Journal of Antivirals & Antiretrovirals
Open Access
ISSN: 1948-5964
ISSN: 1948-5964
Research Article - (2016) Volume 8, Issue 2
Some anti-infective drugs exhibit also anti-tumor activity. The anti-tumor activity of anti-infective drug (AID) containing a complex of molecular iodine with polypeptides, α-dextrins and lithium halogenides, was studied in human and murine tumor cell lines in vitro.
The experiments were performed with human (HeLa and K562) and murine (L5178Y) tumor cell lines. As a control was used dog kidney epithelial cell line MDCK. The anti-tumor activity of AID was assessed by measuring of IC50. The drug was used at the concentrations of 500 μg/ml; 250 μg/ml; 125 μg/ ml; 63 μg/ml; 32 μg/ml; 16 μg/ml; 8 μg/ml; 4 μg/ml; 2 μg/ml; 1 μg/ml; and 0.5 μg/ml. It was found that the IC50’s were 112 μg/ml, 11.8 μg/ml, 10.3 μg/ml and 40.6 μg/ml for Hela, K562, L5178Y and MDCK cells, respectively.
The probable mechanism of the drug’s cytotoxic activity is explained by the results of molecular modeling process and DFT calculations.
AID interacts with the onco-DNA, and the following inhibiting complexes are formed: lithium (Li) halogenide included in the AID, forms a complex with the phosphate group while the molecular iodine is coordinated by the adenosine or guanosine nitrogenous bases and Li halogenide.
These inhibitory centers disturb the interaction of topo I active site with phosphate group, and two new nucleoprotein complexes are formed. In one the Arg amino acid residue is bounded by the Li halogenide complex with the phosphate group, in the second one Tyr is bounded by the adenosine complex with molecular iodine and Li halogenide.
Keywords: Anti-Cancer Activity, α-Dextrins, DNA, anti-infective drugs
DNA topological modifications play an important role in the manifestation of its functional activities such as replication, transcription, and recombination. The topological state of the cell DNA modifies and regulate the DNA topoisomerase. They act on the DNA isomerizing or changing its topology.
Topoisomerases fall into two basic types according to their mechanism of action. Topo I enzymes do not require the presence of energy-rich cofactors, such as ATP (adenosine triphosphate). Topo II type enzymes produce a temporary double-stranded break, hydrolyzing ATP.
The study of eukaryotic topoisomerases of type I and II is largely stimulated by the fact that these enzymes are a target for a variety of anticancer drugs [1-3].
Some anti-infective drugs also exhibit anticancer activity [4-6]. Among the anti-infective drugs containing molecular iodine, a special place is occupied by compound called “Armenicum” [7,8] and a newly synthesized anti-infective drug (AID) [9]. In addition to the molecular iodine complex, these drugs contain potassium and lithium halogenides.
Drugs containing molecular iodine complexes with organic ligands are very toxic, and therefore are used only topically. As part of the antiinfective drugs [7-9] the molecular iodine is in a specific active form, and because of that when it is used internally, the toxic effects in humans minimizes.
We have shown [10] that the electronic structure of I2 in active complex (AC) of AID differs from that in complexes with organic ligands or I2 in its free form. In the ACs the molecular iodine displays acceptor (donor) properties towards polypeptide (lithium halogenides).
Toxicological research of drugs has shown that the classification Globally Harmonized Classification System can be attributed to the 4 categories [11].
In our resent study [12] we have shown that anti-infective drug (AID) inside the α-dextrin helix contains three active centers: molecular iodine, coordinated by polypeptide lithium halogenides (LiCl (I) -I2-α-dextrin-peptide complexes), triiodide and lithium halogenides.
The results of our study published in 2014 [13] showed that the combined action of the drug containing LiCl (I)-I2-α-dextrin-peptide complexes and doxorubicin, significantly enhances the antitumor activity of doxorubicin on the growth of Ehrlich ascites carcinoma. We explained the mechanism of this phenomenon, based on the results of quantum chemical molecular modeling.
