Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
![]() +44 1478 350008
+44 1478 350008
ISSN: 2329-9029
![]() +44 1478 350008
+44 1478 350008
Research Article - (2014) Volume 2, Issue 2
The pentacyclic triterpenoids from birch (Betula platyphylla suk) have broad pharmacological activities and can be potentially used for the development of anti-cancer and anti-AIDS drugs. In this study, we explored the effects of spraying 3-year old white birch with different concentration of methyl jasmonate (MeJA) and salicylic acid (SA) on the expression of key genes in triterpenoid biosynthesis pathways and on the accumulation and physiological characteristics of triterpenoids in birch saplings. The results show that, spraying different concentration of MeJA and SA could obviously promote accumulation of total triterpenoids in 3-year old white birch. The triterpenoid content in the stem bark was significantly increased after 1d of treatment with 1 mmol·L-1 MeJA (MJ2) and after 14 d of treatment with 5 mmol·L-1 SA(SA1), triterpenoid contents of MJ2 and SA1 reaching 81.86 mg/g and 91.40 mg/g, increased by 46.11% and by 45.07% compare control(CK), respectively. In addition, MeJA and SA treatment increased the contents of chlorophyll a and b, antioxidant enzymes superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT), and photosynthetic performance, and affected the content of soluble sugar and soluble protein in birch leaf. Fluorescence quantitative polymerase chain reaction (qPCR) results showed that MeJA and SA treatment deferentially enhanced the expression of key genes farnesyl diphosphate synthase (FPS), cycloartenol synthase
(BPX and BPX2), lupeol synthase (BPW) and beta-amyrin synthase (BPY) in triterpenoid synthesis pathway in birch bark and leaves. The results showed that MeJA and SA induced triterpenoid synthesis of birch plant is closely related with not only the expression of key genes of triterpenoid synthesis pathway but also photosynthesis, anti-stress response and physiological indexes, suggesting regulation of triterpenoid synthesis of birch by MeJA and SA may involve in more complex mechanisms at physiological and molecular level.
Keywords: Birch; Triterpenoid; Physiological characteristics; Gene expression; SA; MeJA
OSC: Oxidosqualene Cyclase; β-AS: β-Amyrin Synthase (BPY); LUS: Lupeol Synthase (BPW); CAS: Cycloartenol Synthase (BPX)
Extractions from birch (Betula platyphylla suk.) have broad application potentials in medicine, cosmetics and health aspects [1-6]. Its rind contains many different types of triterpenoid compounds, mainly betulin and betulinic acid of lupeol type, oleanolic acid of oleanane type and β-sitosterol of cycloartenol type [7-8].
Biosynthesis of secondary metabolites in plants is more complex compared with primary metabolism [9-13]. It is tightly regulated by cellular factors and easily influenced by environmental factors. The latter could regulate transcription of key enzymes involving secondary metabolism process, and subsequently change metabolic flux and reaction rate of secondary metabolic pathways, leading to enhanced production of secondary metabolites [14-17]. Methyl jasmonate (MeJA) and salicylic acid (SA) are naturally occurring plant compounds and could regulate several plant physiological processes in response to pathogen attack, wounding, and ozone [18-21]. Exogenous application of MeJA and SA triggers defense responses that resemble those initiated by pathogen and also modulates the production of certain secondary metabolites in a variety of plant species by inducing transcription of genes important to secondary metabolism process and promoting synthesis of enzymes responsible to production of many plant secondary metabolites such as paclitaxel, camptothecin, isoflavone, ginseng saponin, silymarin trans-resveratrol, among others [21-27]. Many studies have confirmed that MeJA and SA can promote the accumulation of a variety of plant secondary metabolites, but the physiological and molecular mechanisms that MeJA and SA induce secondary metabolism products are not completely clear. In this paper, we systematically studied the effects of MeJA and SA on the synthesis of photosynthesis products, activity of antioxidant enzymes, and expression of genes key to triterpenoid synthetic pathways in birch.
Plant materials
The experiments were conducted in Maoershan Experimental Base of Northeast Forestry University (Haerbin, China) in July and August, 2011. Three-year-old white birch (Betula platyphylla suk.) seedlings were planted in plastic pots with diameter of 25 cm and height of 35 cm, containing 10 kg of soil mixed with peat, nursery soil : turfy soil :sand at ratio of 2:1:1 (V/V/V) under conventional management with same watering quantity for each pot.
