Journal of Agricultural Science and Food Research
Open Access
ISSN: 2593-9173
ISSN: 2593-9173
Research Article - (2018) Volume 9, Issue 4
Eight endophytic bacterial strains (Bacillus spp., Stenotrophomonas maltophilia, and Pseudomonas geniculata) recovered from healthy cultivated tomato (Solanum lycopersicum L.) were screened for their plant growth-promoting potential on tomato plants challenged with Fusarium oxysporum f. sp. lycopersici (FOL) and for their in vitro and in vivo antifungal activity against FOL. S. maltophilia CT16 and Bacillus subtilis subsp. inaqosorum CT43 and their filtrates were the most efficient in controlling disease by 55-87.5% and in improving growth parameters in inoculated tomato plants by 8.4-46.8%. Pathogen sporulation was inhibited and FOL mycelial growth was reduced using whole-cells and filtrates of the eight strains, and organic extracts from the two active ones. Extracellular metabolites remained effective after heating at 50-100°C with a decline in activity beyond 100°C, when added with proteinase K and their pH adjusted at 2 and 12. Chitinase and surfactin genes were detected using PCR amplification and sequenced for S. maltophilia CT16 and B. subtilis subsp. inaqosorum CT43, respectively. Five strains have shown chitinase- and proteases-activities. B. subtilis subsp. inaqosorum CT43 and S. maltophilia CT16 were able to produce siderophores and salicylic acid. Hydrogen cyanide production was achieved only with S. maltophilia CT16.
Keywords: Biocontrol; Endophytic bacteria; Fusarium oxysporum f. sp. lycopersici ; Growth-promoting; Metabolites; Solanum lycopersicum
Fusarium wilt, caused by Fusarium oxysporum f. sp. lycopersici (Sacc.) W.C. Snyder and H.N. Hans (FOL), is one of the most serious fungal vascular diseases affecting tomato worldwide and which continue to present major challenges for tomato production in Tunisia [1,2]. This soil borne disease is difficult to control due not only to the ability of the pathogen to grow and colonize vascular tissues, and to the long survival of its resting structures i.e., chlamydospores in the soil, but also to the limited range of effective fungicides and resistant varieties [3,4]. Alternative control strategies have been investigated, some of which were more focused on using non-pathogenic microbial agents such as endophytic microorganisms. Endophytes, such as bacteria and/or fungi, may remain at their entry points or spread throughout the plant tissues without causing any harmful effects on their host, which could better limit pathogen progress within vascular tissues [5,6]. They can be isolated from surface-disinfested plant tissues of stems, roots, flowers, leaves, fruits, and seeds or extracted from inside the plant [7,8]. Endophytic bacteria obtained from various plant species have been used for the control of phytopathogenic fungi causing vascular diseases such as Fusarium oxysporum f. sp. vasinfectum on cotton, FOL on tomato and Verticillium dahliae on colza, eggplant and potato [9-13].
Searching for microbial communities associated to tomato plants may contribute to identify potential candidates for biological control of tomato vascular diseases and for plant growth promotion. In fact, this solanaceous species has been well studied in terms of genetics, genomics and breeding but has not been valorized enough as natural source of biocontrol and biofertilizer agents [14]. For instance, Azospirillum brasilense, Burkholderia ambifaria, Gluconacetobacter diazotrophicus , and Herbaspirillum seropedicae were shown able to colonize roots, stems and leaves tissues of S. lycopersicum var. lycopersicum and to stimulate its growth [15]. Furthermore, Brevibacillus brevis W4 recovered from S. lycopersicum stems and leaves have successfully inhibit the development of Botrytis cinerea [16]. Moreover, healthy S. lycopersicum plants have been exploited as natural source of isolation of endophytic bacteria with nematicidal, antibacterial, and/or antifungal activities [17,18]. Biologically active metabolites produced by endophytic bacteria and involved in controlling plant diseases include cell wall-degrading enzymes, lipopeptide antibiotics, and other bio-chemical compounds [10,19,20].
Therefore, the aims of the current study were:
(1) To evaluate, for the first time in Tunisia, the antifungal activity of eight endophytic bacteria isolated from surface-sterilized tissues of healthy tomato plants against FOL.
(2) To assess their ability to promote plant growth.
(3) To elucidate the mechanism deployed by the most active endophytic strains tested.
Plant material
Tomato cv. Rio Grande was used in this study. This cultivar is known by its susceptibility to Fusarium wilt disease incited by Fusarium oxysporum f. sp. lycopersici (FOL) races 2 and 3 [21]. Seedlings were grown in alveolus plates (7 × 7 cm) filled with sterilized peat® (Floragard Vertriebs GmbH fur gartenbau, Oldenburg) under greenhouse conditions (16 h photoperiod, 60-70% relative humidity and air temperatures ranging between 20 and 30°C). They were watered regularly until reaching the two-true-leaf growth stage. Seedlings with approximately similar heights were used for all the in vivo bioassays.
Pathogen culture
F. oxysporum f. sp. lycopersici (FOL) strain used in this study was kindly provided by the Phytopathology Laboratory of the Regional Research Centre on Horticulture and Organic Agriculture at Chott- Mariem, Sousse, Tunisia. FOL strain was cultured for 7 days on PDA medium supplemented with streptomycin sulphate (300 mg/mL w/v) and incubated at 25°C before use.
