Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Short Commentary - (2015) Volume 3, Issue 2
Keywords: Obesity, Adipose tissue, Mutations
There is a worldwide epidemic of obesity, which predisposes to Cardiovascular Diseases (CVD), type2 Diabetes Mellitus(DM), hypertension, stroke and many cancers, not only in developing world but the incidence is rising dramatically in developing countries too [1,2].
Although bariatric surgery offers some form of relative permanent relief it is not only costly, but efforts are required to maintain the weight lost, besides the risks to life with surgery in such obese population carrying so much metabolic dysfunction.
Hence, a demand for more nonsurgical medical pharmacological therapies is rising. Unfortunately so many oral antiobesity drugs discovered have either been banned like sibutramine, rimonabant due to undesired side effects, or a tolerance has been developing to monotherapies like topiramate and hence, Combination Therapies (CT) are looked for . Even with CT cardiovascular side effects are prevalent with Qsymia, hence one need to look for other novel therapies.
Recently interest has emerged towards development of pharmacotherapies which try to augment Brown Adipose Tissue (BAT) to raise energy expenditure following the accidental discovery and presence of BAT in adult humans as well besides their presence in smaller mammals and human infants, when on using 18F FDG/ PET scanning in cancer patients several discrete areas of metabolically active BAT was suggested [3]. Further recent data indicate that normal adult humans contain significant depots of UCP-1 positive brown fat which can be detected by FDG/PET/CT scanning methods especially in supraclavicular and neck area [4-8]. On 18FFDG/PET they found that the amount of active tissue is inversely associated with BMI and the activity increased by cold exposure. Further studies by Zingarelli showed that this tissue has histological appearance of AT, expressing AT molecular markers, with UCP 1 being one of them [9]. In several studies it has been shown that beige selective markers were expressed more in human brown fat samples, as compared to classical brown fat of rodents [10-12]. Wu have cloned beige cell lines while brown fat cell lines have also been cloned with identification of unique molecular markers of each cell types [10]. Very recently Shinoda has characterized a human brown fat cell line from stromal vascular fraction which share more molecular features with beige as compared to brown adipocytes [13]. In some adult humans both brown and beige characteristic possessing cell characteristics possessing cells have been identified according to the depth of the neck [12,14,15].
Over a century Adipose Tissue (AT) in mammals has been classified to two major types namely, white and brown. WAT has been highly adapted to store excess energy in the form of triglycerides, BAT, on the other hand oxidizes chemical energy to produce heat to combat hypothermia and prevent obesity. WAT develops in distinct intraabdominal depots and in the subcutaneous layer. Visceral WAT is the one which correlates with the development of the metabolic syndrome and CVD, while the expansion of subcutaneous WAT, and has been suggested to promote insulin sensitivity [16,17].
Atit and colleagues observed that cells derived from the central dermatomyotome, molecularly defined as cells that sometime in their development had expressed the homeobox transcription factor-Engrailed 1, developed into three types of tissue, dermis, muscle and BAT [18].
Timmons found that cell cultures of precursors from BAT (not WAT precursors) usually demonstrated remarkable expression of genes that had always been considered to be muscle specific. Hence a myogenic signature was expressed in brown adipocyte, not shared by white adipocytes [19]. Seale by in vivo fate mapping studies demonstrated that cells expressing the myogenic transcription factor myf5 during development develop into cells constituting muscle or BAT but never into the cells found in WAT [20].
