at 4ºC, the supernatant was discarded and 350 μL of RNA extraction lysis buffer (RNeasy Plus Mini® Kit, Qiagen, Germany) was added to the cell pellets and mixed by vortexing for 1 minute. Samples were then snap-frozen in liquid nitrogen and stored at -80ºC until further processing.
RNA isolation and reverse transcription
Total RNA was extracted from the oocytes and cumulus cells using the RNeasy Plus Mini® Kit as described by the manufacturer (Qiagen, Germany). After purification, RNA samples were eluted in 60 μL of RNase-free water and stored at −80ºC till further use. Total RNA concentration was measured by spectrophotometry method (absorbance value at 260 nm) and purity was assessed using the A260/A280 nm ratio with expected values between 1.8 and 2.0. The cDNA synthesis was carried out with 6 μL of the total RNA (350-400 ng) using PrimeScript™ RT reagent Kit (Perfect Real Time, TAKARA, Japan), and both Oligo-dT and random hexamers as primers according to the manufacturer instructions. In order to test the effectiveness of specially designed gDNA Eliminator spin columns (provided by the RNA extraction kit manufacturer) in removal of the genomic DNA contamination, transcriptase minus samples were run together with test samples.
Selection of the cumulus and oocyte genes
Levels of the cumulus expression of all selected genes have been previously confirmed to be highly associated with the future developmental competence of the oocyte. Genes encoding for the two most important oocyte secreted factors were also selected. A summary of the studied genes, their full names and the principle functions are presented in Table 1.
| Gene | Target Cell | Primer Sequences (5‘ to 3’) | Primer Source | Fragment size (bp) / GenBank Accession No. | Function |
|---|---|---|---|---|---|
| hyaluronan synthase 2 | C | F:ATAAATGTGGCAGGCGGAAGAAGGR:GTCTTTGTTCAAGTCCCAGCAGCA | [36] | 184/NM_174079.2 | ECM formation and stabilization |
| tumor necrosis factor, a-induced protein 6 | C | F:GTCTGGCAAATACAAGCTCACCTR:GGATCTGTAAACACACCACCACA | [36] | 297/NM_001007813 | ECM formation and stabilization |
| BCL2-associated X protein | C | F:TGCTTCAGGGTTTCATCCAG R:GTGTCCCAAAGTAGGAGAGG |
[22] | 425/NM_173894 | Pro-apoptotic |
| B-cell /lymphoma 2 | C | F:GAGATGTCCAGTCAGCTGCACC R:ATAGGCACCCAGGGTGATGC | [22] | 366/U92434 | Anti-apoptotic |
| Follicle Stimulating Hormone Receptor | C | F:AATCTACCTGCTGCTCATAGCCTC R:TTTGCCAGTCGATGGCATAG |
[23] | 76/NM_174061 | Cumulus expansion and nuclear maturation |
| Adiponectin Receptor 1 | C | F:CTCATCTACCTCTCCATCGTCTGR:CCTGCTCTTGTCTGCCGGTGCTTG | [6] | 101/NM-001034055 | Adiponectin Function |
| Adiponectin Receptor 2 | C | F:AGGTCGGGAGCGTCTTGTAGR:CATGAACCCCTCATCTTCCTGAG | [6] | 109/NM-001040499 | Adiponectin Function |
| glyceraldehyde-3-phosphate dehydrogenase | C/O | F:ACCCAGAAGACTGTGGATGG R:CAACAGACACGTTGGGAGTG |
[36] | 178/Bt.87389 | Basal cell metabolism |
| bone morphogenetic protein 15 | O | F:ATCATGCCATCATCCAGAACC R:TAAGGGACACAGGAAGGCTGA |
[23] | 72/NM_001031752.1 | OSF |
| growth and differentiation factor 9 | O | F:AGCGCCCTCACTGCTTCTATAT R:TTCCTTTTAGGGTGGAGGGAA |
[23] | 80/NM_174681 | OSF |
F = Forward; R = Reverse; OSF = Oocyte Secreted Factor; C = Cumulus; O = Oocyte
Table 1: Primer sequences, target cells, expected fragment size of the amplified products, GenBank accession numbers and principle functions of the housekeeping and study genes.
Primer selection and optimization
All primers were selected from previous studies and re-evaluated by AlleleID® software (Version 7.7, Premier Biosoft International) for meeting the real-time RT-PCR primer selection criteria and also checked in GenBank for specificity. A primer optimization study was performed using the real-time RT-PCR SYBER Green assay for each primer set to find the best primer pair concentrations in terms of reaction sensitivity and specificity at a constant annealing temperature of 60ºC. The primer characteristics, i.e., oligonucleotide sequences, target cells, expected fragment size of the amplified products and GenBank accession numbers are shown in Table 1.