MeJA and SA treatment
Seedlings with consistent growth were chosen for MeJA and SA treatment. Jasmonic acid (MeJA) and salicylic acid (SA) (Sigma; St. Louis) were dissolved in 95% ethanol as stock solution and then diluted with distilled water desired concentration with approximately 0.5% ethanol. Seedlings were assigned into 5 groups, MJ1, MJ2, SA1, SA2 and CK, and sprayed per seedling with 100 ml of 0.5 mmol·L-1 MeJA, 1 mmol·L-1 MeJA, 5 mmol·L-1 SA, 50 mmol·L-1 SA or 0.5% ethanol aqueous solution as control, respectively, under the same conditions. The pots were placed in a sealed plastic greenhouse and cultivated for 2 d until MeJA or SA were absorbed and then transferred to outdoor. Samples were collected at 0 h, 6 h, 12 h, 1 d, 2 d, 3 d, 5 d, 7 d, 14 d and 28 d of post treatment. Each treatment was repeated with 3 plants and total triterpenoid content in stem bark and leaves were detected.
Determination of photosynthetic indexes
The photosynthetic gas exchange parameters were measured with a Li-6400 Portable Photosynthetic System (Li-6400, Li-Cor, Lincoln, NE, USA) at different time of post MeJA and SA treatment. Net photosynthetic rate (Pn), transpiration rate (Tr), and stomatal conductance (Gs) of built-in leaf area of 1.5 cm2 were measured at conditions of CO2 concentration of 500 μmol·s−1 (controlled using a built-in Li-Cor 6400 CO2 controller), and irradiance of 1000 μmol m−2s−1 (provided by a built-in red LED light source) between 9:30am and 11:30am on cloud-free days. Ten leaves in middle of each of the 3 repeats were measured and detected photosynthesis index of, calculating the average value.
Determination of chlorophyll content
Chlorophyll a and chlorophyll b were determined spectrophotometrically according to the methods reported previously [28]. The leaves were extracted with 85% (v/v) acetone aqueous solution and centrifuged at 4000 rpm for 10 min to collect the supernatants. After suitable dilution, its absorption at 452.5nm, 644nm and 663 nm was measured using a Beckman Coulter DU® 520 UV-vis spectrophotometer (USA) and the contents of chlorophyll a and chlorophyll b were measured as described previously [28].
Enzyme activity assay
The birch leaves (0.50 g) were homogenized in 5 ml of phosphate buffer (50 mM, pH 7.8) containing 2% PVPP and 0.2 mM EDTA. The extracts were then centrifuged at 10,000 rpm for 15 min at 4ºC, and the supernatant was used to determine the activity of super-oxide dismutase (SOD) and peroxidase (POD) according to the methods of Lu et al. [29] and the activity of catalase (CAT) as reported previously by Hong et al. [30].
Malondialdehyde (MDA) content assay
The content of MDA was determined using thiobarbituric acid (TBA) assay [31] with slight modification. Leaf samples (0.50 g) were homogenized in 5.0 ml of 5% (w/v) trichloroacetic acid (TCA) and centrifuged at 8000 rpm for 30 min. The supernatant was mixed with 2.0 ml of 0.67% TBA, heated at 100ºC for 30 min and then quickly cooled down on ice. After centrifugation at 3000 rpm for 15 min, absorbance of the supernatant was measured at 450, 532 and 600 nm, respectively.
Contents of soluble sugar and soluble protein assay
Soluble carbohydrate content was measured using throne sulfuric acid reagent as previously reported [32]. Soluble protein content was determined using Bradford method [33].
Determination triterpenoid content
Triterpenoid contents were measured using the method described by Li [34] with slight modification. In brief, dry samples (0.050 g) were homogenized and soaked in 5.0 ml of 95% (V/V) alcohol for 24h. After sonicated at 70 kHz for 40 min, samples were incubated at 70ºC for 1 h in a water bath and then at room temperature for 10 min to extract triterpenoid. A total of 100 μL extracts were dried by evaporating at 70ºC and re-dissolved in 200 μL of 5% vanillin-acetic acid solution. After mixed with 800 μL of perchloric acid and incubated at 70ºC for 15 min, the solution was cooled to room temperature in a water bath and diluted with ethyl acetate to 5 mL. The content of triterpenoid was measured as absorption value at 551 nm. Reagents without sample was used as blank.
Rreal-time quantitative PCR (qPCR)
Total RNA was extracted using CTAB method as described by Chang et al. [35]. After treated with RNase-free DNase I (TaKaRa, Japan), 500 ng RNA was reverse transcribed into cDNA using TaKaRa PrimeScript™ 1st Strand cDNA Synthesis kit, and expression of genes FPS [36], BPX, BPX2, BPY and BPW was measured using qPCR Master Mix-Plus-kit (QPK-212T, TOYOBO) as described previously [37] using housekeeping ß-tubulin (Tu) and ubiquitin (UbQ) genes as the internal controls. Data analysis was performed as described previously [36].