The result of this study presents experimental data obtained in murine and human tumor cells line in regards of the anticancer activity of AID. The mechanism of anti-cancer activity is discussed and molecular modeling of the key structures is carried out.
Understanding the mechanism of anti-cancer activity of the AID may help find its new versions with improved anti-tumor properties.
It is shown that LiCl (I) I2-α-dextrin-peptide complexes form complexes with the nitrogenous bases of the onco-DNA displacing the peptide, while the lithium halogenides form complexes with the phosphate groups. These complexes become inhibiting centers for amino acid residues such as arginine (Arg) and tyrosine (Tyr) which are parts of the topo I active site. In these structures, Arg and Tyr are bounded to inhibitory centers by coordination bonds stronger than hydrogen bonds.
Studies on the noncovalent interactions of small molecules with DNA continue to be a fruitful area for the discovery of potential new tumor agents [14]. In contrast to the already known compounds AID' s active centers bind to the DNA with coordination bonds stronger than stacking interactions.
Experimental models
For the research are used the following mouse and human tumor cells lines: HeLa (human cervical adenocarcinoma, ATCC-CCL-2, USA), L5178Y (mouse lymphoma, ATCC-CRL-9518, USA), K562 (human erythroblastoid leukemia, ATCC-CCL- 243, USA). As a control has been used MDCK cell line (dog kidney epithelial cells, Cell Biotechnology Laboratory, Russia).
Test substances
Tumor and normal cell lines were treated with AID substances with the following final concentrations: 500 μg/ml; 250 μg/ml; 125 μg/ ml; 63 μg/ml; 32 μg/ml; 16 μg/ml; 8 μg/ml; 4 μg/ml; 2 μg/ml; 1 μg/ml; and 0.5 μg/ml.
Defrozing and culturing tumor cell lines
Cell lines were removed from liquid nitrogen, washed with 20% complete growth medium containing 20% FBS, 4 mM L-glutamine, and 100 mg/ml of AAS, by centrifugation at 300 * g for 5 min. After washing, the cells were resuspended by 1 ml of 20% complete growth medium for cell viability quantification.
The culture medium was replaced every 2-3 days according to the cancer cells specifications. Culturing of the cells was carried out in a CO2-incubator at 37° C, 5% CO2 and 95% humidity [15].
The monolayer cell culture covering over 80% of growth flask can be recultured into the fresh flask or 96 well plates by dissociation of the monolayer cells with trypsin solution.
Dissociation of monolayer cell culture is best achieved by washing the monolayer cells with Hanks' balanced salt solution (1x HBSS) followed by incubation with trypsin solution (0.25% crude or 0.01-0.05% purified) for 30 seconds.
After that the trypsin is removed by centrifugation, cells are counted, suspended in a medium, determine the number and cultured into new flasks.
Cell suspension requires neither washing nor trypsin treatment, since the cells do not adhere to growth flasks.
They can be gathered into a container and centrifuge to remove the old growth medium, calculate the required number of cells and culture onto a fresh growth flask with a fresh 10% complete growth culture medium.
Monitoring of cell viability
To determine the number of living cells in 1ml, 20 μl of cell suspension were collected to 1.5 ml eppendorf tube and 20 μl of 0.4% trypan blue was added (in ratio 1:1). Then the cells were placed under a cover glass in the Goryaev chamber, the total number of cells was counted and the percentage of viable cells was determined by the addition of trypan blue. Trypan blue penetrates into dead cells, binding to the proteins of the plasma membrane and stains them blue [15].
Unstained cells were counted and the percentage of living cells was calculated by the formula below (1):
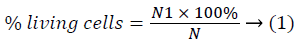
Where: N – the total number of cells in 5 small squares;
N1 – the number of unstained cells in 5 small squares.
The concentration of living cells was calculated by the formula below (2):
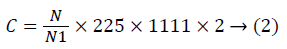
Where: C – the total number of cell per ml;
N – the number of cells in 5 squares;
N1 – the number of squares;
225 – the total number of cells in the chamber;
– Goryaev chamber coefficient;
2 – the number of dilutions.
