Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2013) Volume 4, Issue 1
Biomass of phytoplankters in relation to the physical and chemical conditions of the water column in Lake Kuriftu was studied from August 2005 to April 2006. The sampling stations of the present study were selected from contrasting segments of Lake Kuriftu as impacted at shore and non-impacted at the center. Secchi depth was estimated with a standard Secchi disc of 20 cm diameter while pH was measured in situ by a portable digital pH meter and temperature was determined with a digital oxygen meter. Nitrate-N was determined by the Zinc Reduction Procedure while phosphate and silica were measured calorimetrically by the Ascorbic Acid and Molybdosilicate methods. Phytoplankters samples were examined with an inverted microscope and identification to genus or species level was made on the basis of the various taxonomic literatures available on phytoplankters. The concentration of Chlorophyll a was calculated by using absorbance measurements made at 665 and 750 nm to calculate the biomass. All the physical, chemical and biological parameters measured varied spatially and temporally. The Lake’s transparency (ZSD) was always less than 0.6 m with smaller values coincident with periods of rainfall and negatively correlated (r=-0.36) with phytoplankters biomass. The chemistry of the lake was basically similar to the dilute East African lake waters, with maximum pH and total alkalinity values of 8.8 and 3.1 mEq/l, respectively. The inorganic nutrients, which were of moderately high levels varied temporally as a function of hydrological, hydrographic and biological conditions of the lake. The phytoplankters community which was consistently dominated by cyanobacteria exhibited low species diversity. Phytoplankters biomass measured as chlorophyll a varied from 18.35 to 45.18 mg Chl a m-3 at the near-shore station and from 17.24 to 55.6 mg Chl a m-3 at the central station. The variations of the parameters indicates how one is influenced by the other and influences the others in a given ecosystems.
Keywords: Biomass; Species; Chemicals
The availability and sustainable use of natural resources determines the food security situation of a country. In order to meet the growing food requirements of the world population great effort is necessary in the development and utilization of the biological wealth of the aquatic environment. Aquatic ecosystems have received attention as actual and potential sources of food. The functioning of the phytoplankton community is also of both practical and theoretical interest.
Phytoplankters are the major primary producers in many aquatic systems and are an important food for consumers [1]. Through the process of photosynthesis these microscopic plants nourish the entire food web of the oceans. The global carbon cycle, which regulates the temperature of our planet, and the life-sustaining oxygen, which is essential to the metabolism of all aerobic organisms, are controlled by the photosynthetic activity of the phytoplankters. Biomasses of phytoplankters are affected by an array of chemical, physical, and biological factors. There is no doubt that in general the more frequently a lake is stirred by winds to the bottom, the faster the nutrients are recycled from the mud into the photosynthetic zone where they may accelerate the rate of productivity [2]. The dynamics of phytoplankters standing stock and productivity in African lakes was reported to vary intimately with the fluctuation in water level [3,4].
Solar radiation is of fundamental importance to the entire dynamics of an aquatic ecosystem. Nearly all the energy that controls the metabolism of lakes comes directly from solar energy [5]. Light intensity has received the most extensive attention of all the physical factors controlling aquatic primary productivity and it can directly influence the physiology and species composition of phytoplankters. Blooms of blue-green algae are favored by the high temperature of the warmer months of the year [6]. In addition to light energy, phytoplankters are dependent upon adequate supply of nutrients. Algal cells require elements in relatively fixed proportions in order to reproduce. The various nutrients required by algal cells for growth and multiplication may not always be found in relative proportions required by phytoplankters [7]. Phosphorous and nitrogen in all groups and silicon in diatoms are regarded as growth-limiting nutrients.
