Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Review Article - (2013) Volume 0, Issue 0
Pancreatic cancer is a leading cause of cancer death and most patients have advanced disease at the time of diagnosis, making early detection of paramount importance. Intraductal papillary mucinous neoplasm (IPMN) is a cystic precursor lesion to pancreatic cancer that is being diagnosed with increasing frequency often in asymptomatic patients. While some IPMNs may progress to invasive carcinoma and thus, require resection, others will remain insignificant and may be followed without surgical intravenation. Unfortunately, our understanding of the natural history of IPMN is limited, and we are not yet able to confidently segregate high-risk and malignant lesions from low-risk ones. Molecular profiling that allows us to gain insight into the biology of the disease may provide us with clues to improve preoperative decision making. In this review, we will discuss the most recent findings in the molecular alterations of IPMN including those in tumor suppressor genes and oncogenes, chromosomal copy number abnormalities, epigenetic alterations, and miRNAs, followed by the implementation of those markers in the analyses of cyst fluid that can be obtained via endoscopic ultrasound guided fine needle aspiration. We will also touch upon pancreatic duct glands (PDG), a possible cell origin of IPMN.
<Keywords: Pancreatic cancer, Endoscopic ultrasound guided fine needle aspiration
Pancreatic cancer is the fourth leading cause of cancer death in the United States. In 2012, the American Cancer Society estimated that 43,920 new cases would be diagnosed in that year, and 37,390 patients would die of the disease [1]. Despite significant progress in our understanding of pancreatic cancer and decreased mortality and morbidity secondary to pancreatic surgery, patients with pancreatic cancer still face dismal outcomes. For all stages combined, the 1- and 5-year relative survival rates are 26% and 6%, respectively. At the time of diagnosis, less than 20% of patients are surgical candidates. Even for those with local disease that undergo surgery, the 5-year survival rate is only 22% due to disease recurrence or hidden metastases. More than half of patients are diagnosed at an advanced stage, with a 5-year survival of only 2% [1].
Early detection is believed to offer the best opportunity to eventually reduce the human suffering caused by pancreatic cancer. Thus, it is important to improve our understanding of its precursors. Three histologically defined precursors to invasive adenocarcinoma of the pancreas (PDAC) have been described: microscopic, non-cystic, pancreatic intraepithelial neoplasia (PanIN), and cystic, mucinous cystic neoplasm (MCN) and intraductal papillary mucinous neoplasm (IPMN). Pancreatic intraepithelial neoplasia (PanIN) is a microscopic papillary or flat, non-invasive epithelial neoplasm that exhibits a gastric epithelial morphology and usually involves intralobular and/or small interlobular pancreatic ducts [2]. PanIN exhibits different degrees of cytological and architectural atypia and are classified into three grades accordingly, from low-grade to high-grade dysplasia. Most PanIN lesions are 0.5-cm or smaller, and thus, they are not diagnosed clinically, although they may be found in pancreata resected not only for PDAC but also for other entities including unbearable chronic pancreatitis [2]. MCNs are mucin-producing premalignant lesions found almost exclusively in the distal pancreas of middle-aged women and can be detected with cross-sectional imaging. We recently reported that MCN now accounts for only 16% of resected cystic tumors of the pancreas at our institution [3]. MCN is an aggressive entity once it becomes invasive: the 5 year disease-specific survival for non-invasive MCN is 100%, whereas that of invasive MCN is only 58% [4,5]. Given their epidemiological target group and premalignant nature, MCNs are usually resected regardless of the presence of stigmata of malignancy. IPMNs are the third group of precursor lesions and most are 1-cm or larger, and thus, easily detectable radiographically [2]. IPMN was first described as a rare entity by Ohashi et al. in 1982 [6], but it has been increasingly diagnosed. In fact, IPMN accounted for only 2% of resected cystic tumors in our hospital before 1991, but today (over the past 7 years), 49% of resected cystic tumors of the pancreas are IPMNs. Similarly, a recent study of 2,832 consecutive abdominal CT scans undertaken for unrelated issues has found a prevalence of asymptomatic pancreatic cysts (the vast majority of which are considered to be IPMN) to be 2.6% among all comers and 8.7% among those above the age of 80 [7]. The presence of invasive components in IPMN also dictates worse outcomes. While the 5-year survival of patients with resected noninvasive IPMN is as high as 77-94%, invasive IPMN carries a much poorer survival of 33-43% [8-12]. As described in the next section, IPMN is classified into main-duct, combined and branch-duct types based on the differential involvement of the pancreatic duct system. We have shown that main-duct and combined type IPMNs are more likely to have invasive carcinoma compared to branch-duct type (48% and 42% vs. 11%), and subsequently, the 5-year disease specific survival rates of main-duct and combined type IPMNs are significantly lower than that of branch-duct type (65% and 77% vs. 91%) [4].
