Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Short Communication - (2015) Volume 4, Issue 6
MUC1 mucin is a type I transmembrane glycoprotein expressed at the apical border of healthy epithelia that line the respiratory, reproductive and gastrointestinal tracts and is overexpressed in various cancer cells. The goal of the study was to check the effect of berenil complex of Platinum(II) applied together with anti-MUC1 antibody on induction of programmed cell death in human skin fibroblasts. The influence of novel platinum(II) complex used with anti-MUC1 on the concentration of selected markers of apoptosis such as p53, Bax, cytochrome c, caspase-8, -9 and caspase-3 was determined using the ELISA technique. The results from combined treatment were compared with those obtained using monotherapy and combined treatment with cisplatin and anti-MUC1. In our study we observed that combined treatment of Pt12 with anti-MUC1 had no influence on the induction of programmed cell death in normal cells such as human skin fibroblasts. Also, we showed that cisplatin in dose 20 μM strongly induced apoptosis in human skin fibroblasts. We observed higher concentrations of all tested apoptotic markers such as Bax protein, p53, caspases-3,-8,-9. The proapoptotic effect was weaker after the combined treatment of cisplatin together with anti-MUC1. The obtained results proved that only cisplatin in dose 20 μM induced both apoptotic pathways. It activated the death receptor pathway associated with higher concentration of caspase-8 as well as mitochondrial pathway connected with cytochrome c and caspase-9 releasement. On the contrary cisplatin used together with anti-MUC1 induced only death receptor pathway. We observed higher concentration of caspase-8 in cell lysates as compared with that in control group. The novel Platinum(II) complex together with anti-MUC1 showed no harming effect in the normal cells. Taken together, our results suggest that the combined treatment with anti- MUC1 is a possible way to improve selectiveness of chemotherapeutic agent.
<Keywords: Apoptosis; Proapoptotic Bax; Caspases; Platinum(II) complexes; Combination therapy
The mucin MUC1 is a type I transmembrane glycoprotein expressed at the apical border of healthy epithelia that line the respiratory, reproductive and gastrointestinal tracts. In contrast to normal cells, cancerous tissues aberrantly express the MUC1 protein and it is distributed over the entire tissue surface [1-4]. Its expression is increased by at least 10-fold in most malignant carcinomas [5,6]. Tumour-associated MUC1 has also shorter and less densely distributed O-glycan chains compared to normal MUC1, which is useful as a promising therapeutic target for the treatment of breast cancer patients. The function of MUC1 in the healthy state is still unclear, but its role in cancer metastasis, anti-apoptosis [7-10] and immune suppression [11-13] is well documented [14]. Apoptosis, or programmed cell death is a complex, multistep process, which plays a key role in maintaining the homeostasis of the organism [15]. Induction of apoptosis signal can be emitted by other cells and by exogenous factors: anti-cancer drugs, ionizing radiation or hyperthermia. This process may take place in various ways by starting at the different biochemical pathways. There are known two basic apoptotic signalic pathways in mammalian cells: the extrinsic (receptor) and intrinsic (mitochondrial) pathways. The extrinsic pathway of apoptosis is initiated by FasL binding to the extracellular receptor and results in formation of the DISC that activates caspase-8. In some cells, caspase-8 interacts with intrinsic apoptotic pathway by cleaving Bid and leading to the release of cytochrome c [16]. The intrinsic apoptotic pathway is activated by various intracellular stimuli, including DNA damage, growth factor deprivation, and oxidative stress [17]. The mitochondrial pathway is associated with increased permeability of the mitochondrial membrane and cytochrome c translocation to the cytoplasm. Then cytochrome c binds to the Apaf-1 and procaspase-9, forms the apoptosome and catalyzes the activation of caspase-9. The activation of both pathways activates effector caspases: -3, -6, and -7, which are responsible for irreversible changes in cells. These enzymes have been implicated in the proteolysis of the protein substrates and are capable of inducing the inflammatory process by stimulating the production of