Quantitative real-time polymerase chain reaction
SYBR Green quantitative real-time RT-PCR assay was performed to quantify the mRNA transcripts of all examined target and housekeeping genes. The PCR mix included 10 μL of SYBR® Premix Ex Taq II (Tli RNaseH Plus, TAKARA, Japan), reverse and forward gene specific primers at final optimized concentrations for each gene, 2 μL of 1:5 diluted cDNA (20 ng/μL) and enough DEPC-treated water to the final reaction volume of 20 μL. The PCR was performed using MiniOpticon™ Real-Time PCR Detection System (Bio-Rad, USA) and the thermal cycling program was (identical for all genes): initial sample incubation at 95ºC for 30 seconds, followed by 40 cycles at 95ºC for 5 seconds and at 60ºC for 45 seconds, and finally, heating from 55ºC to 95ºC with increments of 0.2ºC and holding for 2 seconds to generate the melting curve in order to confirm the specificity of the PCR product. All samples were run in triplicate and the mean value of each triplicate was used for all further calculations. Reverse transcriptase minus and blank (water replacing cDNA) controls were run together with test samples. Standard curves were generated for each gene by serial dilution of the cDNA samples and the slope of the resultant curve was used to calculate the individual real-time PCR efficiencies (E = 10 (-1/slope)). The relative expression ratios were calculated by a mathematical model, which included a correction for real-time PCR efficiency of the individual transcripts [21], as follows: Ratio = (E target) ΔCt target (control-sample) / (E ref) ΔCt ref (control-sample). The relative expression ratio of a target gene was computed based on its real-time PCR efficiencies (E) and the threshold cycle difference for an unknown sample versus a control (Ct control–sample).
Statistical analyses
Data were tested for normality by Shapiro-Wilk normalization test. The oocyte nuclear maturation, cumulus expansion and embryo development data were compared using the Chi-squared test and p < 0.05 was considered statistically significant. The difference between the relative expression of the investigated transcripts was tested for significance by two independent samples t-test (p < 0.05 was considered significant and p ≤ 0.09 was considered as a tendency towards a significant level). All statistical analyses were performed using a computer-aided statistical software package (IBM® SPSS Statistics version 22 for windows) [22]. The experiment was carried out in five independent replicates.
Adiponectin concentration in the follicular fluid of the preovulatory follicle
The mean (± SD) for the volume of follicular fluid collected was 1.3 ± 0.3 mL. Adiponectin concentrations in the follicular fluid of the preovulatory follicles were 4.05 ± 0.31 μg/mL (mean ± SD). Therefore, in the current study, the oocyte culture media were supplemented with rh adiponectin at the final physiologic concentration of 5 μg/mL.
Effect of adiponectin on cumulus cell expansion, oocyte nuclear maturation rates and early embryo development
The normal distribution of cumulus expansion and nuclear maturation data was confirmed by Shapiro-Wilk normality test (p > 0.05). Results of the two independent samples t-test showed that the percentages (mean ± SD) of MII oocytes and full expansions in the control group (80.79 ± 10 and 71.15 ± 14%, respectively) was not statistically different from adiponection group (76.9 ± 10 and 71.27 ± 9.7%, respectively) as summarized in Figure 1 and Table 2. Bovine early embryonic development was not significantly affected (p > 0.05) with the presence of adiponectin in the oocyte maturation medium, as summarized in Table 3.
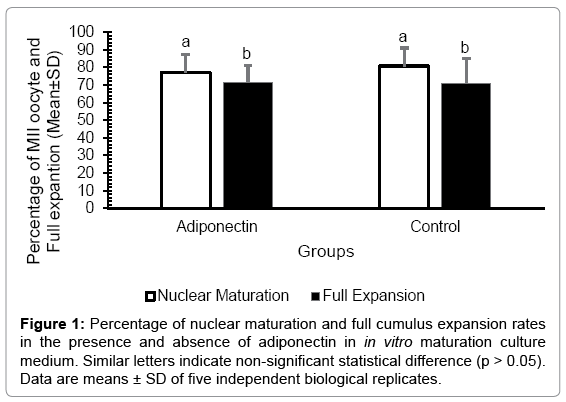
Figure 1: Percentage of nuclear maturation and full cumulus expansion rates in the presence and absence of adiponectin in in vitro maturation culture medium. Similar letters indicate non-significant statistical difference (p > 0.05). Data are means ± SD of five independent biological replicates.