The statistical analysis
The DPS7.50 software and Duncan’s method for different between treatment significant variance analysis.
The effect of MeJA and SA treatment on photosynthetic characteristics of birch
The net photosynthetic rate (Pn) refers to the rate of photosynthesis to produce sugars and could directly reflect the ability of matter production per unit leaf area. In theory, it can be used as a reliable index to measure the level of plant biomass production. The stomatal conductance (Gs) reflects stomatal opening degree and could directly affect plant photosynthesis, respiration and transpiration. The transpiration rate (Tr) refers to the amount of H2O lost during transpiration per leaf area unit in a certain period and is an important index for quantification of transpiration.
Leaf Pn increased gradually with time of posttreatment, except that at 28 d of post-treatment in MJ1 group. In addition, leaf Pn was higher in all treatment groups at 1 d, 7 d and 14 d of post-treatment with MeJA than that of the control group (CK) in the order of MJ1 >MJ2 >CK. Leaf Gs shows a trend of first decrease, then increase, and decrease again. At 14 d of post MeJA treatment, Gs in the treatment group were higher than that of the control. Tr showed a trend of first decrease, then increase, and decrease again, similar to that of Gs in MeJA treatment group. Overall, the results indicate that MeJA treatment could significantly impact Pn, Gs and Tr in birch leaf, and Pn, Gs and Tr in the 14d were higher than of the control, and the low concentration was have more advantage (Figure 1).
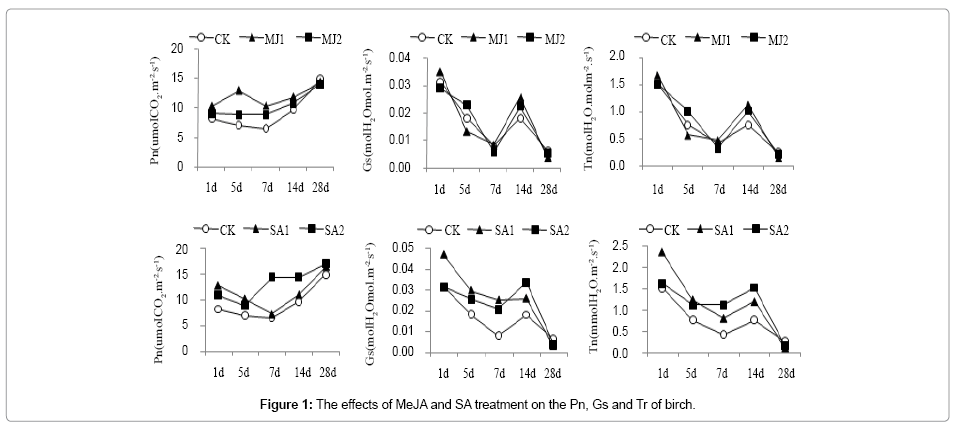
Figure 1: The effects of MeJA and SA treatment on the Pn, Gs and Tr of birch.
Leaf Pn increased with different SA treatment, that was 2.19-fold of control in SA2 group at 7 d of post treatment. Gs and Tr showed similar trends of first increase then decrease and increase again with treatment time, reaching the highest at the beginning of SA treatment in SA1 group, But Gs and Tr reached the highest at 7 d and 14 d of SA treatment, show that SA treatment was conducive to increase of leaf transpiration (Figure 1).
The effects of MeJA and SA treatment on chlorophyll content in birch leaves
High concentration MeJA treatment increased the content of chlorophyll a and b in birch leaf (Figure 2) at late stages in a dose dependent manner. The amount of chlorophyll a and b reached their maximum of 1.66 mg/g and 0.6 mg/g at 28 d of post-MeJA treatment, which was 1.8-fold and 2-fold of the control. Similarly, SA treatment significantly increased the amount of chlorophyll a and b in birch leaf at 14d and 28 d of post-treatment. In addition, SA treatment significantly decreased the ratio of chlorophyll a to chlorophyll b compared with that of the control and MeJA treatment only decreased the ratio of chlorophyll a to chlorophyll b at 5d and 28d of post treatment.