proinflammatory cytokines. Researchers are still looking for new compounds that are selective for cancer cells with proapoptotic potential, whereas the least toxic to normal cells. The alkylating agents, including cisplatin, cause numerous side effects; resistance of cancer cells appears and results in the ineffectiveness of the treatment [18-20]. The novel class of compounds are dinuclear platinum(II) complexes which have a different mechanism of action than cisplatin. They bind to the minor groove of DNA and have a particular affinity to the adeninethymine pairs and the weaker to the sequence of guanine-cytosine [21]. New dinuclear berenil-platinum(II) complexes may be an alternative treatment of tumors resistant to cisplatin. Recently we obtained by organic synthesis novel berenil-platinum(II) complex - Pt12 [22,23]. To improve the effectiveness of such a treatment we used Pt12 together with monoclonal antibody against MUC1, which is selective against cancer cells. Such a treatment resulted in the greatest cytotoxicity and induced apoptosis in breast cancer cells in vitro, which was confirmed by several biochemical tests: loss of mitochondrial membrane potential, DNA fragmentation and caspases activation. The combined therapy was more effective compared to the monotherapy: Pt12, anti- MUC1 or reference compound – cisplatin. The higher cytotoxicity is also associated with the inhibition of DNA biosynthesis [22]. The goal of the study was to check the effect of such the combined treatment on programmed cell death in normal cells represented by human skin fibroblasts. The concentration of selected apoptotic markers involved in both apoptotic pathways such as proapoptotic Bax, p53, cytochrome c, caspases-3,-8,-9 was measured in cell lysates of normal cells using ELISA technique.
Materials
Dimethylformamide, K2PtCl4, KI, acetone, 4-ethylpyridine, diethyl ether, methanol, cisplatin, monoclonal antibody anti-MUC1 GP1.4 were purchased from Sigma Chemical Co. (USA). Stock cultures of human skin fibroblasts were purchased from the American Type Culture Collection (USA). Dulbecco’s minimal essential medium (DMEM) and fetal bovine serum (FBS) used in a cell culture were products of Gibco (USA). Glutamine, penicillin and streptomycin were obtained from Quality Biologicals Inc. (USA). ELISA’s kits were purchased from Uscn Life Science Inc. and BioVendor. The chemical synthesis and structure of Pt12 was presented previously [22].
Cell culture
Human skin fibroblasts were maintained in DMEM (Dulbecco`s Minimal Essential Medium) supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 50 U/mL penicillin, 50 mg/mL streptomycin at 37°C in a humidified atmosphere containing 5% CO2. Cells were incubated with anti-MUC1 (10 μg/mL), Pt12 (10 μM), Pt12+anti- MUC1 (10 μM+10 μg/mL), cisplatin (10 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL) for 24 hours and then used to prepare cell lysates. Briefly, trypsinized cells were washed three times with cold PBS and centrifuged at 1000 g for 5 min at 4°C. The cells (1 × 106) were suspended in lysis buffer for whole cell lysates. After centrifugation the supernatants were frozen immediately at -70ºC. The concentration of propapoptotic markers was measured. Cells without addition of compounds were treated as controls.
Determination of proapoptotic Bax protein
The high sensitivity assay kit (Uscn Life Sci Inc.) was used to determine the concentration of proapoptotic Bax protein in cell lysates. The microtiter plate provided has been pre-coated with a monoclonal antibody specific to Bax. Standards and samples were added to the appropriate microtiter plate wells and incubated for 2 hours at 37ºC. After first incubation step a biotin-conjugated polyclonal antibody specific for Bax was pipetted and incubated for 1 hour at 37ºC. After washing away any unbound substances, avidin conjugated to horseradish peroxidase was added to each microplate well and incubated. After another aspiration and washing step a TMB substrate solution was added to each well. The enzyme-substrate reaction was terminated by the addition of a sulfuric acid solution and the color change was measured at a wavelength of 450 nm. The assay was performed in duplicate and the concentration of Bax in the samples was then determined by comparing the O.D. of the samples to the standard curve. Range of the standard curve for Bax was: 0.78-50 ng/ mL. The minimum detectable dose of human Bax was generally less than 0.32 ng/mL.