| Groups | Total no. of cultured oocytes | Mature (%) | Immature (%) | In resumption of Meiosis (%) |
Degenerated (%) |
|---|---|---|---|---|---|
| Control | 334 | 80.79 ± 10.2 | 15.5 ± 7.5 | 0.3 ± 0.95 | 3.4 ± 3.2 |
| Treatment | 220 | 76.95 ± 10.6 | 16 ± 2.7 | 6.6 ± 8.5 | 0.46 ± 0.92 |
Data are Means ± SD
Table 2: Summary of nuclear maturation stages of the bovine oocytes after in vitro maturation in the presence / absence of adiponectin.
| Groups | Oocytes (n) | Cleaved on D2, n (%) | Morula on D6, n (%) | Blastocyst on D8, (%) | Morula and blastocyst on D8, n (%) | Blastocyst on D8/cleaved (%) |
|---|---|---|---|---|---|---|
| Control | 199 | 124 (62.3 ± 10.7) | 82 (41.2 ± 10.3) | 32 (15.7 ± 6.0) | 43 (21.3 ± 7.0) | 26.0 ± 11.0 |
| Treatment | 198 | 129 (64.7 ± 6.8) | 81 (40.7 ± 4.5) | 31 (15.4 ± 5.0) | 42 (20.9 ± 7.1) | 24.5 ± 10.3 |
Data (Means ± SD) are generated from five independent replicates
Table 3: Effect of adiponectin in in vitro maturation medium on bovine early embryonic development.
Effect of adiponectin on the cumulus cells gene expression
The normal distribution of gene expression data was confirmed by Shapiro-Wilk normality test (p > 0.05). Results of the relative gene expression study in the cumulus cells are presented in Figure 2.
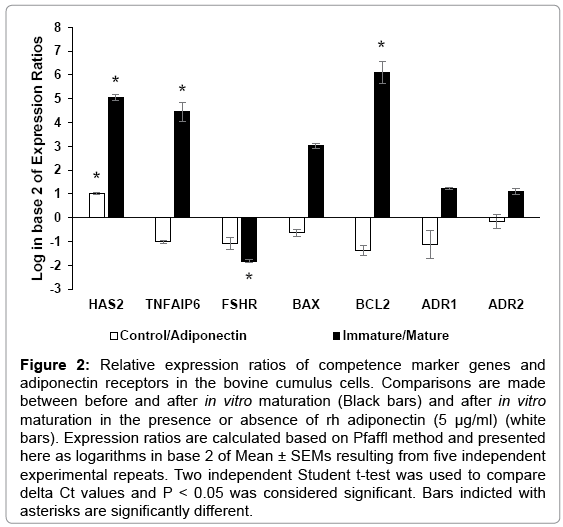
Figure 2: Relative expression ratios of competence marker genes and adiponectin receptors in the bovine cumulus cells. Comparisons are made between before and after in vitro maturation (Black bars) and after in vitro maturation in the presence or absence of rh adiponectin (5 μg/ml) (white bars). Expression ratios are calculated based on Pfaffl method and presented here as logarithms in base 2 of Mean ± SEMs resulting from five independent experimental repeats. Two independent Student t-test was used to compare delta Ct values and P < 0.05 was considered significant. Bars indicted with asterisks are significantly different.
Has2 gene was up-regulated in the adiponection group compared to the control group by a mean factor of 2.015 ± 0.05 times, significantly (p = 0.01). The relative abundance of the Has2 transcripts was significantly higher (p = 0.04) in the in vitro matured COCs compared to immature COCs by a mean factor of 33.56 ± 3 times.
Tnfaip6 gene expression is not significantly different between adiponectin and control groups. However, Tnfaip6 is differentially expressed in the in vitro matured COCs compared to immature COCs by a mean factor of 21.88 ± 5.8 times, significantly (p = 0.01). Fshr gene is not significantly expressed in the adiponection group compared to the control group. However, Fshr gene is significantly down-regulated (p = 0.009) in the in vitro matured COCs compared to immature COCs by a mean factor of 0.28 ± 0.01 times.
Bax, Bcl2andBcl2/Bax ratio
Bax and Bcl2gene expression was not different significantly between adiponectin and control groups, although there was a tendency for the Bax gene towards significant downregulation (p = 0.08) by a mean factor of 0.64 times. The relative abundance of the Bax gene transcripts was not significantly different in the in vitro matured COCs compared to immature COCs. However, the Bcl2gene was significantly up-regulated (p = 0.001) in the in vitro matured COCs compared to immature COCs by a mean factor of 69.34 ± 21 times. The Bcl2/Bax ratio was not significantly different between adiponectin and control groups. The expression of Adipor1 and Adipor2 was not statistically different either between adiponectin and control groups or between in vitro matured COCs and immature COCs.