Methods of studying the cytotoxicity of substances in vitro
The biological activity of the medications under the study was assessed by using the MTT colorimetric method [16].
This method is based on the use of a water soluble vital dye yellow bromide 3-(4.5-dimethylthiazol-2-yl)-2.5-tetrazoliyabromid (MTT), which allows to determine the viability of cell cultures due to the ability of living cells to convert the soluble yellow MTT to insoluble purple-blue intracellular formazan crystals.
Dead cells do not have this ability. Cell lines were cultured in 96- well plates (BD Biosciences) in the desired concentration (5*104 cells per 1 well). Plates were incubated in an incubator at 37.0°C, 5% СО2.
The growth medium was removed from the wells after 24 hours of incubation, and 100 μl of the medium and 100 μl of the material were placed into the well in the test concentrations. 200 μl of growth media were added to the wells containing negative control.
After 40-44 hour exposure to the test medications 20 μl of the MTT solution were added to each well and incubated for further 4 hours in a CO2-incubator at 37°C, 5% CO2 with 95% humidity.
After incubation time completion, the supernatant was removed. And 100 μl of DMSO were added to each well. The optical density of dissolved formazan was measured photometrically using the Sunrise RC.4 Microplate Reader with the length of the main filter of 540 nm and the reference wavelength of 620 nm. This makes possible to determine the number of living cells after incubation with the medication. This test takes into account the advanced stage of cell death [16].
Cytotoxicity of AID in tumor cells
The direct effect of AID has been studied in human (HeLa and K562) and murine L5178Y and tumor cell lines. The results of these experiments are presented in the following Tables 1-5.
| Tubes No | Final concentration of AID, µg/ml | % Viability | Inhibition concentration 50% (IC50) |
|---|---|---|---|
| 1 | 1000 | Stock solution | 112 µg/ml |
| 2 | 500 | 10.0±0.4 | |
| 3 | 250 | 23.1±1.4 | |
| 4 | 125 | 44.1±0.2 | |
| 5 | 63 | 68.9±1.7 | |
| 6 | 32 | 77.4±2.4 | |
| 7 | 16 | 78.4±2.3 | |
| 8 | 8 | 78.7±1.7 | |
| 9 | 4 | 81.4±0.4 | |
| 10 | 2 | 87.6±2.8 | |
| 11 | 1 | 88.7±0.4 | |
| 12 | 0.5 | 101.5±3.0 |
Table 1: Monotreatment of AID on HeLa cell line.
| Tubes No | Final concentration of AID, µg/ml | % Viability | Inhibition concentration 50% (IC50) |
|---|---|---|---|
| 1 | 1000 | Stock solution | 11.8 µg/ml |
| 2 | 500 | 8.2±0.8 | |
| 3 | 250 | 10.1±1.1 | |
| 4 | 125 | 11.0±0.6 | |
| 5 | 63 | 12.2±0.7 | |
| 6 | 32 | 12.4±1.6 | |
| 7 | 16 | 39.6±1.4 | |
| 8 | 8 | 59.4±0.8 | |
| 9 | 4 | 100.4±1.7 | |
| 10 | 2 | 101.8±0.7 | |
| 11 | 1 | 103.4±1.4 | |
| 12 | 0.5 | 104.3±1.1 |
Table 2: Monotreatment of AID on K 562 cell line.
| Tubes No | Final concentration of AID, µg/ml | % Viability | Inhibition concentration 50% (IC50) |
|---|---|---|---|
| 1 | 1000 | Stock solution | 10.3 µg/ml |
| 2 | 500 | 3.1±1.4 | |
| 3 | 250 | 8.5±2.3 | |
| 4 | 125 | 11.4±1.9 | |
| 5 | 63 | 14.7±1.3 | |
| 6 | 32 | 16.0±0.7 | |
| 7 | 16 | 25.1±1.5 | |
| 8 | 8 | 59.8±0.9 | |
| 9 | 4 | 64.7±3.0 | |
| 10 | 2 | 73.1±1.8 | |
| 11 | 1 | 92.6±2.3 | |
| 12 | 0.5 | 98.0±0.5 |
Table 3: Monotreatment of AID on L 5178Y cell line.