Compared to many other countries in the world, Ethiopia is endowed with a large number of standing water bodies, whose sustainable use can contribute to the economy of the country. The lakes are critical to the survival of local communities as they are the actual and potential sources of food and income. Furthermore, their range of variations in morphometry, physical and chemical features offers opportunities for superb comparative limnological studies. Although primary productivity of phytoplankters in Ethiopian lakes is one of the well-studied aspects, only few studies were conducted on long-term basis. The long term investigations on primary productivity of phytoplankters include those of in Lakes Kilole and Arenguade, in five Ethiopian Rift Valley Lakes, and in Lake Awassa, in Lake Ziway, in Lake Chamo and in Lakes Awassa, Chamo and Ziway [8-13]. Published information on the photosynthetic primary productivity, algal biomass and species composition of phytoplankters in Lake Kuriftu is, however, not well studied.
In order to obtain adequate understanding of the temporal dynamics of the lake biology and the underlying environmental variables, one needs to study the changes in species composition, relative abundance, primary productivity and biomass of phytoplankters in relation to some physical and chemical factors. The general objective of this research study was, therefore to study the temporal and spatial dynamics of photosynthetic productivity of phytoplankters, species composition in relation to some physical and chemical factors in Lake Kuriftu in consideration of the human impacts on the lakes chemistry and biology in the long term.
Description of the study area
Lake Kuriftu is one of the lakes found in and around the town of Bishoftu. It is found at an altitude of 1860 m, some 47 Km southeast of Addis Ababa. The lake is located at 8° 47’ N and 39° 00’E. It is a shallow ( ≈ 6 m) with an area of 0.4 km2 and has a volume of 3.0×106 m3, Secchi depth of 0.15-0.20 m and pH of 7.9-8.4 [14] lake formed by diverting and damming the tributary of the perennial Mojo River, Belbela River, for the irrigation practice in the area [15].
The lake gains water primarily from the Belbela River, with a small contribution from precipitation. Groundwater inflow plays a minor role in the water balance of this lake as the static water level in the area is well below the lake and the occurrence of loss of water through seepage is not well known [15]. The region around the lake is characterized by moderate rainfall, varying around about 850 mm per annum, high incident solar radiation and low relative humidity. The region has two rainy periods, the minor one extending roughly from February to April and the major one beginning in June and ending in September [16]. The trees found around the lake include Accacia abyssinica, Jacaranda mimosifolia and species of Eucalyptus and Juniperus. Macrophytes including Passifloraceae and Passiflora subpeltata ortega were observed.
Sampling protocol
The sampling stations of the present study were selected from contrasting segments of Lake Kuriftu. Two sampling stations were selected: one from an area of high human impact (near-shore station) and another from a relatively less impacted area (central station). Water samples were collected at least once a month from the two stations with a bottle sampler. The water samples were collected from selected depths (0.00, 0.5, 1.0, 1.5, 2.0 m) distributed within the euphotic zone. The samples collected from different depths were mixed in equal proportions to produce composite samples and were used for chemical analyses and measurement of biomass as chlorophyll a concentration.
Measurement of physical and chemical parameters in the field Some physical and chemical parameters, which were believed to have a bearing on the species composition and biomass of phytoplankton, were measured in situ. The parameters measured in situ include the following:
Secchi depth was estimated with a standard Secchi disc of 20 cm diameter.pH was measured in situ by a portable digital pH meter (Jenway 3200).Temperature was determined with a digital oxygen meter (Hanna 9024).
Analysis of inorganic nutrients Water samples filtered through Glass Fiber filters (GF/C) were used for the analyses of Soluble Reactive Phosphate (PO4-P), Silica (SiO2) and Nitrate (NO3-N). Nitrate-N was determined by the Zinc Reduction Procedure [17] while phosphate and silica were measured calorimetrically by the Ascorbic Acid and Molybdosilicate methods respectively.
Measurement of biological parameter
Species composition and abundance of phytoplankton: Major species of phytoplankters preserved with Lugol’s iodine found in samples were identified. The phytoplankters samples were examined with an inverted microscope and identification to genus or species level was made on the basis of the various taxonomic literatures available on phytoplankters [18-22].
Estimation of phytoplankton biomass: Phytoplankters biomass was estimated as chlorophyll a concentration spectrophotometrically from water samples filtered through glass fiber filters (GF/C). Chlorophyll a was extracted from the phytoplankters concentrate with aqueous acetone (90%). The filters were manually ground with a glass rod to enhance extraction of pigments. The concentration of Chlorophyll a was calculated according to Talling and Driver [23] using absorbance measurements made at 665 and 750 nm.