Effect of adiponectin on the expression of marker genes in the oocyte
Results of the relative gene expression study in the oocyte are presented in Figure 3. There was a tendency towards a significance differences (p = 0.06) in the relative abundance of the Bmp15 transcripts between adiponectin and control groups where Bmp15 gene tending to be up-regulated in the adiponectin group by a mean factor of 1.46 times. However, there was no significant difference between immature and in vitro matured COCs. The expression of Gdf9 gene was not statistically different between adiponectin and control groups. However, the Gdf9 gene was significantly down-regulated (p = 0.009) in the in vitro matured COCs compared to immature COCs by a mean factor of 0.01 ± 0.002 times.
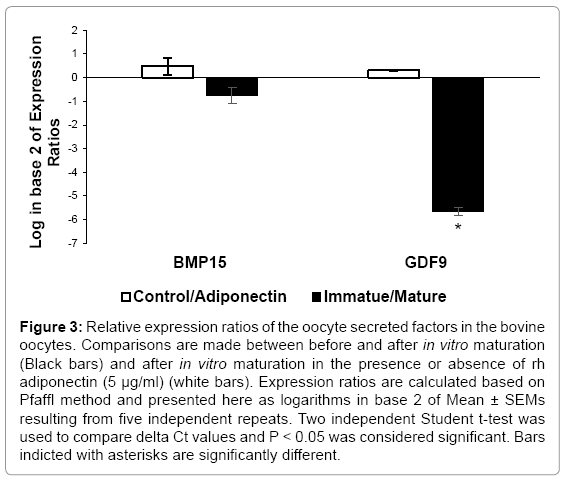
Figure 3: Relative expression ratios of the oocyte secreted factors in the bovine oocytes. Comparisons are made between before and after in vitro maturation (Black bars) and after in vitro maturation in the presence or absence of rh adiponectin (5 μg/ml) (white bars). Expression ratios are calculated based on Pfaffl method and presented here as logarithms in base 2 of Mean ± SEMs resulting from five independent repeats. Two independent Student t-test was used to compare delta Ct values and P < 0.05 was considered significant. Bars indicted with asterisks are significantly different.
The nuclear maturation and cumulus expansion are two critical steps in oocyte competence acquisition, normal oocyte development, ovulation, and fertilization [17,21,23,24]. Oocytes with no cumulus expansion have limited potential for implantation [25]. Our findings demonstrated that rh adiponectin at physiological concentrations had no significant effects on bovine oocyte nuclear maturation and degree of cumulus cell expansion. Similarly, Maillard et al. [14] reported that supplementation of in vitro maturation medium with rh adiponectin had no significant effects either on cumulus expansion or nuclear maturation rates of the bovine oocytes derived from small follicles, or the 48 hours cleavage and day 8 embryo development rates. Also, rh adiponectin alone did not stimulated mouse COCs expansion; however, it enhanced significantly the in vitro fertilization and development of the mouse embryos to the blastocyst stage [7]. In the pig, however, Chappaz et al. [13] found that recombinant porcine adiponectin significantly decreased the frequency of meiotic immature oocytes derived from large follicles (3–6 mm) but not from small follicles (< 3 mm), suggesting an acceleration of the meiotic maturation of porcine oocyte by adiponectin. This discrepancy between different studies could be linked to the animal species or to the size of the follicles used for COCs recovery and also the dose of the adiponectin they used. Our results further showed that addition of rh adiponectin during in vitro maturation does not influence on the subsequent early embryonic development. We purposefully used BSA instead of FBS in the culture media to be able to control the rh adiponectin level in the culture dishes. We used, in contrast to most previous studies, the physiologic concentrations of the rh adiponectin detectable in the follicular fluid of the large pre-ovulatory ovarian follicles for performing the in vitro study.