Based on examining precursors from the purest WAT depots (epidydimal) Petrovic reported that chronic treatment with the peroxisome proliferator activated receptor γ agonist rosiglitazone promoted the expression of PGC-1α and mitochondriogenesis in these cells, along with norepinephrine augmentable UCP-1 gene expression in a significant subset of the cells which provided the cells with a genuine thermogenic capacity. Although these cells expressed functional thermogenic genes they did not reveal the transcripts of transcription factors which are associated with classic brown adipocytes, namely (zinc in cerebellum (zic 1), lim homebox8 (lhx8), mesenchymal homeobox2 (Meox 2) and characteristically PRDM16) or those genes associated for myocyte, like myogenin and myomirs and retained the characteristics of white adipocytes like Hoxc9 expression. These UCP-1 expressing cells are not proliferating classic brown adipocytes (adipomyocytes) as shown by co-culture studies and hence these cells (Figure1) therefore constituted a subset of adipocytes (“brite” adipocytes) with a developmental origin and molecular characteristic distinguishing them as a separate class of cells [21]. Further studying the gene patterns from 9 different adipose depots, Walden found that each depot displayed distinct gene expression finger profiles. Three major depots were identified:
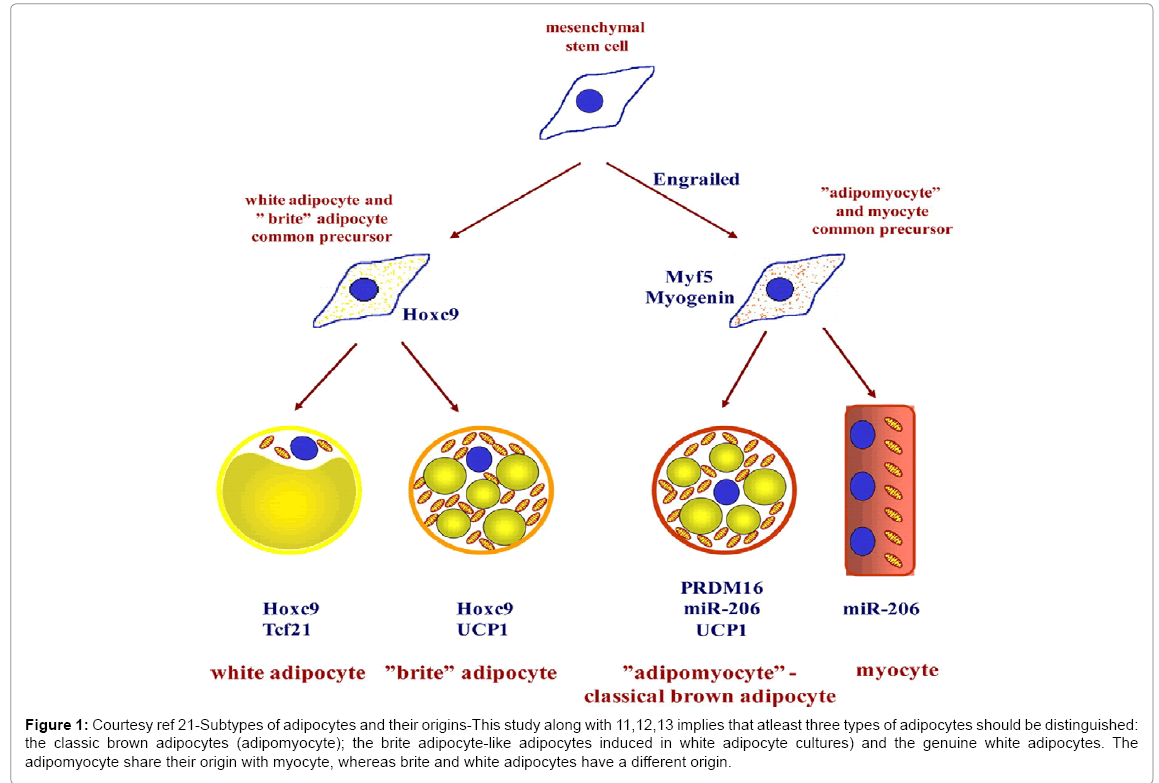
Figure 1: Courtesy ref 21-Subtypes of adipocytes and their origins-This study along with 11,12,13 implies that atleast three types of adipocytes should be distinguished: the classic brown adipocytes (adipomyocyte); the brite adipocyte-like adipocytes induced in white adipocyte cultures) and the genuine white adipocytes. The adipomyocyte share their origin with myocyte, whereas brite and white adipocytes have a different origin.