| Tubes No | Final concentration of AID, µg/ml | % Viability | Inhibition concentration 50% (IC50) |
|---|---|---|---|
| 1 | 1000 | Stock solution | 40.6 µg/ml |
| 2 | 500 | 1.7±1.8 | |
| 3 | 250 | 4.4±0.5 | |
| 4 | 125 | 4.6±0.9 | |
| 5 | 63 | 67.3±1.1 | |
| 6 | 32 | 73.3±1.8 | |
| 7 | 16 | 79.6±0.5 | |
| 8 | 8 | 84.4±1.0 | |
| 9 | 4 | 86.9±2.3 | |
| 10 | 2 | 82.9±1.1 | |
| 11 | 1 | 90.0±1.7 | |
| 12 | 0.5 | 96.5±3.1 |
Table 4: Monotreatment of AID on MDCK cell line.
| Drug | IC50 | |||
|---|---|---|---|---|
| Cell Lines | ||||
| HeLa | K562 | L5178Y | MDCK | |
| AID | 112 µg/ml | 11.8 µg/ml | 10.3 µg/ml | 40.6 µg/ml |
Note: IC50 is a cytotoxic concentration of the agent that leads to the death of 50% of cells
Table 5: Cytotoxicity of AID in human and murine tumor cells.
As can be seen in Table 5, the cytotoxic effect of AID at relatively low concentrations was seen only in two tumor cell lines, namely K562 and L51178Y.
The IC50’s were equal to 11.8 μg/ml and 10.3 μg/ml, respectively. The IC50 for HeLa cells was almost 3-fold higher than that for MDCK cells. The toxicity of AID substances on MDCK normal cells has been observed at concentrations of 41 μg/ml. Hence, the cytotoxic effects of AID on tumor cell lines K562 and L5178Y was determined at 4-fold lower concentrations than the control ones. But at the same time the drug was ineffective against HeLa cells (IC50 is almost 3 fold higher than that for the control –MDCK cells).
This shows the possibility to use the AID as antitumor drugs against certain cancer cells, namely against leukemia.
Computational details
An adequate description of the associates involved in the reaction, including a large number of molecules that are bound by hydrogen bonds, requires the electron correlation to be considered. A detailed analysis of CCSD(T) and DFT (BLYP, BPW91, B3LYP, B3PW91)- calculations of dissociation energies, bond lengths, harmonic frequencies obtained for polyiodide-anions and comparision with experimental data (IR, Raman spectroscopy) is presented in [17].
So the DFT/B3PW91 method with the 6-31G ** basis set for the atoms of C, N, O, H, Li, Cl and the midi basis set for I was applied for structures Ia,b-IXa,b. Spatial structures Ia, b-IXa, b were calculated with full geometry optimization.
In [12] we used UV and IR spectroscopy to investigate water solutions of the systems containing lithium and potassium halogenides, molecular iodine, and amine acid. Using UV spectroscopy, we investigated the interaction of LiCl(I)-I2-α-dextrinpeptide with the AGA nucleotide triplet. The geometry of systems containing molecular iodine, organic compounds or AGA nucleotide triplet, lithium halogenides adequately reproduced by method DFT/ B3PW91. The calculations of the spectral parameters of the proposed structures are in good agreement with the experimental data of UV and IR spectral investigations.
PM3 [18] semi-empirical method was used to calculate the complex molecular iodine with the nucleotide triplets. Spatial structures of complexes were calculated with full geometry optimization.
The human topo I consists of the four main domains: N-terminal and core domains, a linker domain and C-terminal domain [19]. The C-terminal domain together with the core domain can provide of almost complete enzymatic activity of topo I [20].