Statistical Analysis: The relationships between the different physical and chemical and biological parameters were tested statistically using Minitab ver.14.
Physico-chemical features
Physical Parameters: The physical characteristics of Lake Kuriftu during the study period are given in Table 1. Lake’s transparency (vertical visibility) varied between 0.35 m in April, 2006 of the minor rainy season and 0.6 m in December, 2005 of the dry season at the open water station. The high Secchi depth values of Lake Kuriftu are greater than those recorded for Legedadi (0.082-0.11 m;) [24] and Koka (0.28 m) [25] reservoirs and Lakes Ziway (0.35 m), Abaya (0.43 m) [26], and Chamo (0.21-0.375 m) [12], in which attenuation of underwater light is primarily due to silt and clay, which are known to form a stable colloidal suspension in Koka Reservoir and Lake Langano [25,27,28].
| Date | Surface water temperature (°C) | ZSD(m) | Kd(units m) | Zeu(m) |
|---|---|---|---|---|
| 8-08-05 | 29 | 0.51 | 2.82 | 1.63 |
| 27-08-05 | 31.5 | 0.55 | 2.62 | 1.75 |
| 8-10-05 | 31.1 | 0.53 | 2.72 | 1.69 |
| 15-10-05 | 31 | 0.50 | 2.88 | 1.59 |
| 29-10-05 | 28.5 | 0.55 | 2.62 | 1.75 |
| 19-11-05 | 26.4 | 0.55 | 2.62 | 1.75 |
| 15-12-05 | 33.3 | 0.6 | 2.4 | 1.92 |
| 9-01-06 | 22.8 | 0.51 | 2.82 | 1.63 |
| 5-02-06 | ND | 0.52 | 2.77 | 1.66 |
| 1-03-06 | 30.1 | 0.55 | 2.62 | 1.75 |
| 25-03-06 | ND | 0.58 | 2.48 | 1.85 |
| 26-04-06 | 24.2 | 0.35 | 4.11 | 1.12 |
Table 1: Surface Water Temperatures (°C), Secchi depths (ZSD), Mean vertical extinction coefficients (Kd) and Euphotic depths (Zeu) of the central station in Lake Kuriftu August, 2005 to April 2006).
But, the lakes found in the region of the present study lake, Lakes Kilole (ZSD=0.37-1.8 m) [29] and Babogaya (ZSD=1.48-4.46 m) [30] have much higher transparency than Lake Kuriftu. The temporal variations seen in the transparency of Lake Kuriftu seem to be related to changes in the extent of re-suspension of inorganic particles resulting from wind-driven mixing, variation in phytoplankters biomass and external loading of particulate materials through runoff. The correlation between Secchi depth readings and phytoplankters biomass was negatively correlated (r=-0.36) probably indicating the greater importance of a biogenic turbidity to the underwater light climate of Lake Kuriftu.
Vertical coefficients for total underwater light (mean vertical extinction coefficient, Kd) can be estimated from Secchi depths using the widely used equation, which was proposed by Holmes [31] as a better estimator of the vertical extinction coefficient of total underwater light of turbid waters (Table 1). The values of Kd, calculated according to Holmes [31], ranged from a minimum of 2.4 to a maximum of 4.11 corresponding to euphotic depths (Zeu), approximated as Zeu=4.6/ Kd [32], of 1.12 m and 1.91 m. The calculation of euphotic depth using the equation given above assumes that light reduction conforms, approximately, to a single average vertical extinction coefficient, Kd.