Determination of caspase-8 and caspase-9
Caspase-8 and caspase-9 concentrations in cell lysates were determined by using an enzyme-linked immunosorbent assay kit (BioVendor). A monoclonal antibodies specific for caspase-8 or caspase-9 have been pre-coated onto a microplate. Standards and samples (100 μL each) were pipetted into the wells in duplicate and antigen was bound by the immobilized antibody. The working solution of antibody was also added to all wells. Then the microplate was incubated for 2 hours at room temperature (RT). After washing away any unbound substances, an enzyme-linked polyclonal antibody specific for caspase-8 or caspase-9 (100 μL) was added to each well for 1 hour at RT. Following a wash to remove any unbound antibody enzyme-reagent, a substrate solution (100 μL) was added to the wells for 15 minutes; the colour developed in proportion to the amount of antigen bound in the initial step. Colour development was stopped by phosphoric acid, and the intensity of the colour was measured at a wavelength of 450 nm. The minimum detectable dose (MDD) of caspase-8 was 0.1 ng/mL and caspase-9 was: 0.4 ng/mL. The concentrations of the samples were calculated from the standard curve and ranged from 0.16-10 ng/mL for caspase-8 and 1.6-100 ng/mL for caspase-9. The results were presented in nanogram per mililiter (ng/ mL). There was no cross-reactivity with other caspases.
Determination of cytochrome c and p53
Cytochrome c and p53 concentrations in cell lysates were detected by using ELISA kit (BioVendor). The microplate was coated by monoclonal antibody specific for cytochrome c or p53. Samples and standards were added into appropriate wells in duplicate and antigen bound to antibodies adsorbed to the microwells. Next step was the addition of a biotin conjugated anti-human cytochrome c or p53 antibody. The plate was incubated for 2 hours at room temperature (RT). Then, unbound biotin-conjugated anti-human cytochrome c or p53 antibody was removed during a wash step. Streptavidin-HRP was added and bound to the biotin-conjugated anti-human cytochrome c or p53 antibody and the microplate was incubated for next 1 hour at RT. After a wash step substrate solution reactive with HRP was added to the wells. A coloured product was formed in proportion to the amount of human antigen present in the sample or standard. The reaction was terminated by addition of acid and absorbance was measured at 450 nm. The concentrations of antigens were calculated from standard curve (for cytochrome c: 0.08-5 ng/mL; for p53: 0.78-50U/mL). The minimum detectable dose for cytochrome c and p53 was 0.05 ng/mL and 0.33 U/mL, respectively.
Determination of active caspase-3
The concentration of caspase-3 was checked using assay kit from Life Sci Inc. Samples and standards (100 μL/well) were added into wells and microplate was incubated for 2 hours at 37°C. After two hours antibody (1:100) was added and plate was incubated for next 60 minutes at 37°C. The washing procedure was performed and polyclonal antibody with enzyme was pipetted and kept for 30 minutes at 37°C. After washing away any unbound substances substrate solution (90 μL/ well) was added. The reaction was stopped by the addition of sulfuric acid. The absorbance was checked at the wavelength 450 nm. The concentration of caspase-3 was calculated from standard curve (0.156- 10 ng/mL). The minimum detectable dose was 0.054 ng/mL. The results were presented in nanogram per milliliter (ng/mL).
Statistical analysis
Experimental data was represented as means ± SD. Each experiment was repeated three times. The differences between control (untreated cells) and treatments were analyzed using one-way ANOVA. All the analyses were performed using GraphPad Prism Version 6.0 (San Diego, CA, USA). A statistically significant difference was defined at p<0.05.