Given the significant difference in survival between invasive and non-invasive IPMNs as well as between main-duct and branch-duct IPMNs, clinical guidelines have been adopted to assist clinicians in determining when a lesion should be surgically resected [13]. However, while sensitive (97-100%), these guidelines have proven to be highly non-specific (23-30%), especially among branch-duct IPMN [14-16]. The guidelines have recently been modified to improve the specificity [17], but their performance is yet unknown. Given the prevalence of asymptomatic cysts especially in an elderly population who tend to have comorbidity, accurate and reliable diagnostic methodologies are needed to identify which cystic lesions carry a high risk for malignant transformation and need to be resected, and which may be watched safely. Unveiling the molecular phenomena underlying the development of IPMN will help us improve our understanding of these lesions and will shed new light on how to approach these tumors.
In this review, we will discuss the overview of IPMN followed by the most recent findings in molecular alterations of IPMN, from the most common mutations shared with PDAC, to the more specific epigenetic and miRNA alterations. Further, we will discuss the role of cyst fluid analysis in the preoperative diagnosis of IPMN and how the molecular findings are being incorporated into new evidence-based diagnostic approaches. Lastly, we will discuss the recently described pancreatic duct glands (PDG), a distinct compartment that appears to be the origin of IPMN.
IPMN is a tumor characterized by intraductal proliferation of neoplastic mucinous cells with various degrees of cytologic atypia, which usually form papillae and lead to cystic dilatation of pancreatic ducts. IPMN is most common in patients in the seventh decade of life. Previously thought to be a disease of the male gender, it has been observed that gender distribution of IPMN varies in different geographic areas [18]. Currently, the majority of patients diagnosed with IPMNs are asymptomatic and the lesions are incidentally discovered during the evaluation of unrelated abdominal or thoracic conditions [19]. When associated with symptoms, patients present with abdominal pain, weight loss, diabetes, jaundice and acute pancreatitis [4]. Macroscopically, IPMN is classified into the branch-duct (BDIPMN), main-duct (MD-IPMN) or combined type based on the extent of pancreatic duct involvement by neoplastic cells. Microscopically, IPMN exhibits variable degrees of cytological and architectural atypia that are classified into low-grade dysplasia (formerly adenoma), intermediate-grade dysplasia (formerly border-line), high-grade dysplasia (formerly in situ carcinoma) and invasive carcinoma [2,20].
Histologically, IPMN demonstrates significant heterogeneity in their epithelial morphology. Four histological subtypes have been described: gastric, intestinal, oncocytic and pancreatobiliary (Figure 1) [21]. Important differences exist between these subtypes; not only they are morphologically distinct, but also they exhibit different mucin glycoproteins and different molecular alterations, and more importantly, they correlate with the histological subtypes of invasive carcinoma (Figure 2) and ultimately with prognosis [20,21]. Gastrictype and pancreatobiliary-type IPMNs usually give rise to invasive tubular adenocarcinomas, morphologically similar to conventional pancreatic adenocarcinoma. On the other hand, most of intestinal-type IPMNs give rise to colloid carcinomas, while some develop into tubular adenocarcinomas. Finally, invasive carcinoma arising from oncocytic type IPMN mostly retains its oncocytic features. Collectively, patients with invasive IPMN have a threefold better 5-year survival than those with conventional PDAC (not associated with IPMN). However, we recently showed that this strong advantage is due to the indolent tumor biology of colloid and oncocytic carcinomas, while the tubular type has a prognosis similar to that of conventional PDAC [20].
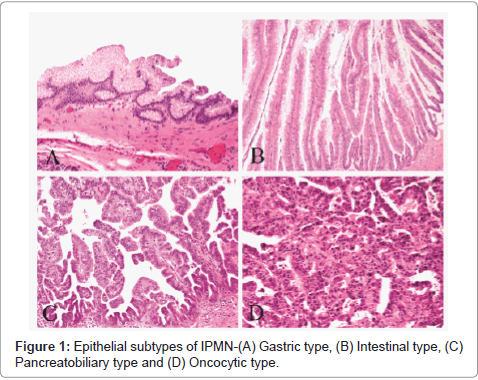
Figure 1: Epithelial subtypes of IPMN-(A) Gastric type, (B) Intestinal type, (C) Pancreatobiliary type and (D) Oncocytic type.