Euphotic depths (the depth at which 1% of the surface irradiance is detected) estimated using vertical extinction coefficients calculated from Secchi depths seem to indicate that the vertical extent of the euphotic zone was always shallower than 2 m. The surface water temperatures at the central station of Lake Kuriftu ranged from a minimum of 22.8°C in February, 2006 to a maximum of 33.3°C in January, 2006 with most values between 28°C and 30°C. The maximum surface water temperature of Lake Kuriftu is closer to those of Lake Chamo (26-30°C) [12] in Ethiopia and Lake George, in Uganda (26- 36°C) [33]. But, greater than those reported for the Ethiopian Rift Valley Lakes including Lakes Ziway (18.5-27.5°C) [11], Abijata and Langano (18-27°C) [26] and Awassa (23.8-28.4°C) and other lakes of the same region as the present study lake, Lakes Kilole (18.5-24°C) [29], Babogaya (20.5-28.4°C) [30] and the Legedadi Reservoir (22.2- 23.9°C) [24].
The vertical distribution of temperature in Lake Kuriftu shows the occurrence of small differences between successive depths of the water column down to 4 m. In this lake there seemed to be no thermal stratification considering its shallow depth and exposure to wind action although the oxygen-temperature meter used did not allow the determination of temperature at depths deeper than 4 m. The thermal regime of Lake Kuriftu is probably comparable to the shallowest Crater Lake Kilole and Lake Ziway [11] which are almost continually stirred to their bottoms.
Water column mixing in the Ethiopian Rift Valley lakes is frequent [27] for there has been no indication of strongly developed persistent thermal stratification. Studies on the thermal characteristics of the Bishoftu crater lakes in Ethiopia were made [34]. The two deepest Lakes, Bishoftu (87 m) and Pawlo (65 m), showed the most stable thermal stratification and an anoxic layer. Baxter et al. (1965), in their studies on African lakes, have noted that complete mixing is normally frequent in lakes with maximum depth (Zmax) of less than 15-30 m and thermal stratification is largely diurnal, a water column condition documented for a large number of Ethiopian Rift Valley lakes [25].
Chemical features of Lake Kuriftu
pH of Lake Kuriftu: The pH of the surface water of Lake Kuriftu at the central station ranged from a minimum of 8.2 in August, 2005 to a maximum of 8.8 in November, 2005. The pH values recorded for this lake in the present study are slightly higher than those reported in an earlier investigation (7.9-8.4) [14]. The pH values of Lake Kuriftu, which are comparable to those of Lake Awassa were generally lower than those recorded in the nearby crater lakes, Lake Bishoftu [35,30] Babogaya [30] and the Rift Valley Lake Chamo (8.53-9.44) [12].
pH values, which are remarkably different from those observed for Lake Kuriftu were reported only from saline lakes, including Red Rock Tarn in Australia [36], Mariut in Egypt and Arenguade in Ethiopia [8], which have pH values between 9.0 and 10.5 that tend to remain at high levels owing to the high buffering capacity of the lake waters [2].
Ambient Concentration of Inorganic Nutrients: The temporal variations in the concentration of inorganic nutrients in relation to phytoplankton biomass at the central and near-shore stations of Lake Kuriftu are shown in Table 2 and Figure 2. Concentrations of soluble reactive phosphate (PO4-P) recorded in this study ranged from a minimum value of 8.64 in August, 2005 to a maximum of 41.5 μg/l in March, 2006 at the near-shore station, and from 10 in August, 2005 to 51 μg/l in February, 2006 at the open station. The maximum level of soluble reactive phosphate (SRP) observed in Lake Kuriftu is much lower than those reported for other crater lakes of the same region including Lakes Bishoftu (280 μg/l;) and Arenguade (3200 μg /l) [26,35] and the rift valley lakes Ziway (mean value of 90 μg/l) and Lake Chamo (26.4-91.7 μg /l) [12] although it is still higher than values recorded for other nearby crater Lakes Babogaya (1-11 μg/l); [30] and Kilole [3,35].