The relaxation reaction catalyzed by IB-type topoisomerases is initiated by a nucleophilic attack of the DNA phosphodiester bond by the tyrosine amino acid residue which is located at the active site of the enzyme.
The mechanism of a DNA phosphodiester bond cleavage was suggested based on the results of X-ray analysis, and the results of the impact on the activity of enzymes of amino acid substitutions were made using site-directed mutagenesis [21].
The interaction of amino acid residues of the topo I active site with the splitted phosphoester group of DNA is shown in Figure 1. The nucleophilic attack by the tyrosine OH-group of the О5′-Pphosphoester bond leads to the formation of a covalent bond between Tyr and 3′-end of the splitted DNA. The collinear arrangement of the tyrosine hydroxyl group with respect to the О5′-P-phosphoester bond creates conditions for a nucleophilic attack of that bond by the tyrosine OH-group and the formation of a covalent bond between Tyr and 3′- end of the splitted DNA.

Figure 1: The interaction of amino acid residues of the active site of the human topoisomerase I with the splitted phosphoester group of DNA.
The formation of a hydrogen bond of Arg488 and Arg532 with one of the oxygen atoms of the internucleoside phosphate group is required for stabilization of the transition state. The other oxygen atom of the phosphate group forms a hydrogen bond with His632. His632 only coordinates the phosphate group in the active site.
Inside the α-dextrin helix, lithium halogenides are coordinated by OH groups of α-dextrin and donor-active groups of polypeptide. The interaction energy of lithium halogenide with phosphate group is greater than with the donor-active groups included of polypeptides or α-dextrin. Therefore, when AID interacts with onco-DNA phosphate groups coordinate lithium halogenide (Figure 2, complexes Ia and Ib). In [22] lithium chloride is shown to inhibit the growth of tumor cells.

Figure 2: The interaction of AID active centers with onco-DNA.
It was shown in our recent paper [12] that during the interaction of the AID on DNA nucleotides its active site interact with the nitrogenous base. The adenosine and guanosine nitrogenous bases displace the polypeptide and form complexes with the molecular iodine and lithium halogenide (IIa, b-IIIa and b complexes) (Figure. 2).
The interatomic distance in I2 molecule is~ 2,77Å in structures IIa, b-IIIa, b, therefore in this complexes the lithium halogenide is located behind the sugar-phosphate moiety and becomes available to the amino acid residues of the topo I active site.
Structures Ia, b-IIIa, b may become centers of inhibition of the topo I active site, if the interaction energy of arginine and tyrosine amino acid residues that are the most important for the cleavage of their phosphoester bond with Ia,b-IIIa,b complexes is greater than that with the phosphate group or the complex of the topo I active site.
Structures Ia, b-IIIa, b may become centers of inhibition of the topo I active site, if the interaction energy of arginine and tyrosine amino acid residues that are the most important for the cleavage of their phosphoester bond with Ia,b-IIIa,b complexes is greater than that with the phosphate group or the complex of the topo I active site.
At the time of a nucleophilic attack of topo I on the DNA the Arg488 and Arg532 amino acid residues are bound by hydrogen bonds to one of the oxygen atoms of the phosphate group (Figure 3 structure IV). However, in this structure, the energy of interaction (ΔE1) of Arg488 and Arg532 with the phosphate group is weakened by electrostatic repulsion of the positively charged Arg488 and Arg532.
In the interaction of lithium halogenide with phosphate group it is energetically more favorable (~ -65 kсal/mole) when the lithium halogenide displaces one of the amino acid residues Arg488 from the coordination sphere of the phosphate group, and forms a complex with the other one (Figure 3 Va, b structures).

Figure 3: The interaction energy (ΔE, kcal / mol) of Arg inside active site and with Ia,b-IIIa,b complexes:
Figure 3 shows the interaction energy of the arginine amino acid residue with two inhibitory centers: lithium halogenide forms a complex with a phosphate group (Va, b structures), and molecular iodine and lithium halogenide forms complexes with adenosine (structures VIa, b) and guanosine (structures VIIa, b).