The evaluation of Bax protein, caspases:-8,-9,-3, cytochrome c and p53 concentrations was performed using enzyme-linked immunosorbent assay ELISA. The cells were incubated for 24 hours with the tested compounds. We used compounds alone, anti-MUC1 at two concentrations: 10 and 20 μg/mL, cisplatin and Pt12 in concentrations: 10 and 20 μM. Also we checked the combined effects of anti-MUC1 (10 μg/mL) together with cisplatin (10 and 20 μM), and the anti-MUC1 (10 μg/mL) in combination with Pt12 (10 and 20 μM). The results were compared with the control cells, cultured without drugs. It has been shown that cisplatin at a concentration of 20 μM increased the concentration of Bax (92 ng/ mL) in cell lysates compared with the control (66 ng/mL). The Platinum(II) complex at dose 10 μM caused the lowest release of pro-apoptotic protein. Bax protein concentration after 24 hours of incubation with Pt12 was 64 ng/mL (Figure 1).
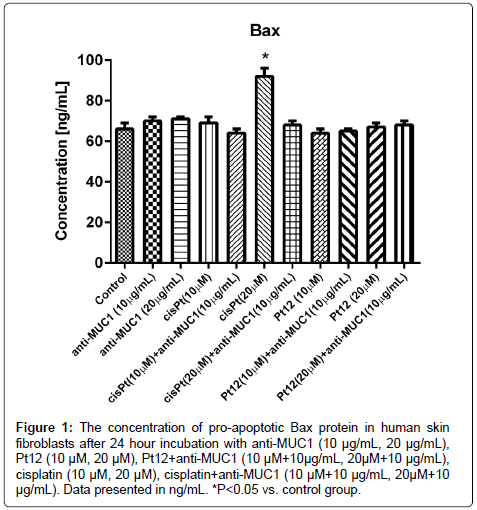
Figure 1: The concentration of pro-apoptotic Bax protein in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
It was observed that cisplatin at a concentration of 20 μM increased the releasement of cytochrome c (40 ng/mL) compared to the control (29 ng/mL). The monoclonal antibody anti-MUC1 used together with Platinum(II) complex caused the lowest release of cytochrome c. After 24 hours of incubation with anti-MUC1 at dose of 10 μg/mL in combination with Pt12 (20 μM) the concentrations of cytochrome c was 29 ng/mL (Figure 2). The similar effect was observed after 24 hour incubation with Pt12 at dose 10 μM.
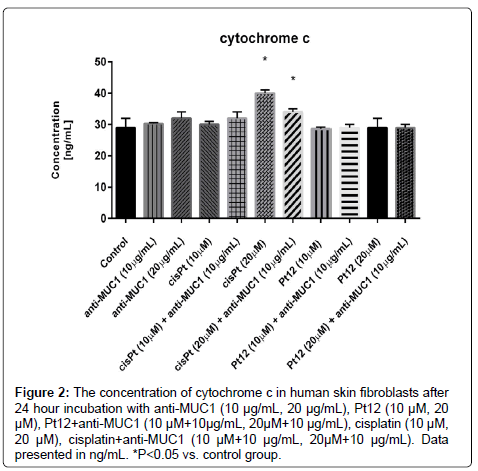
Figure 2: The concentration of cytochrome c in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
After 24 hours of incubation with cisplatin we observed the statistically significant increase of caspase-9 concentration (76 ng/mL) above the control value. The lowest concentration of caspase-9 was detected after Pt12 treatment. The concentration of caspase-9 after 24 hours of incubation was 56 ng/mL, compared to a control, where the concentration was 60 ng/mL, but this difference was not of statistical significance (Figure 3).