Endophytic bacteria culture
Eight endophytic bacterial strains (S. maltophilia CT12, S. maltophilia CT13, S. maltophilia CT16, P. geniculata CT19, B. amyloliquefaciens CT32, B. subtilis subsp. inaquosorum CT43, B. licheniformis SV4 and B. subtilis SV5), recovered from healthy tomato plants and selected as the most efficient plant growth promoting strains on tomato free-pathogen were used in this study [22]. Isolation procedure, characterization and identification using 16S rDNA gene sequencing were described in Abdallah et al. [22]. The accession numbers and isolation sources of the eight strains are given in Table 1. Stock cultures were maintained at -20°C in Nutrient Broth (NB) supplemented with 40% glycerol. These bacteria were previously grown on NA and incubated at 25°C for 48 h before use.
| Strain | Species | Accession number | Organ | Localitya | Month (2013) |
|---|---|---|---|---|---|
| CT12 | Stenotrophomonas maltophilia | KR818058 | Fruit | Teboulba, Sousse, Tunisia | April |
| CT13 | S. maltophilia | KR818059 | Root | Teboulba, Sousse, Tunisia | April |
| CT16 | S. maltophilia | KR818060 | Root | Teboulba, Sousse, Tunisia | April |
| CT19 | Pseudomonas geniculata | KR818061 | Stem | Knaies, Sousse, Tunisia | May |
| CT32 | Bacillus amyloliquefaciens | KR818062 | Leaf | M’saken, Sousse, Tunisia | May |
| CT43 | B. subtilis subsp. inaquosorum | KR818063 | Flower | Chott-Mariem, Sousse, Tunisia | November |
| SV4 | B. licheniformis | KR818064 | Stem | Chott-Mariem, Sousse, Tunisia | November |
| SV5 | B. subtilis | KR818065 | Stem | Chott-Mariem, Sousse, Tunisia | November |
Table 1: Endophytic bacterial strains recovered from healthy tomato and their isolation sources. aGPS locality: Teboulba, Monastir, Tunisia (N35°38'38.256"; E10°56'48.458"), Knaeis, Sousse, Tunisia (N35°40'59,999"; E10°31'0,001"), M’saken, Sousse, Tunisia (N35°43'32,073''; E10°34'48,90''), Chott-Mariem, Sousse, Tunisia (N35°43'32.073''; E10°34'48.90'').
Tests of the plant growth-promoting ability on tomato seedlings challenged with pathogen and Fusarium wilt suppressive potential
Healthy tomato seedlings (cv. Rio Grande), at the two true-leaf stage, were carefully removed from alveolus plates and transplanted into individual pots (12.5 cm × 14.5 cm) containing sterilized peat.
Application of whole-cell suspensions of endophytic bacteria
Bacterial strains were applied to tomato seedlings by drenching the substrate with 25 mL of each cell suspension (108 cells/mL) [23]. Inoculation with FOL was performed 6 days post-bacterial treatment by a substrate drench with 25 mL of a conidial suspension (106 conidia/mL) [24].
Application of cell-free filtrates of endophytic bacteria
Cell-free culture filtrates were prepared by centrifugation the liquid culture of bacteria at 9777 g for 10 min followed by microfiltration through a 0.22 μm pore size filter. Each filtrate was applied to seedlings by drenching the substrate with 10 mL for each pot. Six days posttreatment, tomato seedlings were watered each with 10 mL of FOL conidial suspension (~106 conidia/mL).
For both in vivo tests, uninoculated control (NIC) seedlings were treated with equal volumes of sterile distilled water (SDW) only. The positive control (IC) seedlings were inoculated with FOL and treated with SDW. Five replicates of one seedling each were used for each individual treatment. The whole experiment was repeated once. Plants were grown under greenhouse conditions as described above for about 60 days and watered with tap water every two-three days. Growth parameters (plant height, fresh weight of the whole plant and roots’ fresh weight) were measured in all tomato plants at 60 DPI with pathogen. Parameters of Fusarium wilt severity noted 60 days postinoculation (DPI) with FOL are disease severity rate using 0-4 scale [25], the vascular browning extent (from collar) and FOL re-isolation frequency (percentage of pathogen colonization of stem fragments) on PDA.
Effect of endophytic bacteria on Fusarium oxysporum f. sp. lycopersici mycelial growth
Effect of whole-cell suspensions: Twenty μL of whole-cell suspensions of eight bacterial strains (~108 cells/mL) grown in Luria- Bertani (LB) broth medium, were suspended separately into a well performed using a sterile Pasteur pipette (6 mm in diameter, 3 mm in depth) at one side of the Petri plate (90 mm in diameter). An agar plug (6 mm in diameter) removed from the growing edge of a 7 day-old culture of FOL was placed at the opposite side of the plate. Control plates were treated with 20 μL of SDW only [26]. Each individual treatment was replicated three times. The whole experiment was conducted twice. After 6 days of incubation at 25°C, the colony diameter of the pathogen was measured and the mycelial growth inhibition rate was calculated [27].
Effect of cell-free filtrates: Cell-free culture filtrates of eight bacterial strains were prepared separately as described above. LB filtrate was used as control treatment. The antifungal activity of cell-free filtrates was assessed using the poisoned method at the concentration of 20% (v/v) [28]. This concentration was previously shown to be more effective towards FOL mycelial growth than other tested concentrations (data not shown). Cultures were incubated at 25°C for 4 days. Each individual treatment was replicated three times. The whole experiment was conducted twice. The colony diameters of the pathogen were measured and the inhibition rate of the pathogen was calculated.
Effect of endophytic bacteria on Fusarium oxysporum f. sp. lycopersici sporulation ability
Two bacterial strains Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 were selected for use in this test because they were the most effective in reducing Fusarium wilt severity using cell suspensions and filtrates.