In our calculations, α-dextrin is replaced with ethanol while the sugar moiety bound to the phosphate group is replaced with two methyl groups.
The interaction energies are calculated as shown below
ΔE1 = Etot.(IV) - Etot.(PO4 (CH3)2Na) - 2Etot.(Arg)
ΔE2,3 = Etot.(Va, b) - Etot.(Ia, b) - Etot.(Arg)
ΔE4,5= Etot.(VIa, b) - Etot.(IIa,b) - Etot.(Arg)
ΔE6,7= Etot.(VIIa,b) - Etot.(IIIa,b) - Etot.(Arg)
In the DNA structure, the phosphate group is at ∼7.8 pH in its ionized state, with the charge of -1. However, the metal ions (such as Na+, and K+ ions present in the internuclear environment form a complex with the phosphate group and the negative charge is neutralized.
Barone et al. [23] using DFT-D/BLYP-D level of theory have shown that sodium ions as counter ions, make a significant contribution to the stability of the DNA double helix, because they reduce electrostatic repulsion of the phosphate groups and strengthen the π-π stacking interaction of Watson- Crick pairs. We have considered the case of compensating the charge of the phosphate group by Na+ ion.
Calculations have shown that the interaction of Arg with Ia, b-IIIa, b complexes is energetically favorable. As can be seen in Figure 3 the binding of arginine by the phosphate group coordinated by lithium halogenide (complexes Ia and Ib) is energetically more preferable than by nitrogenous nucleotide bases (adenine and guanine), coordinated by molecular iodine and lithium halogenide (complexes IIa, IIb and IIIa, IIIb).
The interaction of the arginine acid residue with IIa, b-IIIa, b complexes results in the formation of a nucleoprotein complex, with the molecular iodine, coordinated by the lithium halogenide, becoming its center, and binding the nucleotide and the arginine amino acid residue (VIa, b-VIIa, b structures).
It should be noted that in the most stable structures the HCl (I) ion pair leaves the coordination sphere of the lithium ion and forms a coordination bond with the oxygen atom of cytidine or thymine. The Li-ion is coordinated by ethanol, molecular iodine and a neutral molecule of arginine.
In the active site of topo I the tyrosine amino acid residue is bound with Arg590 through a water molecule (Figure 4, VI structure). Figure 4 shows the interaction energy of the tyrosine amino acid residue in the active site of topo I and the interaction energy of tyrosine with two inhibitory centers.

Figure 4: The interaction energy (ΔE, kcal/mol) of the tyrosine amino acid residue inside active site and with I (a, b) -III (a, b) complexes.
The the interaction energies are calculated as shown below:
ΔE8 = Etot.(VIII) - Etot.(Tyr) - Etot.(H2OArg)
ΔE9,10 = Etot.(IXa,b) - Etot.(Ia,b) - Etot.(Tyr)
ΔE11,12= Etot.(Xa,b) - Etot.(IIa,b)- Etot.(Tyr)
ΔE13,14= Etot.(XIa,b) - Etot.(IIIa,b)- Etot.(Tyr)
As can be seen in Figure 4 the interaction of tyrosine with Ia, b-IIa, b inhibitory complexes is energetically more favorable than with ArgH2O (Figure 4).
The interaction of tyrosine with the adenosine-molecular iodinelithium halogenide IIa, b results in the formation of Xa,b structures, in which a high negative charge (-1.282) is transferred from the tyrosine to the lithium halogenide. As a result, a high negative charge appears on the ion of chlorine (or iodine) ( ~ 0.400).
The ion of chlorine (or iodine) becomes more donor active than the nitrogen atom in the five-membered ring structure of adenosine. Because of that the molecular iodine in Xa,b structures forms a complex with lithium halogenide.
The interatomic space in the Li+ I2Cl or (I3) ion pair show that a new ion pair of lithium ion and triiodide or I2Cl interhalogenide is formed. In Xa,b structure the lithium ion is coordinated by tyrosine, ethanol and I2I (Cl)- ion.