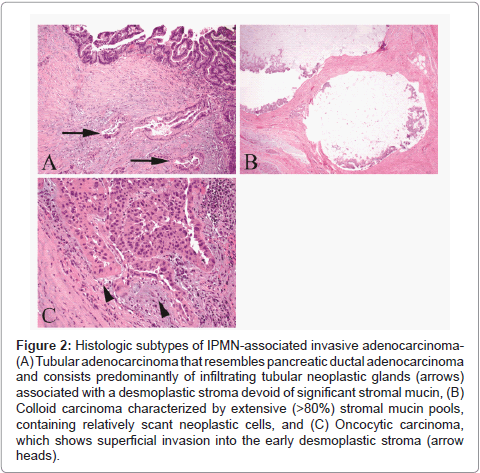
Figure 2: Histologic subtypes of IPMN-associated invasive adenocarcinoma- (A) Tubular adenocarcinoma that resembles pancreatic ductal adenocarcinoma and consists predominantly of infiltrating tubular neoplastic glands (arrows) associated with a desmoplastic stroma devoid of significant stromal mucin, (B) Colloid carcinoma characterized by extensive (>80%) stromal mucin pools, containing relatively scant neoplastic cells, and (C) Oncocytic carcinoma, which shows superficial invasion into the early desmoplastic stroma (arrow heads).
A wealth of molecular alterations has been identified in IPMN. Some of these alterations are shared by PDAC–to various extents– whereas others are almost exclusively found in IPMN. Furthermore, recent technological advancements have made it possible to identify changes at the epigenetic level and to characterize miRNA expression in both tissue and cyst fluid. In this section, we will discuss the most prominent molecular findings in IPMN and their potential implications in tumor progression.
KRAS (Kirsten rat sarcoma viral oncogene homolog) belongs to a family of GTP-binding proteins involved in a host of cellular functions including proliferation, differentiation, motility and survival mediated by the mitogen-activated protein kinase (MAPK) pathway; it has been one of the most actively studied proteins/genes in both PDAC and IPMN [22,23]. Mutations in KRAS, particularly those in codons 12 and 13, can be found in over 90% of PDAC [23,24]. Oncogenic KRAS mutations have now been recognized as one of the earliest events in pancreatic carcinogenesis, observed in more than 90% of PanIN lesions including low-grade PanIN [25]. The prevalence of KRAS mutations in IPMN has been reported by several groups, often with discrepant results that range from 38.5% to 100% [26-28]. Indeed, most studies agree that the frequency of KRAS mutations in IPMN is lower than that of PDAC. The lower mutation prevalence reported in some series raises the possibility of additional genes besides KRAS contributing to cell proliferation in IPMN [29,30]. Nissim et al. conducted a meta-analysis evaluating 13 studies representing a total of 285 IPMN specimens and found that oncogenic mutations in KRAS can be detected in 48% of adenoma/borderline (low/intermediate-grade) IPMN, and 55.1% of IPMN carcinoma (high-grade) [31]. Some studies have shown that IPMN demonstrate a significant increasing rate of KRAS mutations as the degree of dysplasia progresses [32-34]. However, we and others recently found that activating KRAS point mutations occurred in 66% and 87% of IPMN with low-grade dysplasia without significant difference compared with the remaining higher-grade IPMN. These results suggest that, similar to PanIN, KRAS mutations in IPMN occur at an early stage of neoplastic transformation [28,29]. The characterization of 4 epithelial subtypes in IPMN associated with different profiles of mucin expression and variable prognosis has raised the possibility of different molecular pathways in their progression. In a recent study, we have found that gastric type IPMN demonstrate a higher rate of KRAS mutations than intestinal and oncocytic types [29]. This observation is supported by two recent studies. Mohri et al. [35] revealed KRAS mutations in 82% of gastric-type IPMN compared to 27% of the intestinal type (and 56% overall prevalence) as well as a significant correlation between KRAS mutations and the nuclear expression of p-ERK in the gastric-type lesions, and Wu et al. [28] found KRAS mutations in 87% of the gastric type (and 100% of the pancreatobiliary type) opposed to 47% of the intestinal type. Remarkably, KRAS mutations were detected in gastric-type IPMN with low grade-dysplasia in both studies [28,35]. This data suggests a significant similarity between gastric-type IPMN and PanIN, as both of them seem to share activating KRAS mutations as an early event in addition to the gastric morphology. Despite the observed differences between non-invasive IPMN histological subtypes, activating KRAS mutation rates appear to be similar between tubular and colloid carcinomas (approximately 80% in both types) [20]. As described above, colloid carcinoma arising in the background of intestinal type IPMN has a better prognosis than tubular carcinoma; thus, KRAS mutations appear not to predict tumor aggressiveness in invasive IPMN [20] (Table 1).