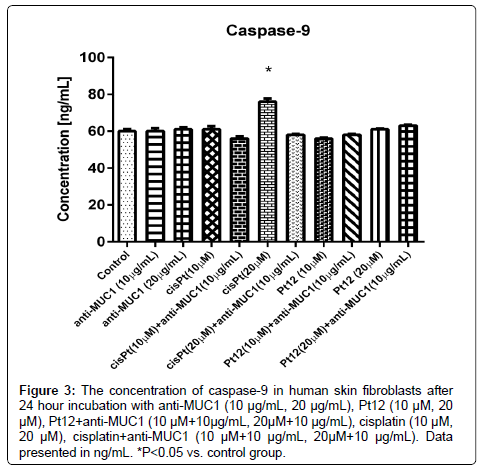
Figure 3: The concentration of caspase-9 in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
The highest concentration of caspase-8 (1.5 ng/mL) was observed after cisplatin treatment alone at dose 20 μM in comparison to control, where concentration was 1.2 ng/mL; however, the difference was statistically insignificant (p>0.05). The lowest concentration of active caspase-8 was demonstrated after 24 hour incubation with anti-MUC1 (10 μg/mL). The concentration of the analyzed apoptotic marker was 1.0 ng/mL (Figure 4).
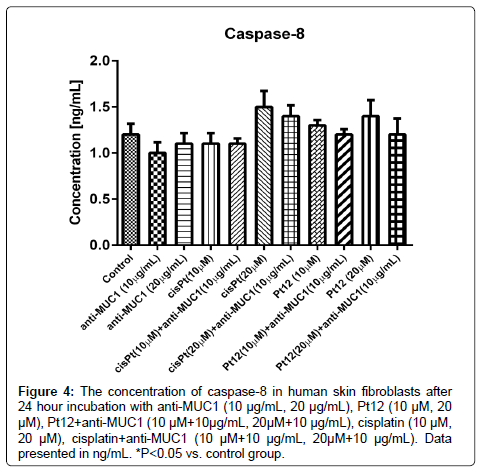
Figure 4: The concentration of caspase-8 in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
P53 is a transcription factor, which regulates downstream genes important in cell cycle and apoptosis [24]. The highest concentration of p53 was observed after 24 hour incubation with cisplatin at dose 20 μM (40 ng/mL). The effect was statistically significant and two times stronger than in control (20 ng/mL) (Figure 5).

Figure 5: The concentration of caspase-9 in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
Finally we determined the concentration of active caspase-3 in cell lysates. The concentration of caspase-3 in control samples was 2.8 ng/mL. After 24 hours of incubation with compounds used in monotherapy and in combination with monoclonal antibody we proved that all differences between control vs. other treatments were statistically insignificant (p>0.05) (Figure 6).
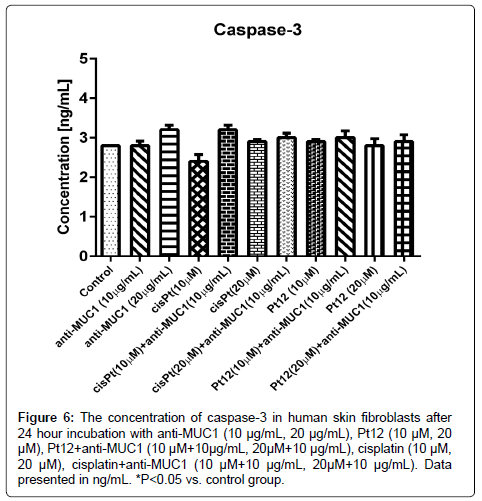
Figure 6: The concentration of caspase-3 in human skin fibroblasts after 24 hour incubation with anti-MUC1 (10 μg/mL, 20 μg/mL), Pt12 (10 μM, 20 μM), Pt12+anti-MUC1 (10 μM+10μg/mL, 20μM+10 μg/mL), cisplatin (10 μM, 20 μM), cisplatin+anti-MUC1 (10 μM+10 μg/mL, 20μM+10 μg/mL). Data presented in ng/mL. *P<0.05 vs. control group.