The bidirectional communication between the oocyte and its cumulus cells is very important to oocyte competence acquisition [26]. In addition, the properties and functions of cumulus cells are influenced by the oocyte secreted factors [27]. Therefore, the gene expression profiles in cumulus cells are important indicators for the quality of the oocyte [15]. Formation of the extracellular matrix during cumulus cells expansion is dependent to the expression of genes including Has2 and Tnfaip6. The expression level of these two genes has been proved to be correlated with the future developmental potential of the oocyte [28]. In the present study, we showed that the expression level of Has2 gene in the cumulus cells significantly increased by a mean factor of 2.015 ± 0.05 times in the presence of adiponection at physiological concentrations. However, the relative abundance of the Tnfaip6 transcripts was not affected significantly. Similar to our findings, Richards et al. [7] reported that rh adiponectin (20 μg/mL) alone stimulated significant increases in mRNA encoding Has2 in mouse COCs. However, this increase in the expression of Has2 was not adequate to lead to an observable positive response in the degree of cumulus cell expansions or early embryonic development in the present study and Richards et al. [7]. The increased level of Has2 expression may has been masked by the potent stimulatory effects of FSH. Richards et al. [7] further showed that, unlike FSH, adiponectin alone did not increase Tnfaip6 transcripts but did enhance FSH mediated increases in Has2 gene expression. It has been established that the FSH plays a positive stimulatory role in cumulus expansion and nuclear maturation rates in several species during in vitro oocyte maturation. A reduction in the expression level of FSH receptor could reduce the efficiency of FSH [29]. Our results showed that the rh adiponectin at THEconcentration of 5μg/ml does not affect the FSHR gene expression significantly. Bax and Bcl2, which are, respectively, pro- and anti-apoptotic proteins of the Bcl2 gene family, participate in the mitochondria-dependent apoptosis pathway [30]. Bcl2 expression in cumulus cells is strongly associated with the ability of oocytes to complete nuclear maturation [31]. The ultimate vulnerability of cells to apoptotic stimuli is determined by the relative ratio of pro- and anti-apoptotic members of the Bcl2 family (Bcl2/Bax ratio) [32-34]. In the current research, rh adiponectin at physiologic concentrations did not affect expression rates of Bax, Bcl2 genes orBcl2/Bax ratio in cumulus cells, although, there was a tendency towards significant down-regulation of Bax gene in the presence of rh adiponectin. However, theBcl2 gene was highly up-regulated in the in vitro matured COCs compared to immature COCs by a mean factor of 69.34±21 times. Similar to our findings, Richards et al. [7] showed that the rh adiponectin (20 μg/ml) in the presence of FSH, did not alter the expression of Bmp2 gene, an apoptotic marker, in the mouse COCs. However, according to Pierre et al. [35], rh adiponectin and its receptors may alter apoptosis in human granulosa tumor cell line (KGN) cells. They showed a strong reduction in Adipor1 expression in KGN cells, a significant increase in the level of cleaved caspase-3 and a significant decrease in the level of Bcl-2-associated death promoter phosphorylation which suggest an increase in the apoptosis. The involvement of adiponectin system in regulation of follicular growth and development was shown recently [6]. They showed that the expression levels of Adipor1 and Adipor2 were higher in bovine cumulus cells and oocyte derived from large growing follicle compared with that in cumulus cells and oocyte derived from small and medium follicles [6]. The association of adiponectin and its receptors genes with follicular dominance and oocyte competence was also confirmed [9]. However, changes in level of expression of adiponectin receptors during in vitro maturation and the effect of adiponectin on their expression have not been examined before. Present study showed that the expression of Adipor1 and Adipor2 in cumulus cells was not statistically different either between adiponectin and control groups or between in vitro matured COCs and immature COCs. The abundance of Has2, Tnfaip6, Fshr andBcl2 transcripts in the cumulus cells from mature COCs showed a significant difference in comparison with immature ones which confirms their involvement in the final maturational processes as reported in previous studies [36,37].
Oocyte secreted factors are crucial regulators of cumulus cells differentiation and function [15]. Bmp15 and Gdf9 are two most important OSFs which their expression in the oocyte are highly correlated with the future developmental potential of COCs [26]. We showed that rh adiponectin at physiologic concentrations did not influence the expression rates of Bmp15 and Gdf9 in the bovine oocyte. However, the difference in Bmp15 transcripts abundance showed a tendency towards a significant increase in the oocytes matured in the adiponectin group. The expression level of Gdf9 was reduced significantly after 24 hours oocyte culture than that of the commencement of the culture at hour 0, while the expression of Bmp15 was not different between in vitro matured and immature oocytes. This finding is similar to the previous report in bovine species [38].
Our findings showed that rh adiponection at physiologic concentrations did not affect the oocyte developmental competence and subsequent early embryo development in spite of an increase in the expression level of Has2 gene, a biomarker of future oocyte developmental competence.
The research was funded by the Center for Biotechnology of Animal Reproduction and Shiraz University. The authors are thankful to Dr. M Masoodian and M Ghahremani-Seno for their technical assistance in the Central Laboratory of the School of Veterinary Medicine, Shiraz University. We warmly appreciate H Jesmani and Z Taghipour for their cooperation in performing the research. The authors declare no conflict of interest on the publication of this paper. This research was approved by the local Ethics Committee of Shiraz University and complies with Shiraz University animal welfare guidelines and policies.