| Gene | Chromosome | Mechanism of alteration | Prevalence in IPMN6 | Prevalence in PDAC7 | |
| Oncogenes | KRAS | 12p | Point mutation in codon 12 | 38-100% | >90% |
| Tumor suppressor genes | p53/TP53 | 17p | LOH1+IM2 | 0-50% | 50-80% |
| SMAD4/DPC4 | 18q | LOH+IM, HD3 | <3% | ~55% | |
| p16/CDNK2A | 9p | LOH+IM, HD, Meth4 | 0-100% | >95% | |
| Chromosomal abnormalities | 5q | CNA5 - Loss | 53.8%* | 18.9% | |
| 6q | CNA - Loss | 75-100%* | 33.6% | ||
| 11q | CNA - Loss | 61.5%* | 5.5% |
Table 1: Molecular alterations shared by IPMN and PDAC.
SMAD4/DPC4: DPC4 (Deleted in Pancreatic Carcinoma, locus 4) is a tumor suppressor gene involved in the TGF-beta signaling cascade, where it mediates TGF-beta induced growth inhibition via its gene product, SMAD4 [36,37]. While inactivation of SMAD4/DPC4 can be detected in the majority of PDACs, inactivation in IPMN is virtually nonexistent in non-invasive lesions and very uncommon in invasive carcinomas [37]. As opposed to KRAS, which is considered to be an early event in the genesis of neoplasms of the pancreatic ducts, loss of DPC4 has been found to be a late event in pancreatic tumorigenesis. In a study by Wilentz et al., all lower-grade PanINs (PanIN-1A, -1B, and -2) revealed preserved expression of DPC4 protein, compared with 69% of PanIn-3 lesions [38]. SMAD4/DPC4 expression is preserved almost universally in non-invasive IPMN, independently of the degree of dysplasia [37,39]. Moreover, loss of SMAD4/DPC4 is a rare event even in invasive IPMN. Iacobuzio-Donahue et al. evaluated 79 IPMNs and found that 97% of invasive carcinomas preserved DPC4/SMAD4 expression; interestingly, the only case of invasive IPMN with loss of DPC/SMAD4 expression was a case of tubular adenocarcinoma [37]. These findings demonstrate that loss of DPC4/SMAD4 expression is a phenomenon much more closely related to advanced PanIN and PDAC than it is to IPMN, suggesting different tumorigenic pathways.
p16/CDKN2A: p16 is a tumor-suppressor protein encoded by the CDKN2A gene in chromosome 9p21 and it is inactivated in over 95% of PDACs [40]. Inactivation of p16 in IPMN has been described by several groups and its prevalence varies widely [41,42]. p16 inactivation appears to correlate with the degree of dysplasia in IPMN, suggesting that this is a late event in tumor transformation. In a study by Biankin et al., 10% of low- and intermediate-grade IPMNs demonstrated inactivation of p16, compared to 100% of high-grade lesions [43]. Nevertheless, p16 inactivation in high-grade IPMN is still less prevalent than in high-grade PanIN [44]. Finally, there is no difference in the expression of p16 between gastric-type and intestinal-type IPMNs [35].
p53/TP53: Gene mutations of TP53 or abnormal p53 protein accumulation can be seen in 50% to 80% of PDACs [45,46]. Alterations in p53 have been well described as a late event in PDAC carcinogenesis, being markedly more frequent in high-grade PanIN (PanIN-3) than in low-grade lesions [44,47]. The overall prevalence of p53 mutations in IPMN varies widely in different series, ranging from 0% to 50%. As it is the case with PanIN, the frequency of p53 mutations in IPMN increases with higher grades of dysplasia [33,35,41]. Nevertheless, p53 mutations in high-grade PanIN are more prevalent than those in high-grade IPMN [44]. No significant difference has been found in p53 mutation rates between gastric-type and intestinal-type lesions, whereas mutations in the pancreatobiliary type are more common than in the oncocytic type (58% vs. 11%) [35,48].