i) The brown
ii) The brite
iii) The white
Differences in gene expression pattern were generally quantitative but the markers which were consistent were
i) zinc in cerebellum 1(zic-1)for the classic BAT depots, with zic being much more specific as compared to other brown cell markers and in much higher levels like Prdm16, lhx8
ii) Hoxc9 for the brite depots
iii) Hoxc8 for the brite and white in contrast to the brown. Concerning the homeobox genes he proposed it is not presently possible to distinguish whether the expression of these genes should be seen only as (a memory of) positional markers (which would mean that white and brite AT) depots may originate from a narrow range of segments during development or whether they still have functions in determination of the phenotype of the cells.
iv) Tcf21 for the white depots. Further the effects of physiologically recruitment of thermogenic function (cold acclimation) on the expression pattern of the genes were quantified; in general the depot pattern dominated over the recruitment effects. Besides they studied the importance of regulatory roles of various genes like sirtuins, carbonic anhydrase3, short homeobox2 and understanding the developmental background of the depots, as are the regulatory functions of various genes [22].
Enerback in a review showed that Brown fat could improve type 2 DM and metabolic health in various experimental animals [23]. These cells express UCP1 and have a high mitochondrial content and thereby dissipating chemical energy in the face of heat .Seale showed that improvement which occurred in glucose tolerance with browning of white fat (inguinal not epididymal studied earlier and the formation of “beige” or “brite”) cells maybe greater than expected only from the effects on body weight and adiposity. This occurs more by transgenic expression of Prdm 16 in fat depots which causes robust development of brown like adipocytes in subcutaneous but not epididymal adipose depots, leading to increased energy expenditure, limited weight gain and improved glucose tolerance in response to high fat diet [24].
Further adipocyte specific deletion of co-regulatory protein Prdm16 caused minimal effects on classical brown fat but markedly inhibited beige adipocyte function in subcutaneous fat following cold exposure or β3 agonist treatment. Thus, ablation of Prdm16and beige adipocytes caused metabolic dysfunction and to subcutaneous adiposity from visceral fat including decreased thermogenic and increased inflammatory gene expression and increased macrophage accumulation. Thus the benefit of transplantation of subcutaneous fat into mice with diet induced obesity showed a loss of metabolic benefit when tissues were derived from PRDM16 mutants [25].
In vivo fate matting studies using UCP-1 TRAP mouse and corresponding UCP-1 TRAP dataset demonstrated that at least a subset of beige cells arise from a smooth muscle like origin. Ectopic expression of PRDM16 converts vascular smooth muscle cells into Ucp-1positive adipocytes in vivo [26].
Exercise is an important component of good health and represents the first line of therapy for humans with obesity and associated metabolic disorders like type 2 diabetes and hepatic steatosis by increasing energy expenditure. Exercise by increasing physical work, leads to increased consumption of calories, as well as increases energy expenditure both through augmentation of body fat and the browning of white fat [27,28]. Regarding human irisin, it is clear that FNDC5 mRNA is increased in skeletal muscle in some exercise paradigms, but not others [28-30]. Irisin has been found to be deficient inhuman patients with diabetes as compared with normal people [31,32].
Although several polypeptides like Fibroblast Growth Factor 21 ( FGF21), bone morphogenetic proteins7/8-(BMP7/BMP8), Natriuretic Peptides(BNP/ANP), orexin have interesting browning effects, developmental regulators like BMP7/BMP8 are unlike pharmacological candidates, given the pleiotropy of their actions [33-37]. FGF21 is actively being explored as a therapeutic tool, although it is unclear whether its beneficial actions depend on browning require UCP1 or brite iWAT) [38]. Recently Veniant etal further corroborated that, FGF21 does not require UCP1 or brite adipocytes to elicit glucose homeostasis and weight loss showing that during FGF21-Fc treated Ucp 1(-/-) mice showed robust improvement in body weight, glucose homeostasis and plasma lipids associated with increased energy expenditure and FGF21-Fc induced PPargc1 expression in inguinal WAT [39].