FOL sporulation ability was assessed in water conidial suspension of FOL (1% v/v) (~2.7 × 103 conidia/mL) in presence of each bacterial strain cell suspension re-suspended to 1% (v/v) in SDW (~108 cells/ mL). The control was a FOL conidial suspension in SDW (1% v/v). The tubes were shaken for few seconds with a vortex. After incubation for 3 days at 25°C, FOL conidia were counted using a Malassez haemocytometer. Four counts per tube were used as a replicate and three replicates for one separate tube each were used for each individual treatment. The whole experiment was repeated twice.
Sporulation of FOL was expressed as the number of conidia per unit volume (conidia/mL). The percentage of sporulation inhibition was determined [29].
Effects of heating, pH adjustment, and proteinase K treatment on antifungal properties towards Fusarium oxysporum f. sp. lycopersici
Two bacterial strains S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 were selected for use in subsequent tests. To determine the stability of extracellular metabolites produced by the two bacterial strains, cell-free filtrates were either: incubated at 50 or 100°C for 15 min; had the pH adjusted at pH 2 and pH 12 or treated with proteinase K (0.1 mg/mL) at 37°C for 60 min before being used for antifungal bioassays [30,31]. Antifungal activity of filtrates was tested at 20% (v/v) using the poisoned technique method [28]. Control cultures contained LB filtrate only. Each individual treatment was replicated three times. The whole experiment was conducted twice. After incubation at 25°C for 4 days, the diameters of FOL colony was measured and the inhibition rate of the pathogen mycelial growth was calculated [27].
Effect of duration of incubation on antifungal activity towards Fusarium oxysporum f. sp. lycopersici
In order to determine the optimal period of production of antifungal metabolites, cell-free culture filtrates of two bacterial strains, S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43, were used for this test. Each bacterial strain was cultured in LB medium at 28 ± 2°C for 1, 2, 3, 4 and 7 days and under continuous shaking at 150 rpm. The control was LB filtrate. Antifungal activity of cell-free filtrates was assessed at 10% (v/v) as described by Karkachi et al. [28]. Each individual treatment was replicated three times. The whole experiment was conducted twice. FOL colony diameter was measured and the mycelial growth inhibition rate of the pathogen was calculated [27].
In vitro antifungal activity of chloroform and n-butanol extracts towards Fusarium oxysporum f. sp. lycopersici
Organic extraction was performed for the two strains S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 using chloroform and n-butanol [30,32]. Sixty milliliters of cell-free culture filtrate of each strain were poured in a separating funnel and 60 mL of solvent (chloroform or n-butanol) were added carefully. In the end of liquidliquid extraction, the solvent was evaporated in a rotary evaporator at 35°C for chloroform and 75°C for n-butanol with a slight rotation at 150 rpm.
To assess their antifungal activity against FOL, chloroform and nbutanol extracts were suspended in ethanol (1:1) (mg/mL) (w/v) and added separately at two concentrations (2.5 and 5%) (v/v) to molten PDA medium amended with streptomycin sulfate (300 mg/L w/v). Control cultures were treated with similar concentrations of ethanol. Two commercial products namely Bavistin® (50% carbendazim, chemical fungicide) and Bactospeine® (16000 UI/mg, Bacillus thuringiensis -based biopesticide) used at 2.5 and 5% (v/v) each were tested to compare their antifungal activity with the obtained organic extracts. After solidification of the mixture, an agar plug (6 mm in diameter) colonized by FOL, removed from 7-day-old cultures, was placed at the center of each plate. Each individual treatment was replicated three times. The whole experiment was repeated once. After incubation for 7 days at 25°C, FOL colony diameters were measured and the inhibition rate was calculated [27].
Hydrogen cyanide production
Hydrogen cyanide (HCN) was detected qualitatively according to Lorck [33]. Eight bacterial strains were streaked individually on NA medium supplemented with glycine (4.4 g/L) (w/v). Control plates contained glycine-NA medium only for comparison. Treatments were performed in triplicate. Experiment was repeated once. The plates were sealed with parafilm and incubated at 25°C for 4 days. Change in color from yellow to light-reddish brown indicates positive production of HCN by the strain tested.
Enzymatic activity
Eight bacterial strains were assessed for the proteolytic and chitinolytic activities onto sterilized skim milk agar 3% (v/v) medium and minimum-chitin® (MP Biomedicals, LLC, IIIKrich, France)-agar medium (0.5% w/v) according to Tiru et al. [27]. Water bacterial suspensions (~108 cells/mL) were streaked separately on each agar plate. Control plates contained either: skim milk agar and/or chitinagar medium only. Three plates were used for each individual treatment. Each experiment was repeated once. The diameter of the clear zone formed around the bacterial spots was measured after 48 h and/or 72 h of incubation at 28 ± 2°C.
Detection of chitinase (ChiA ) gene and sequence analysis
Two selected endophytic bacteria (S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43), as the most effective in reducing Fusarium wilt severity using whole-cell suspensions and cell-free filtrates, were screened for the presence of chitinase gene (ChiA ). The primers used for ChiA gene are 5'-GATATCGACTGGGAGTTCCC-3' and 5'-CATAGAAGTCGTAGGTCATC-3'. The expected amplicon size was about 225 bp. PCR conditions were 94°C for 4 min, then 35 cycles of 92°C, 58°C, and 72°C for 1 min each, and 72°C for 7 min [34]. Each PCR was conducted at least three times. PCR products were purified using PCR-clean up kit, (Nucleo- Spin®). Purified products were visualized by agarose gel electrophoresis and quantified using a Qubit 2.0 Fluorometer (Life Sciences®), then cloned into the pGEMT-Easy vector (Promega®) according to the manufacturer's instructions. Recombinant clones were screened by PCR (as described above). Plasmids were extracted using a NucleoSpin® Plasmid/Plasmid (NOLid) protocol and sequenced.