GNAS: GNAS codifies for the alpha subunit of the G protein (Gsα) and it has been found to be mutated in several endocrine neoplasms and less frequently in non-endocrine tumors [49,50]. Until very recently, GNAS had not been recognized as a relevant gene in pancreatic neoplasms. It is now known that a large number of IPMNs carry mutant variants of GNAS and their role in the pathogenesis of pancreatic cystic neoplasms is being extensively studied. Two studies have almost simultaneously revealed that 41% and 66% of IPMNs carry somatic mutations in GNAS [28,51]. All the mutations observed involved codon 201. Remarkably, no mutations were found in conventional PDACs. Furthermore, both studies also looked at KRAS mutation status. Furukawa et al. found that KRAS mutations were present in 49% of the examined IPMNs, whereas 41% harbored GNAS mutations and 25% had concurrent mutations in GNAS and KRAS [51]. On the other hand, Wu et al. found that 81% of IPMN carry KRAS mutations, 51% harbored both mutations, and 96.2% had at least one of the two genes mutated [28]. Furukawa et al. did not find any correlation between mutation status and clinicopathological features. Neither study found a significant correlation between GNAS status and grade of dysplasia. However, Wu et al. found variable rates of GNAS mutations (Figure 3) based on IPMN histological subtype: 100% of intestinal-type IPMNs had GNAS mutations, compared with 42% of the pancreatobiliary type and 65.4% of the gastric type [28]. We recently conducted a pilot study at our institution evaluating GNAS and KRAS mutation status in 17 resected IPMNs with low- to intermediate-grade dysplasia using a multiplex PCR assay (SNapShot platform). We found GNAS mutations in 67% of gastric-type IPMNs and 80% of intestinal-type IPMNs. Further, we found KRAS mutations in 67% of the gastric type and 20% of the intestinal type. Concurrent mutations were observed in 67% and 20%, respectively. These results indicate that gastric type IPMN has a tendency to harbor both GNAS and KRAS mutations even at its early stage [52].
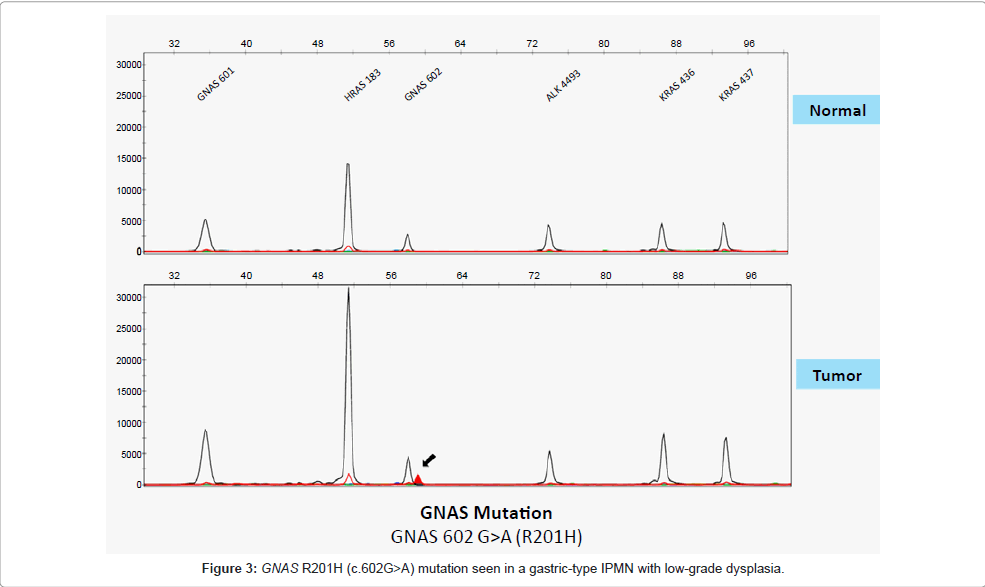
Figure 3: GNAS R201H (c.602G>A) mutation seen in a gastric-type IPMN with low-grade dysplasia.
PIK3CA: PIK3CA mutations are present in a minority of IPMN, with different series reporting from 0% to 11%. Mutations have been found in the entire spectrum of dysplasia, from low-grade IPMN to invasive carcinoma [48,53,54].
BRAF: Few studies have analyzed BRAF mutations in IPMN and the results reveal a low prevalence (2%-12%) scattered across IPMN with low-grade dysplasia to invasive cancer [30,35].
STK11/LKB1 mutations due to allelic inactivation have been reported in IPMN of patients with Peutz-Jeghers syndrome, and it is also present in 25% of sporadic cases [55,56]. Allelic inactivation of the wild-type APC gene has been observed in patients with familial adenomatous polyposis [57], while abnormalities in the Wnt signaling pathway (demonstrated by immunohistochemistry) have been reported in sporadic IPMN [58].