| NO3-N (µg/L) | PO4-P (µg/L) | SiO2(mg/L) | B (mg Chl a m-3) | |||||
|---|---|---|---|---|---|---|---|---|
| NS | Op | NS | Op | NS | Op | NS | Op | |
| 8-08-05 | 34.67 | 33.3 | 11.926 | 10.2813 | 8.1688 | 7.5946 | 18.348 | 17.236 |
| 27-08-05 | 35 | 36 | 8.6363 | 21.796 | 6.9247 | 7.0204 | 23.352 | 35.028 |
| 8-10-05 | 40 | 40.6 | 20.151 | 34.956 | 7.1161 | 7.4145 | 22.24 | 31.692 |
| 15-10-05 | 33.3 | 33.3 | 10.2813 | 15.2163 | 7.3075 | 7.4032 | 33.638 | 41.144 |
| 29-10-05 | 34.6 | 35 | 9.9113 | 13.5713 | 9.4129 | 11.039 | 39.6 | 55.6 |
| 19-11-05 | 35 | 37.3 | 10.2813 | 13.5713 | 8.743 | 8.9344 | 28.078 | 46.426 |
| 15-12-05 | 33.3 | 34 | 13.5713 | 14.0648 | 8.3602 | 10.465 | 32.248 | 46.426 |
| 9-01-06 | 34 | 35.3 | 30.02 | 11.926 | 9.2215 | 8.743 | 34.472 | 47.816 |
| 5-02-06 | 33.3 | 36.7 | 38.2463 | 51.4063 | 7.656 | 8.1345 | 37.805 | 39.476 |
| 1-03-06 | 36 | 34 | 41.5363 | 36.6013 | 6.6376 | 7.9774 | 44.13 | 40.31 |
| 25-03-06 | 34.6 | 33.3 | 39.5211 | 31.66 | 5.4892 | 6.6376 | 39.6 | 32 |
| 26-04-06 | 46.7 | 43.3 | 39.891 | 34.956 | 6.829 | 7.2118 | 45.175 | 43.09 |
Table 2: Ambient concentrations of NO3-N, PO4-P and SiO2 in relation to Phytoplankters biomass as Chl a (B) at the open (OP) and Near-Shore (NS) station of Lake Kuriftu.
Figure 1: Map showing the relative location of Bishoftu Lakes with the sampling stations in lakeKuriftu indicated [source:Lamb (2001)].
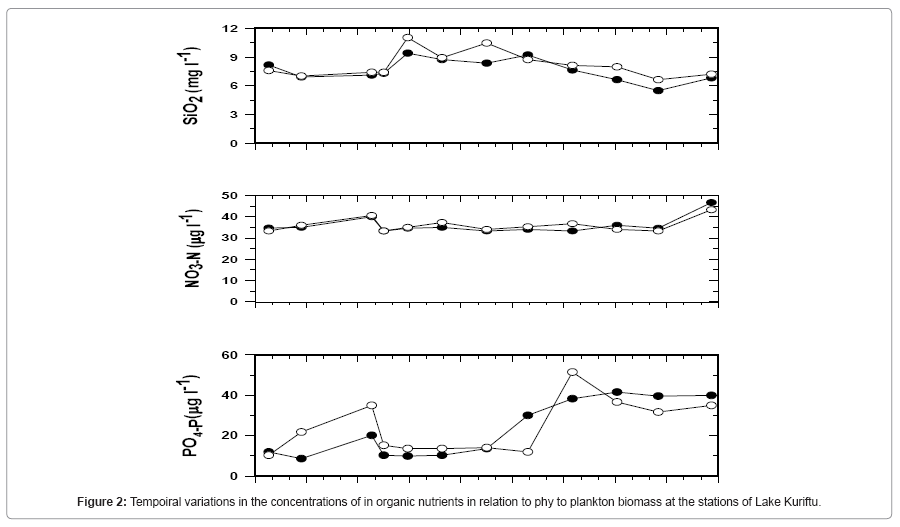
Figure 2: Tempoiral variations in the concentrations of in organic nutrients in relation to phy to plankton biomass at the stations of Lake Kuriftu.
Most of the time phosphate concentrations were low on days of high phytoplankters biomass and higher at the central station than at the near- shore station. The slightly higher concentrations of phosphate at the central station were probably associated with the greater exposure of the station to wind blowing over this small lake whose shore regions are better protected by the elevated surrounding land. It is also possible that there is greater external loading of phosphate through the shoreline which closer to the central station than the near-shore station.