Homologies of chitinase nucleotide sequence and the chitinase amino acid sequence of a given recombinant plasmid were performed using BLAST-N and BLAST-X programmes from GenBank database, respectively. Alignment between the target sequence and the closely related ones was performed using the ClustalX (1.81). Phylogenetic trees of amino acid and nucleotides sequences were constructed based on neighbour joining (NJ) method with 1000 bootstrap sampling.
Detection of lipopeptide antibiotics genes
The endophytic bacteria B. subtilis subsp. inaquosorum CT43 was assessed for the presence of lipopeptide genes (LPs) including genes encoding for surfactin, iturin a, fengycin D and Bacillomycin D biosynthesis. Primers used for respective LPs genes are:
1. sfp (5'-ATGAAGATTTACGGAATTTA-3' 5'- TTATAAAAGCTCTTCGTACG-3').
2. ItuD (5'-GATGCGATCTCCTTGGATCGT-3' and 5'- ATCGTCATGTGCTGCTTGAG-3').
3. FenD (5'-TTTGGCAGCAGGAGAAGTTT-3' and 5'- GCTGTCCGTTCTGCTTTTTC-3').
4. BamC (5'-GAAGGACACGGAGAGAGTC-3' and 5'- CGTGATGACTGTTCATGCT-3').
The expected amplicon size was about 675 pb, 647 pb, 964 pb and 875 pb, respectively. Cycling parameters were 5 min at 95°C then 30 cycles of 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, and 72°C for 10 min [35]. Each PCR was conducted at least three times. Purification of PCR product, cloning and sequence analysis of each LPs gene was described as above.
Salicylic acid production
The two selected endophytic bacteria (S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43) were assessed for their ability to produce salicylic acid according to Nagarajkumar et al. method [36]. Bacterial colonies were grown in succinate medium at 28 ± 2°C for 48 h with continuous shaking. The control contained only succinate medium. The absorbance of the iron-salicylic acid complex was measured at 527 nm. One measurement was used as a replicate with three replicates for each treatment. The whole experiment was conducted twice. A standard curve was prepared using salicylic acid dissolved in the succinate medium. The quantity of salicylic acid present in each culture filtrate was expressed as mg/mL.
Siderophore production
Siderophore production was checked qualitatively according to Lacava et al. for the two selected strains [37]. An agar plug (6 mm in diameter) of bacterial colonies, 2-day-old on NA medium was plated onto chrome azurol S (CAS) agar medium. Control cultures contained the CAS agar medium only. Each individual treatment was replicated three times. Experiment was repeated once. After 5 days of incubation at 28 ± 2°C, the yellow halo formed around colonies was measured.
Statistical analysis
Data were subjected to a one-way analysis of variance (ANOVA) using Statistical Package for the Social Sciences (SPSS) software for Windows version 16.0. Each in vitro and/or in vivo experiment was conducted twice yielding similar results. Means were separated using LSD or Duncan Multiple Range tests to identify significant pair-wise differences at P ≤ 0.05. Correlations between Fusarium wilt severity and plant growth parameters were analyzed using bivariate Pearson’s test at P ≤ 0.01.
Promoting growth on pathogen-inoculated tomato plants and suppressing potential of Fusarium wilt disease
Effect of whole cell suspensions: Analysis of variance revealed a significant decrease (at P ≤ 0.05) in fusarium wilt severity, noted on tomato plants 60 DPI with FOL, depending on tested cell bacterial treatments. As shown in Figure 1A and 1B, whole cell suspensions (~108 cells/mL) of all eight tested strains separately had significantly suppressed leaf yellowing and/or necrosis (P=3.34 E-8), by 35.3 to 76.5%, and reduced the extent of vascular browning (P=5.49 E-11) by 69.8 to 84.9%, compared to untreated and FOL-inoculated controls. Interestingly, tomato plants inoculated with FOL and treated with S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 exhibited 76.5% less wilting severity and 83-84.9% lower vascular browning extent compared to FOL-inoculated control. Furthermore, S. maltophilia CT16- and B. subtilis subsp. inaquosorum CT43-based treatments had significantly similar effects as the disease free control (Figure 1A and 1B). FOL re-isolation frequency from internal stem tissues was also reduced by 60-90% in tomato plants treated by wholecell bacterial tested as compared to FOL-inoculated and untreated control plants. The highest decrease in pathogen re-isolation frequency (90%) was achieved using S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 treatments (Figure 1C).