Chromosomal copy number imbalances have been shown to be common in IPMN. The cumulative and increasingly complex patterns of genomic alterations observed in IPMN appear to reflect the neoplastic progression from low-grade dysplasia to high-grade dysplasia and frank invasive carcinoma [29,42]. In a study evaluating 57 IPMNs using microarray-based comparative genomic hybridization (CGH), we found that chromosomal copy number imbalances were present in all IPMNs with at least intermediate-grade dysplasia, and absent in all IPMNs with low-grade dysplasia [29]. The most common chromosomal gains observed were those of chromosome 7 and 19q, seen in 50% of high-grade dysplasia or invasive carcinoma. The most frequent copy number loss was 6q, which has also been reported by Soldini et al. [59] and Abe et al. [29,60]. Interestingly, no definite tumor suppressor gene has been clearly identified in this location. Loss of 18q (possibly driven by loss of SMAD4 tumor suppressor gene, located in this locus) has been recognized as an important alteration in IPMN. Soldini et al. observed that synchronous loss of chromosomes 6, 17, and 18 appears to be associated with high-grade dysplasia or invasive carcinoma [59]. We found loss of 18q in 40% of intermediate-grade IPMN, and in 62% of malignant IPMN (high-grade or invasive). These results suggest that loss of 18q might be a critical step in the progression of IPMN from intermediate-grade dysplasia to high-grade and invasive cancer. These findings deserve our attention, given that SMAD4 is rarely mutated in IPMN, as opposed to PDAC [29]. Abe et al. described loss of heterozygosity (LOH) of 10q as the most common finding in 33 IPMNs, 25 of which had associated invasive carcinoma [60] (Table 1).
Epigenetic alterations have been recognized in IPMN. The vast majority of IPMNs demonstrate promoter methylation in at least one locus. Abnormal methylation can be observed even in IPMN with lowgrade dysplasia, and these anomalies increase with higher grades of dysplasia [61-63]. Abnormal DNA methylation has been found mostly in tumor suppressor genes like p16/CDKN2A, CDKN1C and ppENK. However, a recent study evaluating the genome-wide CpG island profile of IPMN revealed that BNIP3, PTCHD2, SOX17, NXPH1, and EBF3 are the genes most likely to be methylated in IPMN with highgrade dysplasia than lower grades [61]. Among those, BNIP3 proved to be the most specific marker for high-grade dysplasia, not methylated in lower-grade IPMNs or normal pancreatic ducts [61].
microRNAs (miRNAs) are small, noncoding molecules of RNA that bind messenger RNAs (mRNA) disrupting adequate gene expression through degradation of the target mRNA or by inhibition of translation. The detection of aberrant miRNAs expression in various cancers including pancreatic adenocarcinoma has underscored their involvement in carcinogenesis by promoting the expression of protooncogenes or by inhibiting the expression of tumor suppressor genes [64,65]. miRNAs have received an increasing attention as potential markers for malignant progression in IPMN, especially since the miRNA expression profile of the latter has been found to differ from that of PDAC [64]. Habbe et al. performed one of the first studies evaluating miRNA expression in IPMN and analyzed a panel of 12 miRNAs previously known to be up-regulated in conventional PDAC. They found that miR-115 and miR-21 were significantly upregulated in the majority of 15 non-invasive IPMNs and that their expression increases along with increasing grades of dysplasia [39,66]. After this study, several other groups have reported their experience, corroborating the above findings and evaluating new target miRNAs. In these new studies, miR-21 has been found to be up-regulated in noninvasive IPMN compared with normal tissue, and in invasive IPMN compared with non-invasive IPMN, suggesting an important role in early pathogenesis that continues through tumor progression [64,67]. Furthermore, miR-21 is an independent prognostic factor for nodal involvement, mortality and disease progression in invasive IPMN, affecting both overall survival and disease-free survival [67]. miR- 155 is up-regulated in a similar fashion, significantly over-expressed in non-invasive IPMN compared with normal pancreas, and also in invasive IPMN compared with non-invasive tumors [64,67]. miR-101 is another miRNA with an important correlation with IPMN. Similar levels of miR-101 expression were found in normal pancreata and noninvasive IPMN; however, levels in invasive IPMN were significantly lower [67]. Moreover, Nakhara et al. found that loss of miR-101 in IPMN causes upregulation of EZH2, a histone methyltransferase involved in human carcinogenesis, suggesting a role in the progression of IPMN [68]. Several other miRNAs have been identified as potential markers for IPMN progression and grade of dysplasia. Lubezky et al. found 14 miRNAs that were differentially expressed in all IPMN grades compared with normal pancreata, suggesting they might play a role in an early stage of carcinogenesis; further, they found 15 miRNAs with significantly different expression between low-grade/intermediategrade IPMN and malignant lesions, raising the possibility of using them as markers for malignancy [64]. miR-217 stands out as one of the most significantly down-regulated miRNAs between low/intermediate grade IPMN and malignant lesions. Conversely, miR-708 is up-regulated in malignant lesions compared to lower grades. Finally, miR-146a upregulation significantly correlated with IPMN grade; moreover, this miRNA has been previously found to be over-expressed in different grades of PanIN compared with normal pancreatic ducts [64]. Authors agree that more prospective studies are needed in order to identify reliable methods and adequate target miRNAs for clinical application.