Nitrate-N (NO3-N), which showed less temporal and spatial variations than phosphate and silica, varied from 33.3 to 46.7 μg/l at the near-shore station and from 33.3 to 43.3 μg/l at the open station with the maximum values in April, 2006 during the minor rainy season. The levels of nitrate determined in the present study in Lake Kuriftu are higher than the values recorded for other crater lakes Babogaya (1-31μg/l) [30] and Bishoftu (25 μg/l) [35] and Lakes Awassa [26]. But considerably lower than Lake Ziway (28-136.5 μg/l) [13]. The high nitrate concentrations at times coincided with relatively low phytoplankters biomass and vice-versa at the open station although the relation between nitrate levels and phytoplankters biomass was not apparent for the near-shore station in Lake Kuriftu.
Molybdate reactive silica (SiO2) also showed small temporal variations at both the open and near shore stations. The values recorded for the near-shore station ranged from 5.49 to 9.14 mg/l while those for the open station varied from a minimum of 6.64 to a maximum of 11.1 in March 2006 and October 2005, respectively. The concentrations of silica were generally higher at the open station than at the nearshore station. The higher concentrations of silica at the open station are probably attributable to the greater input of the nutrient through runoff from the steep slopes found on the side of the open station. The most plausible explanation seems that internal loading of silica was higher at the open station owing to its position relative to the effect of wind blowing over this small- sized lake. Furthermore, the seasonal peaks in silica concentrations at both stations were observed during the dry season suggesting the greater importance of autochthonous sources of silica.
Although the observed concentrations of silica never approached that regarded as limiting to diatom growth (ca, 0.3 mg l-1) [1] they are low in view of the high concentrations (>10 mg l-1) commonly encountered in African lakes [37] including the nearby crater lakes Kilole [38], Hora [38] and Babogaya [30] and the Ethiopian Rift Valley lakes Ziway [8], Koka [39,40]. The only lake for which silica concentrations considerably lower than those of Lake Kuriftu were reported is Lake Chamo of the Ethiopian Rift Valley (0-4 mg l-1) [26,41].
Biological features
Species composition and abundance of phytoplankters: Table 3 and Figure 1 present a list of the major phytoplankters species identified in samples collected during the study period and the relative abundance of groups of phytoplankton respectively. A total of 25 species of phytoplankters belonging to 6 classes were identified. The species composition and diversity of phytoplankters of Lake Kuriftu are similar to those of Lake Babogaya [30]. Phytoplankters diversity in Lake Kuriftu (25 spp) is low when compared to those of the Ethiopian Rift Valley Lakes Ziway, Awassa, and Chamo [42]. Prosser et al. [38] has also recently reported similarly high species diversity for Lakes Ziway, Awassa and Chamo in which the cyanobacteria were qualitatively and quantitatively important.
| Phytoplankton group | Species name |
|---|---|
| Cyanophyceae Cyanobacteria (Blue-green algae) | Cylindrospermopsis africana Kom. and Kaling C. Curvispora M.Watanbe Planktolyngebya tallingii Kom. and Kaling Planktolyngebya contorta (Lemm.)Anagn. And Kom. Microcystis aeruginosa Rab. Anabaena circinalis Rab. Anabaena nygaerdinom. Psuedoanabaena sp. Raphidiopsis |
| Chlorophyceae (Green algae) |
Pediastrum simplex Meyen P. duplex Meyen Scenedesmus armatus Chod. Scenedesmus dimorphus (Turp.)Kutz. Scenedesmus quadricauda (Turp).Breb. Chlamydomonas reticula Phacotus lenticularis (Ehr.) Stein |
| Bacillariophyceae (Diatoms) |
Thalassiosira sp. Navicula cryptocephala Kutz. Nitzschia vermicularis(Kutz.)Grun. N. rostellate. Rhopahodia sp. |
| Dinophyceae (Dinoflagellates) | Peridinium sp. |
| Cryptophyceae (Cryptomonads) | Cryptomonas obovata Skuja |
| Euglenophyceae(Euglenoids) | Phacus longicauda (Ehr.) Duj. Lepocincilis sp. |
Table 3: List of the major species of phytoplankton identified from Lake Kuriftu during the study period
Blue green algae, green algae and diatoms were the major algal (taxonomic) groups in terms of species richness and abundance. The other taxonomic groups- dinoflagellates; cryptomonads and euglenoids -were poorly presented. Microcystis aeruginosa was usually the most important in terms of abundance and formed the most conspicuous populations. The temporal changes in the percentage contributions of the different algal groups to the total abundance of the phytoplankters community are shown in Figure 3. Temporal changes in the relative importance of the different taxonomic groups to the abundance of the phytoplankters assemblage in the Lake Kuriftu were observed.