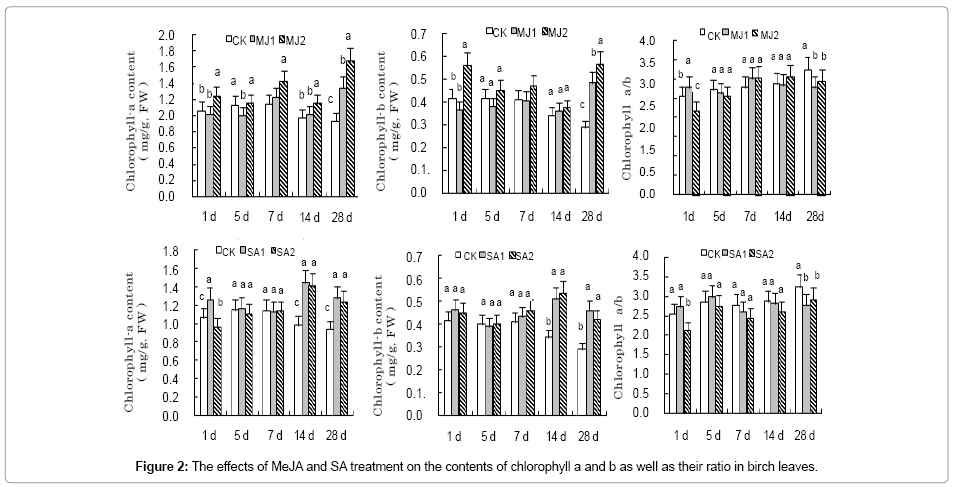
Figure 2: The effects of MeJA and SA treatment on the contents of chlorophyll a and b as well as their ratio in birch leaves.
The effects of MeJA and SA treatment on the antioxidant enzymes and lipid of birch
SOD activity showed a trend of first increase, then decrease, and increase again with time. Compared with the control, SOD activity was higher at 12 h and 5 d of post MeJA treatment but lower at 1~2d and 14- 28d of post MeJA treatment. POD activity was significantly enhanced at 2d and 7d of post MeJA treatment at higher concentration compared with that of the control, reaching maximum fold of 2.76 at 2 d of post treatment, but decreased at other time points. CAT activity was enhanced at 6-12 h of post MeJA treatment, but decreased at 1-5 days of post treatment and then increased again at 14-28 d of post treatment compared with control (Figure 3).

Figure 3: The effects of MeJA and SA treatment on activity of SOD, POD and CAT in birch.
SOD activity showed a trend of first decreased, then increase, decrease again and increase again with time. At 1~3d of post SA treatment, SOD activity was lower than that of CK, shown as CK>SA2>SA1. At 14-28 d of post treatment, SOD activity increased in SA2 group, but decrease in SA1 group compared with that of CK. POD activity did not change at 6-12 h of post SA treatment, but increased significantly at 2-5 d of post SA2 treatment, then decreased at 7 d of post SA treatment and increased again at 14-28d of post SA treatment CAT activity showed a trend of first increase then decrease with time and was in the order of SA2>SA1>CK at the 28d of post treatment. Although SA treatment could affect the activity of these three protective enzymes, their trends differed significantly: POD activity reached the highest at 2-7 d of post treatment with SA at high concentration, while CAT activity was significantly enhanced at 14~28 d of post SA treatment. Visibly, MeJA and SA treatment altered the activity of all the three protective enzymes and theses changes may be coordinated to each other and related to cell membrane protection in birch leaves and accumulated tolerance to stresses (Figure 3).
The MDA accumulation showed two peaks at 1 d and 14 d of post treatment with MeJA and SA indicating that oxidation of membrane lipids by reactive oxygen species occurred at the early and middle stages of post treatment with MeJA and SA. Especially, MDA content at 1 d of post SA treatment reached 2-fold of that of control (Figure 4). We speculate that MeJA and SA treatment could result cell membrane damage in leaf at early stage, at which the activity of protective enzymes was not high enough to eliminate clean reactive oxygen species, thus leading to rapid MDA accumulation. After that, with the increase of protective enzymes activity, MDA accumulation decreased.
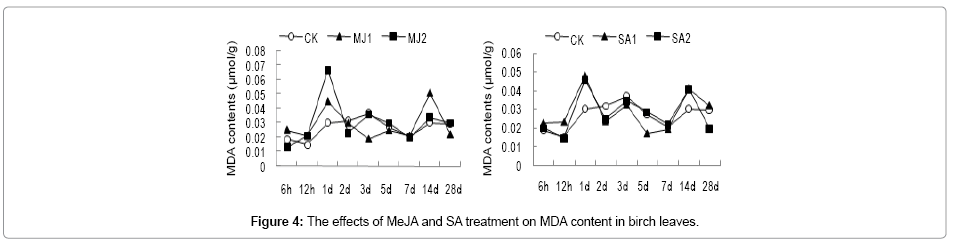
Figure 4: The effects of MeJA and SA treatment on MDA content in birch leaves.