The cystic nature of IPMN makes cyst fluid analysis a logic step in the diagnostic investigation of these lesions. Pancreatic cyst fluid examination has been done for several years and it was initially performed intra-operatively through direct needle aspiration of the cyst contents [69]. Today, fluid from pancreatic cysts is obtained through fine-needle aspiration guided by endoscopic ultrasound (EUS-FNA). EUS has become a very important tool in the diagnostic approach of pancreatic tumors, allowing for visualization of the pancreas through the gastric wall as well as sampling of the cyst contents for both fluid analysis and cytology.
Several biochemical markers have been measured in the cyst fluid in an attempt to distinguish mucinous cysts with malignant potential such as IPMN and MCN from non-mucinous, benign cysts such as inflammatory pseudocyst and serous cystadenoma (SCA); further, the recent addition of molecular analysis of cyst fluid holds the promise of distinction between low-risk and high-risk mucinous lesions in the future.
Among the plethora of biochemical markers that have been studied, carcinoembryonic antigen (CEA) in cyst fluid has proven to be the most useful in distinguishing mucinous from non-mucinous cysts, although it has no utility in identifying malignant lesions [70]. The results of the Cooperative Pancreatic Cyst study evaluating various cyst fluid tumor markers (CEA, CA 72-4, CA 125, CA 19-9, and CA 15-3) revealed that CEA is the most accurate marker for differentiating mucinous vs. non-mucinous cysts, with a sensitivity and specificity of 75% and 84%, respectively, using an optimal cutoff value of 192 ng/ mL [71]. These findings were then corroborated by a meta-analysis of 18 studies using the same cutoff value that demonstrated a pooled sensitivity and specificity of 63% and 88%, respectively, and a likelihood ratio of 4.27 in identifying mucinous cysts [72]. Different cutoff values have been proposed by other groups, often trading off sensitivity for specificity. A pooled analysis of 12 studies showed that a cutoff value of >800 ng/mL is 48% sensitive and 98% specific for mucinous cysts [73]. The Pancreatic Cyst Fluid DNA Analysis (PANDA) study reported that a value of >148 ng/mL captured 74% of mucinous cysts [74]. Lastly, the updated experience from the Massachusetts General Hospital shows that a new cutoff value of 109.9 ng/mL has a sensitivity of 80.9%, specificity of 97.7%, and overall accuracy of 86% in detecting neoplastic mucinous cysts [70]. The results of cyst fluid CEA as a predictor of malignancy are not as promising. Multiple results have demonstrated that CEA levels have no correlation with the degree of dysplasia and cannot predict malignancy in IPMN [70,75,76].
Despite the encouraging results observed in CEA performance, one important pitfall is the amount of cyst fluid needed to run this assay. On average, 1 mL of cyst fluid is needed for CEA analysis [77]. With the increasing number of small–and often millimetric–pancreatic cysts being discovered and investigated, it has been and will be a challenge to aspirate sufficient amounts of cyst fluid for the assay. Now, molecular analysis of cyst fluid has risen as an alternative that might help clinical decision-making for small pancreatic cysts, given that only ~250 μL of fluid are needed to perform the analysis.
DNA: As discussed above, IPMN exhibits a complex array of molecular alterations throughout the course of their progression from low-grade lesions to invasive carcinoma. The goal of molecular analysis is to detect relevant mutations in the aspirated cyst fluid that allow us to differentiate IPMNs with high-grade dysplasia and/or invasive carcinoma, which need to be resected, from IPMNs with lower-grade dysplasia and other benign entities, which can be followed without surgical intervention. Several oncogenes and tumor suppressor genes have been assessed in cyst fluid including KRAS, CMM/RIZ, VHL, APC, P16, PTCH, PTEN, and TP53, among others [78]. Of these, point mutations in KRAS have received a special attention, followed by loss of heterozygosity (LOH) and DNA quality and quantity.
KRAS mutations in cyst fluid have been found to be highly specific for mucinous cysts, with specificities ranging from 92% to 100% [74,79,80]. However, it yields a large number of false negative results (sensitivity of 11%-45%) [74,79,80]. The PANDA study found that a specific sequence of KRAS mutation followed by allelic loss has been found to be 100% specific for mucinous cysts [74]. LOH alone has a sensitivity and specificity of 50% and 71%, respectively [79]. Overall molecular analysis including evaluation of DNA quantity, KRAS mutation and allelic imbalance has been found to have a sensitivity and specificity of 77% and 100%, respectively [80].