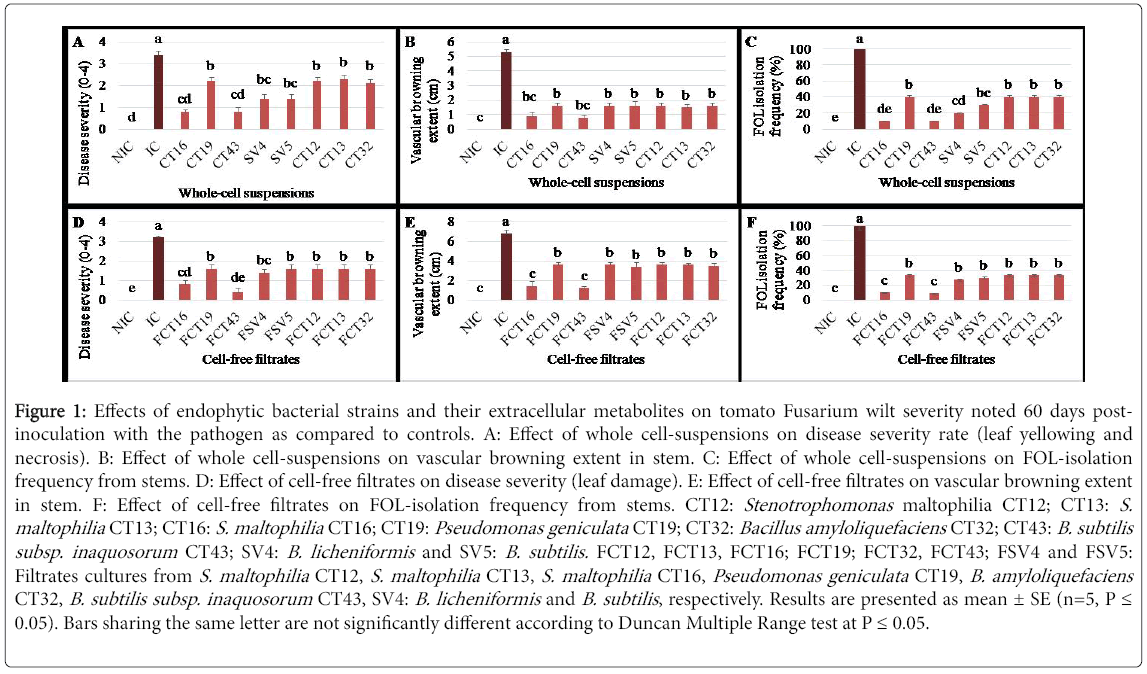
Figure 1: Effects of endophytic bacterial strains and their extracellular metabolites on tomato Fusarium wilt severity noted 60 days postinoculation with the pathogen as compared to controls. A: Effect of whole cell-suspensions on disease severity rate (leaf yellowing and necrosis). B: Effect of whole cell-suspensions on vascular browning extent in stem. C: Effect of whole cell-suspensions on FOL-isolation frequency from stems. D: Effect of cell-free filtrates on disease severity (leaf damage). E: Effect of cell-free filtrates on vascular browning extent in stem. F: Effect of cell-free filtrates on FOL-isolation frequency from stems. CT12: Stenotrophomonas maltophilia CT12; CT13: S. maltophilia CT13; CT16: S. maltophilia CT16; CT19: Pseudomonas geniculata CT19; CT32: Bacillus amyloliquefaciens CT32; CT43: B. subtilis subsp. inaquosorum CT43; SV4: B. licheniformis and SV5: B. subtilis. FCT12, FCT13, FCT16; FCT19; FCT32, FCT43; FSV4 and FSV5: Filtrates cultures from S. maltophilia CT12, S. maltophilia CT13, S. maltophilia CT16, Pseudomonas geniculata CT19, B. amyloliquefaciens CT32, B. subtilis subsp. inaquosorum CT43, SV4: B. licheniformis and B. subtilis , respectively. Results are presented as mean ± SE (n=5, P = 0.05). Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P = 0.05.
Growth parameters of tomato plants (plant height, plant fresh weight and root fresh weight), noted 60 DPI with FOL, varied significantly (at P ≤ 0.05) upon cell-bacterial treatments tested. Plants treated separately with the whole bacterial cells of all eight strains were significantly (P=6.78 E-10) taller by 35.1-41.4% compared to FOLinoculated and untreated control plants. S. maltophilia CT16- and B. subtilis subsp. inaquosorum CT43-based treatments led to a significant increase in plant height by 8.4 to 10.3% relative to free-pathogen controls (Figure 2A). Treatments using whole-cell suspensions of eight strains showed 34.9-44% (P=1.04 E-7) and 74.5-81.1% (P=6.27 E-6) significantly greater plant and root fresh weights than FOL-inoculated and untreated controls, respectively (Figures 2B and 2C). Tomato plants infected with FOL and treated separately with the eight strains exhibited significantly similar plant and root fresh weights as the uninoculated and untreated ones.