Effects of MeJA and SA on accumulation of soluble sugar and soluble protein content in birch saplings
Soluble sugar content showed a trend of first decrease then slowly increases and decrease again after MeJA treatment. Soluble sugar content was decreased significantly (MJ2
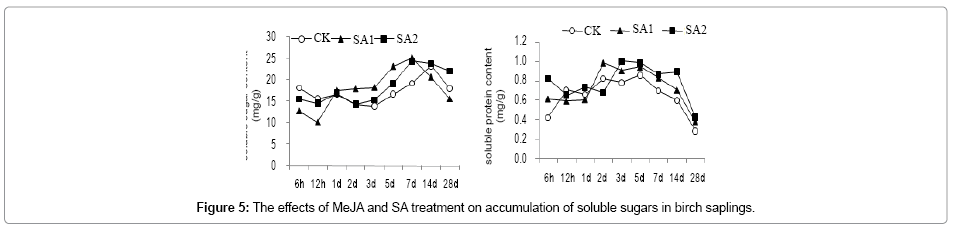
Figure 5: The effects of MeJA and SA treatment on accumulation of soluble sugars in birch saplings.
The soluble protein content was higher than that of the CK at 6h, 5d, 7d and 28d of post MeJA treatment, reaching maximum of 1.16 mg/g (which was 1.35-fold of the control) at 5 d of post MJ1 and after 3d of post SA treatment, reaching its peak at 5d of post SA treatment with the order of SA2>SA1>CK (Figure 6).
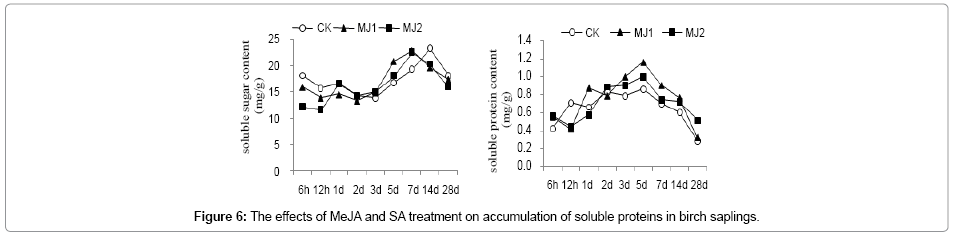
Figure 6: The effects of MeJA and SA treatment on accumulation of soluble proteins in birch saplings.
Effects of MeJA and SA on expression of key genes of triterpene synthesis pathway in birch saplings
Quantitative PCR results showed that key genes FPS, BPX, BPX2, BPY and BPW of triterpene synthesis pathway responded differently to MeJA and SA treatments, and showed different trend in birch leaves and bark. FPS in leaf and FPS, BPX2, BPY and BPW in bark were upregulated at 1d of post MeJA treatment, which was more obvious in at 1~7d of MJ2 group compared with that of MJ1 and CK group. In birch leaf, expression of BPX, BPX2, BPY and BPW was upregulated at 12 h of post MeJA treatment, but decreased at 7d and 14d of post MeJA treatment (Figure 7). Meanwhile, expression of BPX in stem bark was significantly decreased at 7d~14d of post MeJA treatment (Figure 8).
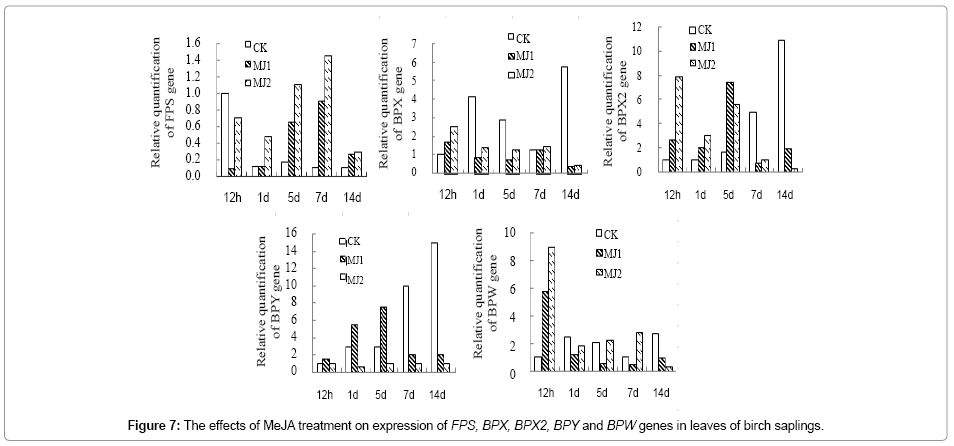
Figure 7: The effects of MeJA treatment on expression of FPS, BPX, BPX2, BPY and BPW genes in leaves of birch saplings.