The interaction of tyrosine with the guanosine-molecular iodinelithium halogenide (IIIa, b complexes) complex neither triiodine, nor interhalogenide is formed. In the XIa,b structures a hydrogen bond is formed between the chlorine ion and the atom of hydrogen in the C-H bond of the guanosine five-membered ring.
Part of the charge transferred from the tyrosine to the lithium halogenide is transferred from the latter to guanosine. Therefore, the nitrogen atom in the five-membered ring becomes more donor active than the ion of chlorine (iodine). In the XIa, b structures the molecular iodine is coordinated by the nitrogen atom in the five-membered ring of guanosine.
The interaction energy of tyrosine with IIIa,b complexes is less than with the water molecule and arginine, therefore IIIa,b complexes (the guanosine-molecular iodine-lithium halogenide complexes) cannot inhibit tyrosine.
Tyrosine can be inhibited only by IIa, b complexes (the adenosinemolecular iodine-lithium halogenide complexes). The binding of tyrosine by IIa, b complex is energetically more preferable than that by Ia,b complex (lithium halogenide complex with the phosphate group).
Experimental data suggest that the anti-cancer effect of the AID is selective: it reveals itself towards K562, L5178V cancer cells, but has no (or very low) anti-cancer effect against HeLa cancer cells.
This effect requires further investigation, but perhaps one of the reasons for the selective anti-cancer effect of the AID discovered by us, is the selective effect the interaction of molecular iodine with nucleotide triplets.
Molecular iodine complexes with nucleotide triplets may contain from one to three molecules of iodine. PM3 semi-empirical method was used to study the effect of the number of iodine molecules and the structure of the nucleotide triplet on the stabilization energy of complexes of iodine with nucleotide triplets.
Calculations have shown that in cases where the complex contains three iodine molecules, the most stable complexes are formed. In the Watson-Crick pair the nitrogen atom if cytosine is bound by a hydrogen bond with the guanosine N-H bond, while the thymidine atom of oxygen is bound the with NH2-group of adenosine.
Spatially available for the formation of complexes are only nitrogen atoms of the five-membered ring of adenosine and guanosine. Therefore, the most stable iodine complexes are formed with nucleotide triplets containing only adenosine and guanosine, since only these nucleotide triplets may form complexes with as many as three molecules of iodine.
Table 6 shows the stabilization energy of iodine complexes with nucleotide triplets, containing three iodine molecules.
| Nucleotide Triplets | ∆E |
|---|---|
| GGG | -28.80 |
| AGA | -29.26 |
| AGG | -6.69 |
| AAA | -26.33 |
| GAA | -10.48 |
| GAG | -2.86 |
Table 6: The stabilization energy (ΔE, kcal / mol) for the complexes of iodine with nucleotide triplets containing three iodine molecule.
As can be seen in Table 6 the stability of the complexes varies greatly and depends on the structure of the nucleotide triplet. When the concentration of the drug, is limited by the maximum allowable dose, iodine complexes will be formed with only four nucleotide triplets: GGG, AGA, GAA, AAA, while the probability of the formation of complexes depends on the stabilization energy.
Experimental research have shown the possibility to use the AID as antitumor drugs against certain cancer cells, namely against leukemia.
Calculations have shown that the action of AID on onco-DNA results in the formation of the following inhibitory complexes: the halogenide lithium-phosphate group complex (Ia,b structure), and complexes in which molecular iodine coordinates nitrogen bases of adenosine or guanosine and lithium halogenide (IIa, b-IIIa, b structure).
Under the action of these inhibiting centers the interaction of the topo I active site with the phosphate group is broken. Two new nucleoprotein complexes are formed, in which the most important arginine and tyrosine amino acids of the topo I active site are bound by Ia, b-IIa, b complexes. Arginine is bounded by the lithium halogenide phosphate group complex and tyrosine is bounded by the adenosinemolecular iodine-lithium halogenide complex.
Calculations were made using a supercomputer at the National Laboratory for Information and Space Technologies (KazNRTU JSC).