Thiazolidinedione increase browning of white fat, but TZD use causes weight gain, not weight loss [21,40]. The natriureteric peptides or inhibitors of natural endopeptidases degrade ANP/BNP are interesting candidates as they are being developed for hypertension and heart failure; but unfortunately blockade of NEP causes weight gain in mice [41,42]. Although COX2 mediates some of the effects of β adrenergic signalling on brown fat development and function, COX2 inhibitors are associated with weight gain [43,44]. Unfortunately promoting global prostaglandin synthesis with a COX2 activator is an unattractive strategy regardless of any potential effects on body weight.
Since irisin is induced during exercise in rodents and is atleast partially responsible for the browning response observed in white fat during chronic exercise this peptide has captured maximum interest [28]. Irisin, a proteolytic hormone derivative (myokine) of the fibronectin III domain containing 5(FNDC5) gene, has been demonstrated in a mouse model to be induced by exercise (mediated by PGC1α and to then directly stimulate phenotypic changes in AT, which mediates changes in systemic metabolism [28]. It is released into the circulation after proteolytic cleavage which occurs at amino acid 30 and 140. The secreted form of FNDC5 contains 112 amino acids and has been named irisin. This irisin preferentially acts on the subcutaneous “beige” fat and causes it to brown by increasing the expression of UCP-1 and other thermogenic genes [10,28]. Wu confirmed that the brown adipocytes detected in 18 FDG/PET scanning studies after cold acclimation, were actually beige adipocytes [10].
The clinical studies by Hu and Lecker have confirmed the positive correlation between increased FNDC5 expression and circulating irisin, with the level of exercise performance [30,45]. FNDC5, besides being expressed also expressed in the brain is also seen in rat pheochromocytoma derived PC12 cells, which get differentiated into neuron like cells [46,47]. FNDC5 knockdown in neuronal precursors impaired the development into mature neurones, which suggests a developmental role of FDNC5 in neurons [48].
Wrann investigated the effects of exercise on FNDC5 expression and function using an established endurance exercise regimen; namely 30days of voluntary free running wheel exercise. This regimen is known to induce BDNF expression, along with neurogenic, dendritic spines and improved memory function in mice [49,50]. The training was sufficient to induce muscle fndc5 gene expression as well as transcriptional regulators pgc-1α, Err–α, known mediators of the exercise response in skeletal muscle. They showed that FNDC5 is elevated in endurance exercise in the hippocampus of mice and that PGC-1α and FNDC5 regulate BDNF expression in the brain [51]. BDNF has also been shown to influence appetite.
Zhang developed a high yield expression of human irisin gene in a yeast expression system. Gene expression studies showed that treating 3T3L-1 derived adipocytes and primary adipocytes with this recombinant–irisin (r-irisin) led to rapid up regulation of brown cell markers, UCP1, PGC1-α, transmembrane protein 26 (TMEM 26), early B cell factor 3 (Ebf3), elongation of very long chain fatty acids like 3 (Elov3) cell death inducing DNA fragmentation factor (Cidea), and cytochrome c oxidase subunit VIIa (COX 7a). It also increased betatrophin (ANGPTL8) gene which affects β cell growth hormone. Mutations of the glycosylated sites in normal irisin asparagine 7 (Asp7and Asp52) led to loss of effects of browning significantly with marked decrease in UCP1 and PGC1α production. Therapeutic effects of r-irisin were further demonstrated by daily ip injection for 14 days which showed mRNA levels of BAT marker genes, UCP1, PGC1-α, COX7A, Elov3, PPARα and Cidea got significantly elevated in AT of normal mice receiving r-irisin and then further proved the mechanism by p38 and MAPK protein kinase pathway was the effect by which irisin produced browning and that significant improvement in glucose parameters and insulin resistance occurred by increased betatrophin gene production. They proposed possibly an irisin receptor is there through which this action occurs although it has to be outlined [52].
Since scale up and production of recombination proteins in yeast is well established, Wu suggested that this irisin reagent can prove to be of great importance in diabetes mellitus and obesity. Hence future steps are applying these and other irisin preparations to models of other diseases. An important factor is trying to identify irisin receptor to open newer possibilities for activation of these areas of exercise science which benefits disorders of liver, heart, muscle and brain (Figure 2) [53].