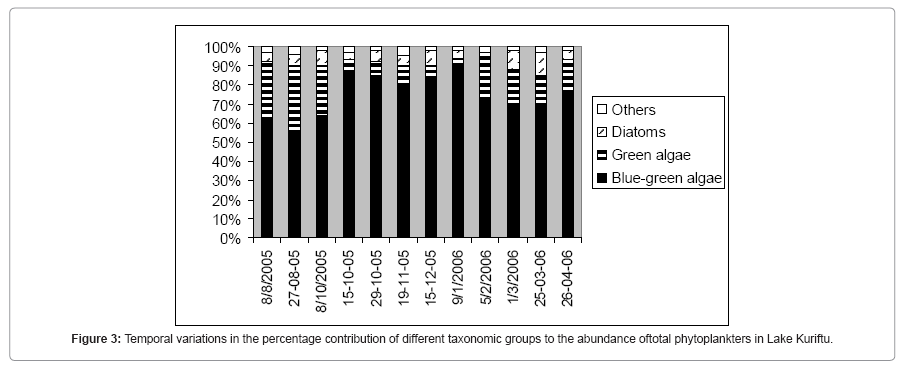
Figure 3: Temporal variations in the percentage contribution of different taxonomic groups to the abundance oftotal phytoplankters in Lake Kuriftu.
The Blue-green algae, with 9 species, were the most diverse and dominant taxonomic group with contributions to the abundance of the phytoplankters assemblage in Lake Kuriftu ranging from a minimum of 56% in August, 2005 to a maximum of 91% in January, 2006. Among the blue greenalgae Microcystis aeruginosa and Cylindrospermopsis africana were the most important in terms of abundance. Green algae, which were of secondary importance to the total abundance of the phytoplankton assemblage, had contributions that ranged from 3 to 34% and which were primarily constituted by Pediastrum duplex and P. simplex from the 7 component species. Diatoms were the third important taxonomic group with contributions much lower than those of the green algae (2 to 12%).
Phytoplankters biomass: The Phytoplankters biomass of Lake Kuriftu exhibited temporal and spatial variations over the study period (Table 2). The values recorded ranged from 18.35 to 45.18 (mg Chl a m-3) at the near- shore station and from 17.24 to 55.6 (mg Chl a m-3) at the open station. The lowest phytoplankters biomass measured as Chl a was observed in August, 2005 at both stations. This seasonal minimum of Chl a coincided with a period of heavy precipitation that resulted in land runoff which brought particulate materials into the lake with consequent reduction in light penetration. The occurrence of low phytoplankters biomass in lakes during periods of heavy rainfall is not unusual and has been reported for Lake Victoria. The peak in phytoplankters biomass (45.18 mg Chl a m-3 ), which was observed in April, 2006 for the near-shore station, coincided with an increase in NO3-N and PO4-P levels while that of the open station (55.6 mg Chl a m-3 )occurred in October, 2005 concomitant with a relatively low level of phosphorus and a slight increase in NO3-N.