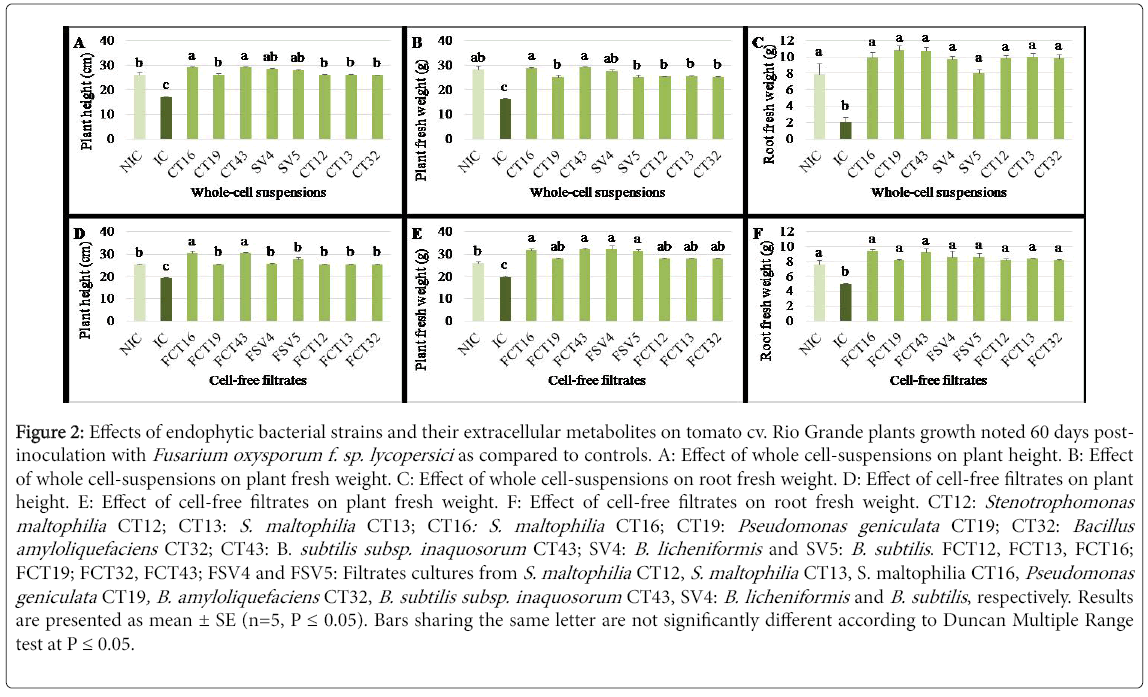
Figure 2: Effects of endophytic bacterial strains and their extracellular metabolites on tomato cv. Rio Grande plants growth noted 60 days postinoculation with Fusarium oxysporum f. sp. lycopersici as compared to controls. A: Effect of whole cell-suspensions on plant height. B: Effect of whole cell-suspensions on plant fresh weight. C: Effect of whole cell-suspensions on root fresh weight. D: Effect of cell-free filtrates on plant height. E: Effect of cell-free filtrates on plant fresh weight. F: Effect of cell-free filtrates on root fresh weight. CT12: Stenotrophomonas maltophilia CT12; CT13: S. maltophilia CT13; CT16: S. maltophilia CT16; CT19: Pseudomonas geniculata CT19; CT32: Bacillus amyloliquefaciens CT32; CT43: B. subtilis subsp. inaquosorum CT43; SV4: B. licheniformis and SV5: B. subtilis . FCT12, FCT13, FCT16; FCT19; FCT32, FCT43; FSV4 and FSV5: Filtrates cultures from S. maltophilia CT12, S. maltophilia CT13, S. maltophilia CT16, Pseudomonas geniculata CT19, B. amyloliquefaciens CT32, B. subtilis subsp. inaquosorum CT43, SV4: B. licheniformis and B. subtilis , respectively. Results are presented as mean ± SE (n=5, P = 0.05). Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P = 0.05.
Effect of cell free filtrates: Fusarium wilt severity, noted on tomato plants 60 DPI with FOL, varied significantly (at P ≤ 0.05) depending on filtrates tested. All cell-free filtrates tested significantly decrease the disease severity rate (yellowing and/or necrosis) by 55 to 87.5% (P=1.71 E-8) and the vascular browning extent by 47 to 82.3% (p=2.6 E-9) compared to FOL-inoculated and untreated control (Figure 1D and 1E). FOL re-isolation frequency from tomato plants challenged with FOL and treated separately with all tested filtrates (Figure 1F) was lowered by 66.6 to 90.9% relative to FOL-inoculated control. Cell-free filtrates from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 have been the most effective in suppressing Fusarium wilt symptoms by 75-87.5% and in reducing FOL colonization in tomato stems by 90-90.9% compared to FOL-inoculated and untreated controls. Furthermore, plants inoculated with FOL and treated with S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 filtrates had significantly similar effects as free disease ones.
Plant height and plant and root fresh weights, noted 60 DPI with FOL, varied significantly (at P ≤ 0.05) upon cell-free filtrates tested. Data shown in Figures 2D, 2E and 2F revealed that significant increase in these three growth parameters was achieved using all filtrate-based treatments. This increase ranged between 24.4-36.8%, 29.3-38.1% and 39-46.8% for plant height (P=2.59 E-8), fresh weight of tomato plants (P=8.81 E-7) and root fresh weight (P=0.002), respectively, as compared to FOL-inoculated and untreated controls. Tomato plants challenged with FOL and treated with filtrates from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 were 17% significantly taller than free-pathogen control (Figure 2D). Except P. geniculata CT19 filtrate-based treatment, the cell-free filtrates tested significantly enhanced the whole plant fresh weight by 17.2 to 18.7% relative to uninoculated and untreated control ones (Figure 2E). Estimated via the root fresh weight, plants treated by all the filtrates tested showed similar growth when compared to uninoculated and untreated control plants (Figure 2F).
Correlation between plant growth parameters and Fusarium wilt severity
Pearson’s correlation analysis revealed that plant height was significantly and negatively correlated to the disease severity rate (r=-0.750; P=0.002) and to the extent of vascular browning (r=-0.750; P=0.002). Furthermore, plant fresh weight was significantly and negatively correlated to the disease severity rate (r=-0.726; P=0.003) and to the extent of vascular browning (r=-0.546; P=0.043). Root fresh weight was also significantly and negatively correlated to disease severity rate (r=-0.604; P= 0.022) and to the vascular browning extent (r=-0.678; P=0.008). FOL re-isolation frequency was significantly and negatively correlated to plant height (r=-0.864; P=6.881 E-5), whole plant fresh weight (r=-0.815; P=3.808 E-4) and root fresh weight (r=-0.771; P=0.001).