Figure 8: The effects of MeJA treatment on expression of FPS, BPX, BPX2, BPY and BPW genes in bark of of birch saplings.
Expression of BPX, BPX2 and BPY gene in birch leaves was upregulated at 12 h of post SA treatment, but expression of BPW was decreased at all-time points of post SA treatment (Figure 9). Expression of FPS, BPX2, BPY and BPW in birch bark was significantly upregulated at 7d and 14d of post SA treatment, but expression of BPX was inhibited significantly at 1d~14d of post SA treatment (Figure 10).
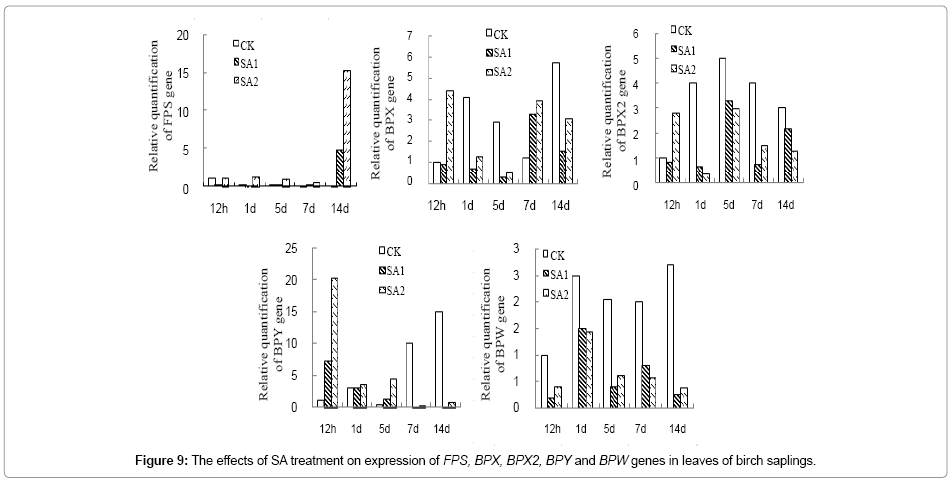
Figure 9: The effects of SA treatment on expression of FPS, BPX, BPX2, BPY and BPW genes in leaves of birch saplings.
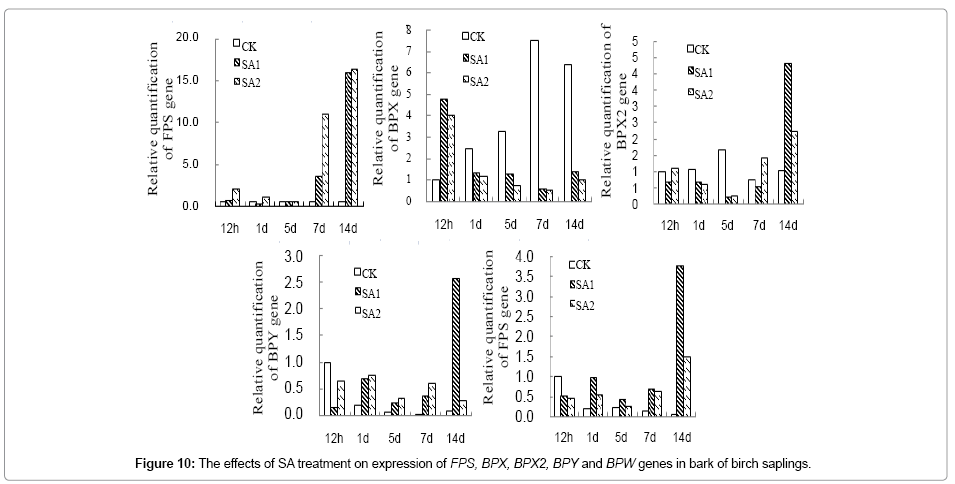
Figure 10: The effects of SA treatment on expression of FPS, BPX, BPX2, BPY and BPW genes in bark of birch saplings.
Effects of MeJA and SA treatment on triterpenoid accumulation in white birch
Triterpenoid content in birch leaves was enhanced by 31.50% at 5 d of post MeJA treatment, reaching the peak of 107.27 mg/g (Figure 11A), and still higher than the control at 14d and 28d of post MeJA treatment with the order of MJ2>MJ1>CK. At 28 d of post treatment with MeJA at high concentration, triterpenoid content was up to 101.57 mg/g, increased by 53.2% compared with that of the control. Similarly, triterpenoid content in birch leaves was enhanced at 1 d and 5 d of post treatment with high concentration of SA, although at lower degree than that of MeJA treatment (Figure 11C).