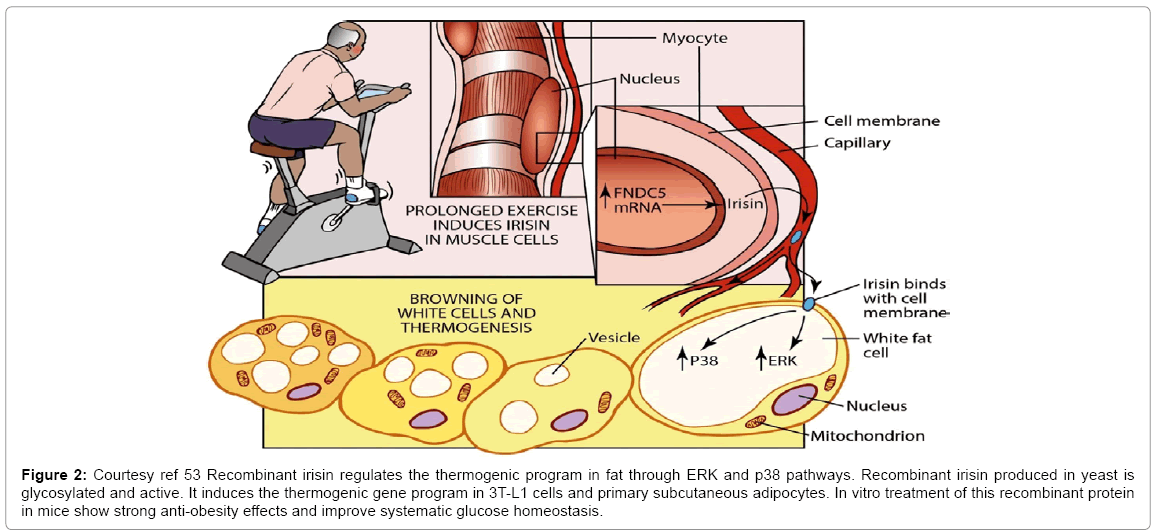
Figure 2: Courtesy ref 53 Recombinant irisin regulates the thermogenic program in fat through ERK and p38 pathways. Recombinant irisin produced in yeast is glycosylated and active. It induces the thermogenic gene program in 3T-L1 cells and primary subcutaneous adipocytes. In vitro treatment of this recombinant protein in mice show strong anti-obesity effects and improve systematic glucose homeostasis.
Other proposed treatment is by modifying microRNA’s to increase energy expenditure after understanding the development of adipocytes for various miR’s which upregulate brown/beige adipocytes [54]. For example overexpression of miR 133a in BAT and SAT cells significantly inhibits and inhibition upregulates Prdm 16 and brown adipogenesis. Even 75% deletion of miR 133a genes results in browning of SAT and such mice express increase insulin sensitivity and glucose tolerance, besides activating the thermogenic gene program more robusltly upon cold exposure [55].
Recently apelin an adipocyte derived hormone, by apelin-AP1 signaling was reported to promote brown adipocyte differentiation, both by increasing brown adipogenic as well as thermogenic transcriptional factors via the PI3K/Akt and AMPK signalling pathway by Than. They further reported that apelin relieves the TNF alpha inhibition on brown adipogenesis, and increased brown adipocyte activity as shown by increased PGC-1 alpha and UCP-1 expression, mitochondrial biogenesis and oxygen consumption. The action of apelin, both in vivo and in vitro to convert white adipocytes to brown like characteristics suggests a potential therapeutic effect to tackle obesity and MS [56].
Merabegron a β adrenergic agonist is used to treat overactive bladder in a dose of 200 mg. The drug has received clinical approval and recently it has been shown to activate human brown adipocytes, which may revoke interest in this pathway [57].
Recently uncouplers /targeted activators of UCP1 or other futile cycles have found to improve insulin resistance, DM, hypertriglyceridaemia by the use of mitochondrial protonophores, which apparently safely uncouples in liver [58].