Apoptosis and necrosis represent two different forms of cell death. Necrosis is nonspecific form of cell death with localized response and damage to surrounding cells and tissue. Apoptosis is a therapeutic strategy to kill tumor cells without comprising normal cell function [25]. Recently we proved that combined treatment (Pt12+anti-MUC1) caused the highest releasement of the pro-apoptotic markers in MDA-MB-231 was breast cancer cells [26]. The concentration of Bax, caspase-3,-8 and -9 statistically increased in cell lysates after 24 hour incubation with drugs used in combination. The results from combined treatment were compared with those obtained using monotherapy. The effect was stronger than treatment with Pt12, cisplatin, anti-MUC1, and anti-MUC1 used with cisplatin. We also observed the highest decrease of mitochondrial membrane potential and the strongest DNA fragmentation after such a combined treatment. Our study proved that Pt12 together with anti-MUC1 strongly induced apoptosis in estrogen receptor negative breast cancer cell line (MDA-MB-231) by activation of intrinsic and extrinsic apoptotic pathways [26]. In this study we wanted to check the influence of Pt12 together with anti-MUC1 on concentration of selected apoptotic markers in normal cells compared to monotherapy (Pt12, cisplatin, anti-MUC1) and combined treatment (anti-MUC1 used with cisplatin). One of the primary regulators of the mitochondria-mediated pathway to apoptosis is the family of Bcl-2 proteins [27,28]. These proteins are characterized into proapoptotic members that include Bax, Bak, Bik, Bad, and Bid and anti- apoptotic proteins such as Bcl-2 and Bcl-xL, based on their ability to suppress or induce the release of cytochrome c [25]. In our study we proved that only cisplatin at dose 20 μM significantly increased the level of proapoptotic protein Bax, which indicated that mitochondria seem to be a target of the alkylating agent. After other treatments the concentration of Bax was similar to control value. The intrinsic mitochondrial pathway is characterized by the rapid release of cytochrome c from the mitochondrial inter-membrane space into the cytosol and it is associated with a loss of outer mitochondrial membrane potential [29], and is tightly regulated by the pro- and anti-apoptotic members of the Bcl-2 family [25]. In our study we observed that combined treatment of Pt12 with anti-MUC1 did not release the apoptotic markers in normal cells. Also we showed that cisplatin in dose 20 μM strongly induced the releasement of other tested apoptotic markers such as p53, caspases-3,-8,-9 in fibroblast lysates. The final pathway that leads to execution of the death signal is the activation of a series of caspases. The intrinsic and extrinsic apoptotic pathways converge to caspase-3 and the upstream caspases that converge to caspase-3 are caspases-9 and -8 in the intrinsic and extrinsic pathways, respectively. The downstream caspases lead to the morphologic manifestations of apoptosis, such as DNA condensation and fragmentation, and membrane blabbing [24,30,31]. In our research the pro-apoptotic effect was weaker after combined treatment of cisplatin together with anti-MUC1. The obtained results proved that only cisplatin in dose 20 μM induced both apoptotic pathways. It activated the death receptor pathway associated with higher concentration of caspase-8 as well as mitochondrial pathway connected with cytochrome c and caspase-9 releasement. The mechanism of action initiated by cisplatin used together with anti-MUC1 induced only death receptor pathway. We observed higher concentration of caspase-8 in cell lysates as compared with that in control group. Our study showed that combined treatment is more selective and in weaker rate induced programmed cell death in normal cells such as human skin fibroblasts.
The combined treatment with anti-MUC1 is a possible way to improve selectiveness of chemotherapeutic agent. Monoclonal antibody against MUC1 sensitizes breast cancer cells and improves the effectiveness of such a therapy, but in normal cells used in combination with cytotoxic agents may play a role as a “selective” inducer of apoptotic markers. The novel Platinum(II) complex together with anti- MUC1 had no influence on induction of programmed cell death in normal cells such as human skin fibroblasts.
This investigation was supported by research grant 154-29585 F from Medical University of Bialystok, Poland.