Figure 11: the effects of MeJA and SA treatment on triterpenoid accumulation in birch leaves and bark.
Triterpene content in birch bark was increased at 1 d and 5 d of post MeJA treatment, reaching 81.86 mg/g at 1 d of post treatment with high MeJA concentration, which was 46.11% more than that of the control. In addition, triterpene content in birch bark also increased at 28 d of post MeJA treatment (Figure 11B). Furthermore, triterpene accumulation of stem bark was enhanced at 1d, 5 d and 14 d of post SA treatment, reaching its peak of 91.4mg/g at 14 d of post treatment with SA at lower concentration, which was 45.07% more than that of the control (Figure 11D).
The above results showed that low concentration of MeJA treatment was favor to triterpene accumulation in birch leaves at early days of post treatment, while high concentrations of MeJA treatment was more conducive to triterpene accumulation in birch bark and low concentration of SA treatment was favor to triterpene accumulation in birch bark at early days of post treatment while high concentration of SA could significant promote triterpene accumulation in birch leaves at early days of post treatment.
Photosynthesis is the key of plant material transformation and energy metabolism. Pn, Gs and Tr were important indexes of photosynthesis reactions of plants. Reportedly, MeJA could inhibit photosynthetic function. Treatment of seedlings with MeJA alone resulted in decreased levels of Chl, photosynthesis and Tr. Pre-treatment of seedlings with MeJA fully blocked the inhibitory effect of paraquat (Pq) on photosynthesis and provided protection against subsequent Pq induced oxidative damage [38]. Ritsema et al demonstrated that MeJA and SA mediate differential phosphorylation of substrates for many kinase families of Arabidopsis thaliana and some plant specific substrates including peptides derived from phytochrome A, and photosystem II D protein. MeJA and SA mediate cross-talks between defense signaling and light responses and act in a synergistic fashion via two (partially) divergent signaling routes [39]. Bilgin showed that though MeJA treatment altered the transcript levels of various defence and ROS scavenging network genes, it did not have a strong effect on photosynthesis-related genes, suggesting that the involvement of MeJA might be more active and direct in defence signaling than photosynthesis-related reactions and ROS scavenging network [40]. Similar to ABA, MeJA induced stomatal closure is an important physiological process responding to cytoplasmic alkalization, ROS and NO in plants [41].
Antioxidants enzymes in plants and their coordination can effectively remove free radicals such as OH- and H2O2, and prevent membrane peroxidation, so as to protect cell membrane damage. SOD can remove O2-, and convert it into low active H2O2, which further reduced to H2O by POD and CAT. MeJA and SA can improve activity of these protective enzymes in various plants, therefore clearing ROS, protecting cell membrane and improving resistance of plants to diseases, drought and cold [42-45]. Involvement of calmodulin for transmission of defense signal by SA has been suggested. A metabolic reprogramming leading to enhanced synthesis of proteins involved in primary and secondary metabolisms is necessary for SA mediated resistance to MYMIV. Enhanced expression of proteins involved in photosynthetic process could restore virus-induced degradation of the photosynthetic apparatus and provides more metabolites required for repartition of resources towards defense [43]. Our results show that MeJA and SA treatment significantly affected Gs Pn and Tr in birch leaves. Treatment with MeJA at low concentration significantly increased Pn, but inhibited Gs and Tr (Figure 1). Treatment with SA at different concentrations increased Pn. Treatment with SA at ow concentration enhanced Gs and Tr at the early stage, while at high concentration had little effect compared with the control; Treatment with SA at high concentration increased Gs and Tr at 7d and 14d of post treatment. Treatment with SA decreased Gs and Tr at 28 d of post treatment (Figure 2). MeJA and SA treatment could obviously increase the contents of chlorophyll a and b at 14d and 28d of post treatment (Figure 3 and 4). In addition, MeJA and SA treatment significantly altered SOD, POD and CAT activity in birch leaves, showing different degree with different concentration at different time (Figure 5 and 6). For example, MeJA treatment significantly enhanced POD activity at 12 h of post treatment, SOD activity at 2d and 7d of post treatment, and CAT activity at 14d and 28d of post treatment, compared with control. The results suggested that MeJA and SA could initiate birch defense system, and coordinate with SOD, POD and CAT to improve its stress resistance, regulate its photosynthesis and affect its secondary metabolite accumulation.