The maximum phytoplankters biomass value of Lake Kuriftu is closer to those reported for Lakes Bishoftu [43] and Ziway [38] and Legedadi Reservoir [24] but, higher than those observed for the nearby crater lakes Babogaya [30]; Hora [43]; the Ethiopian Rift Valley lake Awassa [13] and Koka Reservoir. Much higher phytoplankton biomass values are known from a number of shallow East African lakes including Kilole and Arenguade in Ethiopia [8] and Lakes Nakuru [4], and Simbi [41] in Kenya and Lake George in Uganda [33]. Most of the time the biomass in Lake Kuriftu was higher at the open station (mean=39.69 mg Chl a m-3) than at the near-shore (mean=31.52 mg Chl a m-3) station. This seems to be associated with the generally higher concentration of nutrients and transparency (less turbidity) at the open station than at the near-shore station. Similar spatial trends were also reported for Lake Ziway in which the near-shore station had lower biomass than the open station and Lake Babogaya [30].
Although both phytoplankters biomass and all inorganic nutrients showed spatial variations, the differences between the two stations in the concentrations of nitrate and phosphate (T-value =0.25 for nitrate, 0.48 for phosphate) were not statistically significant while those of Chl a and silica were as the t-test values (2.34 for silica and 2.57 for chl a) indicate significancy. In Lake Kuriftu, the correlations between biomass and the macronutrients nitrate-nitrogen (r=0.109) and phosphate (SRP) (r=0.14) were positive but weak while that between silica and biomass was relatively strong (r=0.691) although diatoms were not as important as the other algal groups in Lake Kuriftu. Temporal dynamics of phytoplankton biomass are known to be controlled by loss processes including grazing by zooplankton and fish. The Cyclopoid copepod, Thermocyclops consimilis dominates the zooplankton community in Lake Kuriftu, followed by rotifers and lower abundance of cladocerans [44]. The feeding habit of Termocyclops is, however, is less defined [45]. The Research conducted has shown the dominance of copepods throughout the study period except in September and October, 2005 when rotifers became more important [44]. In October, 2005 there was a decline in phytoplankters biomass, which was probably associated with the peak abundance of rotifers. Rotifers particularly Brachionus species can have a considerable effect on phytoplankters as they are known to resist the toxicity of blue greens although they mainly ingest particulate material [46].
Carlson’s Trophic Index which relates Secchi depth and concentrations of chlorophyll a, and phosphorus, is a common means of characterizing the trophic state of a lake. According to Carlson (1977), lakes with Trophic Status Index (TSI) ranging from 60 to 70 are eutrophic with dominance of blue-green algae. Lake Kuriftu whose TSI for chlorophyll a and Secchi depth are 66.59 and 69.7 respectively is, therefore classified under the category Eutrophic one in which bluegreen algae are the dominant algal groups [47].
Lake Kuriftu is a shallow, frequently mixing, moderately turbid and productive lake. The lake exhibits temporal variations in photosynthetic biomass and species of phytoplankters. These variations seem to be related to hydrological and hydrographic conditions of the lake, which in their turn determine the levels of nutrients and their spatial variations as Talling has pointed that the temporal changes in biological parameters of lakes are related to input-output relationships of energy and matter, which are dictated by hydrographic (water column structure and circulation) and/or hydrological (water inputoutput) conditions. It was also singled out that flood- related changes in nutrient supply and abiotic turbidity are the over-riding factors in shallow well mixed lakes such as Lake Chilwa. Temporal variations in species composition and relative abundance of different taxonomic groups of phytoplankton were observed. It was the blue-green algae of the genera Microcystis and Cylindrospermopsis, which persistently dominated the phytoplankters community of the lake. The persistence of fairly high levels of nutrients, particularly phosphate in a generally turbid and mixing water column favors the dominance of blue-greens owing to their structural and physiological adaptation for buoyancy regulation.
I would like to express my heartfelt gratitude to my research advisor, Dr. Demeke Kifle, for his unreserved and committed consistent support and guidance in the work of my research study. I also express my gratitude to Dr. Seyoum Mengistu and Dr. Abebe Getahun.Addis Ababa University, Department of Biology deserve special thanks for providing me the financial support and vehicle during the study period. My special thanks also go to my mother Adanech Ulfina and my sister Talile Dessalegn. Finally my thanks also go to Kuriftu Childrens and Integrated Farm Center.