Fusarium oxysporum f. sp. lycopersici mycelial growth using whole-cell suspensions and cell-free filtrates of endophytic bacterial strains
Analysis of variance revealed a significant (at P ≤ 0.05) variation in the colony diameter of FOL depending on tested whole-cell suspensions and/or cell-free filtrates of endophytic bacterial strains tested at 20% (v/v). Results given in Figure 3 showed a significant decrease by 47.8-69% (P=2.61 E-8) in FOL mycelial growth using the eight bacterial strains cells and by 17.3-65.6% (P=1.24 E-8) using their cell-free filtrates, noted after 6 and 4 days of incubation at 25°C, respectively as compared to the untreated controls. Tested using whole-cell suspensions, the highest inhibition (69%) was achieved using S. maltophilia CT16 whole cells followed by 58 and 55.3% inhibitions conferred by B. licheniformis SV4 and B. subtlis SV5, whereas lesser inhibition rates (47.8 and 53.5%) were induced by P. geniculata CT19 and B. subtilis subsp. inaquosorum CT43, respectively (Figure 3). The highest growth inhibition 65.6%, was obtained using the extracellular metabolites from B. subtilis subsp. inaquosorum CT43 relative to the untreated control followed by 52.6%, 36.6-42.7% and 17.3% induced respectively by cell-free filtrates from B. licheniformis SV4, S. maltophilia CT16 and B. subtilis SV5 and by P. geniculaata CT19, S. maltophilia CT12, S. maltophilia CT13 and B. amyloliquefaciens CT32 (Figure 3).
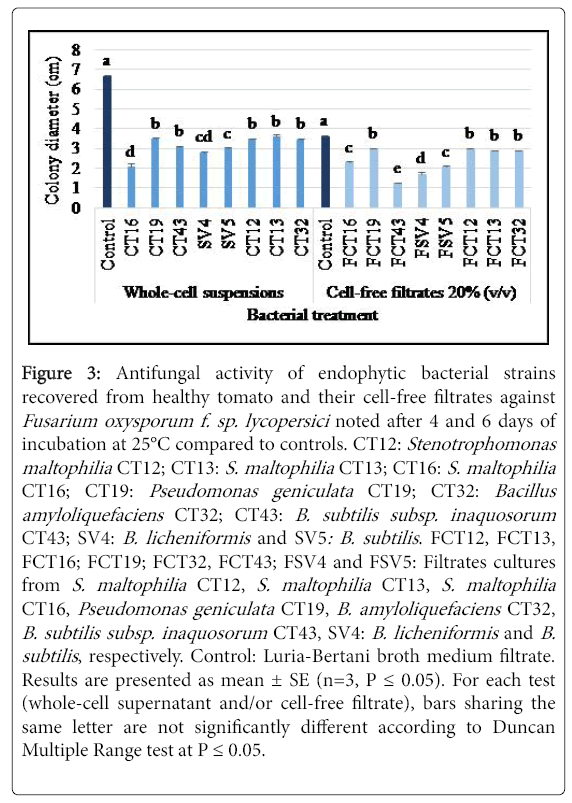
Figure 3: Antifungal activity of endophytic bacterial strains recovered from healthy tomato and their cell-free filtrates against Fusarium oxysporum f. sp. lycopersici noted after 4 and 6 days of incubation at 25°C compared to controls. CT12: Stenotrophomonas maltophilia CT12; CT13: S. maltophilia CT13; CT16: S. maltophilia CT16; CT19: Pseudomonas geniculata CT19; CT32: Bacillus amyloliquefaciens CT32; CT43: B. subtilis subsp. inaquosorum CT43; SV4: B. licheniformis and SV5: B. subtilis . FCT12, FCT13, FCT16; FCT19; FCT32, FCT43; FSV4 and FSV5: Filtrates cultures from S. maltophilia CT12, S. maltophilia CT13, S. maltophilia CT16, Pseudomonas geniculata CT19, B. amyloliquefaciens CT32, B. subtilis subsp. inaquosorum CT43, SV4: B. licheniformis and B. subtilis , respectively. Control: Luria-Bertani broth medium filtrate. Results are presented as mean ± SE (n=3, P = 0.05). For each test (whole-cell supernatant and/or cell-free filtrate), bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P = 0.05.
Fusarium oxysporum f. sp. lycopersici sporulation ability
S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 whole cell suspensions (108 cells/mL) have significantly (P=9.25 E-23) reduced the growth of FOL conidia by 97.2 and 98%, respectively, after three days of incubation at 25°C as compared to the untreated control FOL conidial suspension (2.12 × 103 conidia/mL) (Figure 4).
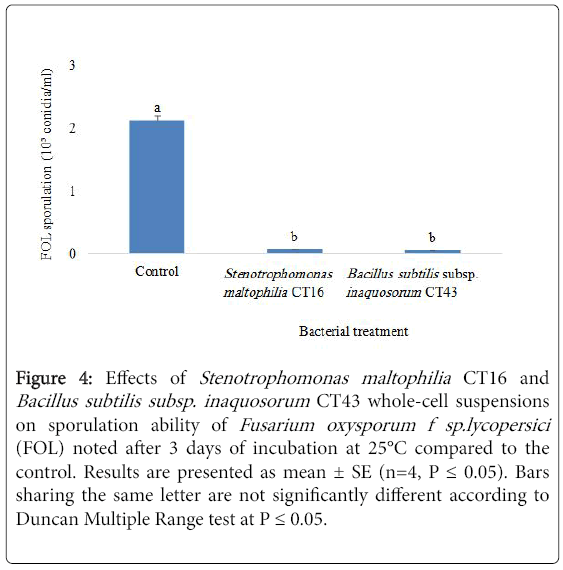
Figure 4: Effects of Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 whole-cell suspensions on sporulation ability of Fusarium oxysporum f sp.lycopersici (FOL) noted after 3 days of incubation at 25°C compared to the control. Results are presented as mean ± SE (n=4, P = 0.05). Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P = 0.05.