As the main infiltration regulator of many plants and energy sources of carbon frame for synthesizing other organic solutes, they also could stabilize cell membrane and protoplasmic colloid and protect intracellular protective enzymes at high concentration of inorganic ions [44,45]. Suárez-González reported that SA, MeJA and other elicitors could increase fructo-oligosaccharide accumulation concomitant with enhanced expression of fructan biosynthetic genes in Agave [43]. Changes in proteins expression can partly represent primary metabolism level of plants. In addition, stresses such as low temperature and drought could affect soluble protein content. Plants produce some special secondary metabolites through a series of enzyme catalysis using primary metabolites as raw materials. Most of the soluble proteins in plants are enzymes involved in various metabolic pathways and their contents may be related to the amount of the secondary metabolites of plants [44]. In this study, the level of soluble sugars in birch leaves was higher than that of the control after MeJA treatment and at 3~7d of post SA treatment (Figure 8), which laid the foundation for regulating cell permeability and provided carbon source for synthesis framework of secondary metabolites. At 6h, 5d, 7d and 28d of post MeJA or SA treatment, soluble protein contents were higher than that of the control, especially at 5 d of post treatment with MeJA at low concentration and with SA at high concentration (Figure 9). Overall, MeJA and SA treatment could influence the primary metabolism in birch leaves an important feature of the terpenoid secondary metabolism is tissue-specific and closely related to the location of the enzymes key their production [46-51]. Birch terpenoids are mainly produced in the cytosol using acetyl CoA as the initial precursor through MVA pathway synthesis. The synthesis process of terpenoids in birch consists of a series of enzymes (HMGR, GPS, FPS, OSCs) and their expression characteristic could affect the synthesis of terpenoids and their precursors. Research has shown that MeJA and SA treatment could affect a variety of secondary metabolites such as paclitaxel, camptothecin, isoflavone and saponin in cultured cells and induce the expression of the key enzymes for their synthesis. Meanwhile, MeJA and SA treatment could promote the synthesis of monoterpenes and sesquiterpenes in spruce, pine, coconut, cherry tomatoes, and Citrus unshiu Marcprolong at the antistaling period [52-56]. MeJA and SA have a positive role in promoting the accumulation of squalene and LUS, the precursors of terpenoids biosynthesis and inducing β-AS mRNA expression and soybean saponin synthetase in cultured cells of licorice (Glycyrrhiza glabra) [48]. Overexpression of genes involved in triterpene biosynthesis may be expected to increase the total amount of withanolides in W.somnifera [55]. Our study examined the expression of genes key to the terpenoids synthesis in birch and found that treatment of MeJA and SA at different concentration has different impacts on the expression of FPS, BPX, BPX2, BPY and BPW genes in birch bark and leaves (Figure 10 and 13 At 1 d of post MeJA treatment, the expression of FPS in birch leaves and FPS, BPX2, BPY and BPW in birch bark started to increase, and this increase was more obvious at 1~7d of post treatment with MeJA at high concentration that with low concentration. This is possibly the cause for rapid accumulation of terpenoids in bark after treatment with MeJA at high concentration (Figure 11B). BPX, BPX2, BPY and BPW genes were up-regulated in birch leaves at 12 h of post MeJA treatment, but downregulated at 7d and 14d of post MeJA treatment, which agrees with the general accumulation trend of terpenoids in leaves. In addition, BPX expression in birch bark was significantly inhibited at 7d~14d of post MeJA treatment (Figure 11), which may determine the dynamic changes of terpenoids accumulation in birch bark and leaves.
The above analyses showed that the synthesis and accumulation of terpenoids in birch induced by MeJA and SA not only depend on the expression of genes key to terpenoids biosynthesis but also are closely related to the physiological levels of photosynthesis, anti-stress enzymes, soluble proteins and soluble sugars, suggesting that synthesis of secondary metabolic products in birch and metabolism network are very complex. Previous researches have shown that distribution of terpenoids components in birch is obviously tissue specific, with high betulin content in the bark and abundant Marana in the leaves [37,56]. In this study, we only examined the total triterpenoid contents in birch. The distribution of its components such as betulin, oleanolic acid, cholesterol and others in birch bark and leaves as well as their relationship to transport, metabolism and synthesis pattern of genes BPY, BPW and BPX need to be further explored and discussed in depth.
We thank Peng Hong-Mei and Yang Yuan-Biao of Maoershan Experimental Forestry Center for greenhouse management of Northeast Forestry University, Harbin, China. This work was supported by Natural Science Foundation of Heilongjiang Province(C201110), Natural Science Foundation of China (31200428, 31070531), and Youth Innovate Foundation Grant of Haerbin city(2012RFQXN007).