The performance of molecular analysis to predict malignancy has also been studied. The PANDA study demonstrated that DNA amount and quality, number and sequence of mutations (i.e. KRAS followed by allelic loss), and mutation amplitude are all significantly associated with malignancy. Mean allelic loss imbalance (MALA) was the most accurate test for detecting malignancy (sensitivity and specificity of 82%). In turn, KRAS mutations followed by allelic loss were the most specific (96%) [74].
Given the discovery of GNAS mutations in IPMN, it is anticipated that GNAS mutation assays will play an import role in cyst fluid analyses. As described above, mutations in GNAS have been reported in a high proportion of IPMN tissue samples. GNAS mutations in codon 201 were also observed in 61% of 84 cyst fluid samples obtained from IPMNs [28]. More importantly, the combination of GNAS and KRAS mutation assays demonstrated high sensitivity (96%) and specificity (100%) for distinguishing between SCA and mucinous cysts (i.e. MCN and IPMN). The evaluation of von Hippel-Lindau gene (VHL) that is often mutated in SCA may enhance the performance of cyst fluid analysis in differentiating the 2 categories [81]. Moreover, the presence of GNAS mutations can also distinguish between IPMN and MCN, although it is not as sensitive as we would hope. Unfortunately, GNAS fail to confidently predict the grade of dysplasia and presence of malignancy in IPMNs [28].
Das-1 monoclonal antibody
Das-1 is a monoclonal antibody with high specificity for normal colonic mucosa [82]. It has been found to be reactive to pre-malignant conditions of the upper GI tract, such as intestinal metaplasia of the stomach and Barrett’s esophagus, and thus its ability to identify highrisk IPMN is being evaluated. We have reported preliminary results of a study revealing that mAb Das-1 is very specific to high-risk or malignant IPMN [83]. Cyst fluid from 16 patients with cystic tumors was analyzed by ELISA and western blot. Fluid from benign lesions (benign cystic neoplasms and pseudocysts) and low-grade IPMN revealed very little reactivity to mAb Das-1, whereas fluid from highrisk and malignant IPMNs showed significantly higher reactivity compared to both benign lesions (p=0.0019) and low-grade gastrictype IPMN (p=0.0013). Furthermore, these results were corroborated by immunohistochemistry on tissue samples from 108 non-invasive IPMN and 11 invasive IPMN from 77 different patients. mAb Das-1 reactivity was significantly higher in high-risk and malignant lesions: intestinal-, and oncocytic- and high-grade gastric-type; tubular and colloid invasive carcinoma. Conversely, normal pancreatic ducts, lowgrade and intermediate-grade gastric-type IPMNs were either nonreactive or minimally reactive. These results yield 77% sensitivity and 95% specificity of mAb Das-1 for high-risk and malignant IPMNs [83].
Strobel et al. recently described distinct specialized gland-like compartments located in the pancreatic duct system, especially in proximal sites, and named pancreatic duct glands (PDG). PDG are gland-like outpouches or coiled glands with mucinous cytoplasm residing within the mesenchymal cuff of large ducts. They have a distinct molecular signature different from that of pancreatic duct epithelium, retaining expression of developmental markers such as Sonic hedgehog (Shh), and exhibiting trefoil factor (TFF)-2 and gastric pyloric gland mucin, MUC6. Furthermore, PDG respond to chronic injury with Shhmediated gastric mucinous metaplasia that resembles PanIN lesions [84]. As described above, there are similarities in the morphology, protein expressions and molecular alterations between PanIN and gastric-type IPMN. Yamaguchi et al. are currently investigating the gastric-type IPMN. Yamaguchi et al. are currently investigating the
IPMN is being diagnosed with increasing frequency, often discovered in asymptomatic patients as an incidental finding. Many of these cysts are small and do not meet criteria for surgical resection; however, some of them may eventually progress and develop invasive carcinoma, while others will remain insignificant with no evidence of malignant progression. Unfortunately, our understanding of the natural history of this disease is limited, and we are not yet able to confidently predict which cysts will progress into invasive carcinoma. Molecular pathology that allows us to gain insight into the biology of the disease may provide us with clues to improve preoperative decision making.
KRAS and GNAS mutations are highly prevalent in IPMN and appear to be an early event in tumor progression. Cyst fluid CEA, KRAS, and GNAS are useful markers for distinguishing mucinous vs. non-mucinous cysts; unfortunately, neither one can accurately predict malignancy. Recent findings in epigenetic alterations and miRNAs in IPMN hold the promise of new and more specific markers for malignancy, such as BNIP3, miR-21, and miR-155. Das-1 monoclonal antibody has risen as a new alternative for predicting high-risk and malignant IPMNs with excellent preliminary results. Finally, the discovery of PDG, a possible cell origin of IPMN, will improve our understanding of the natural history of IPMN.