Physicochemical treatments affecting the antifungal properties of cell-free filtrates of Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43
Antifungal activity of cell-free culture filtrates from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 toward FOL was assessed depending on several factors tested. Analysis of variance of pathogen colony diameter revealed significant interaction between cell-free filtrates used at 20% (v/v) and factors tested, heating (P=0.004), pH modification (P=2.37 E-6) and proteinase K addition (P=1.45 E-4).
As shown in Table 2, heating the culture filtrates of S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 at 100°C for 15 min led to a significant decline in the antifungal activity of the tested cell-free filtrate toward FOL, where pathogen growth was inhibited by 19.8-24.5% compared to 50.6-53.2% and 50.5-54.1% noted using filtrates heated at 50°C and unheated ones, respectively.
| Factors tested | Cell-free filtrates from endophytic bacterial isolates | ||
|---|---|---|---|
| Duration of bacterial culture incubation (days)a | Control | FCT16 | FCT43 |
| 1 | 3.42 a ± 0.03 | 3.35 a ± 0.01 (2) | 3.18 a ± 0.07 (7) |
| 2 | 4.03 a ± 0.04 | 3.6 b ± 0.09 (10.7) | 3.58 b ± 0.04 (11.2) |
| 3 | 3.62 a ± 0.05 | 2.92 b ± 0.07 (19.3) | 2.72 b ± 0.1 (24.9) |
| 4 | 3.7 a ± 0.03 | 2.63 b ± 0.08 (28.9) | 2.36 c ± 0.01 (36.2) |
| 7 | 3.87 a ± 0.1 | 3.3 b ± 0.01 (14.7) | 3.33 b ± 0.03 (13.9) |
| Heat treatmentb | |||
| Untreated | 3.88 ± 0.04 | 1.92 ± 0.07 (50.5) | 1.78 ± 0.05 (54.1) |
| 50°C, 15 min | 3.85 ± 0.2 | 1.9 ± 0.03 (50.6) | 1.8 ± 0.01 (53.2) |
| 100°C, 15 min | 4.2 ± 0.1 | 3.37 ± 0.03 (19.8) | 3.17 ± 0.07 (24.5) |
| pH modificationc | |||
| Untreated | 3.92 ± 0.1 | 2.03 ± 0.03 (48.2) | 1.88 ± 0.03 (52) |
| pH 2 | 1.63 ± 0.05 | 1.13 ± 0.02 (30.7) | 1.07 ± 0.02 (34.3) |
| pH 12 | 3.57 ± 0.07 | 2.45 ± 0.03 (31.4) | 2.35 ± 0.01 (34.2) |
| Enzymatic degradationd | |||
| Untreated | 3.87 ± 0.02 | 1.92 ± 0.05 (50.4) | 1.88 ± 0.03 (51.4) |
| Proteinase K, 37°C, 60 min | 3.85 ± 0.09 | 2.88 ± 0.05 (25.2) | 2.55 ± 0.03 (33.8) |
Table 2: Characterization of the antifungal activity of cell-free filtrates of Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 against Fusarium oxysporum f. sp. lycopersici (FOL) mycelial growth as compared to controls. xAntifungal activity noted after 4 days of incubation at 25°C and evaluated as mean diameter of FOL colonies (cm); ± (Standard error). For each line, numbers in parenthesis indicate the percentage (in%) of FOL mycelial growth inhibition as compared to control. FCT16, FCT43: Cell-free filtrates from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43, respectively. Control: Luria-Bertani broth medium filtrate. aCell-free filtrates tested at 10% (v/v). For each incubation duration, values followed by the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05. b, c, d Cell-free filtrates tested at 20% (v/v). b LSD (Bacterial treatment × Heat treatment): 0.36 cm at P ≤ 0.05. c LSD (Bacterial treatment × pH modification): 0.21 cm at p ≤ 0.05. d LSD (Bacterial treatment × Enzymatic degradation): 0.59 cm at P ≤ 0.05.
Adjustment of the pH of cell-free filtrates of S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 to pH2 and pH 12 had reduced the antifungal potential of the filtrates where FOL growth was inhibited by 30.7-34.3% and 31.4-34.2%, respectively, compared to 48.2-52% recorded with the non-adjusted pH 6.4 of control filtrates (Table 2).
The protease K treatment led to a significant decrease in the antifungal activity of cell-free filtrate of both strains tested against FOL. Pathogen growth decreased by 25.2-33.8% with proteinase Ktreated filtrates compared to 50.4-51.4% noted using untreated ones (Table 2).
Optimization of the incubation duration of Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 cultures for the production of antifungal metabolites
Analysis of variance revealed a significant (at P ≤ 0.05) variation in the diameter of pathogen colonies treated with the cell-free filtrates of S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 tested at 10% (v/v) issued from 2-, 3-, 4-, and 7- day-old cultures in LB medium at 28 ± 2°C. Results given in Table 2 revealed that all filtrates issued from 1 day-old cultures did not significantly decrease (2 and 7%) FOL mycelial growth. The highest significant FOL growth inhibition by 28.9 and 36.2% (P=2.78 E-7), was achieved with the cell-free filtrates extracted from 4 days-old cultures of S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43, compared to 19.3-24.9% (P=0.003), 14.7-13.9% (P=0.05), and 10.7-11.2% (P=0.01) noted at 3, 7 and 2 days of incubation, respectively.
Assessment of antifungal potential of organic extracts from Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 towards Fusarium oxysporum f. sp. lycopersici
Analysis of variance revealed a significant (at P ≤ 0.05) variation in FOL colony diameter depending on organic extracts (chloroform and n-butanol extracts) tested and concentrations used, and the existence of a significant interaction between both factors. Chloroform and nbutanol extracts from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43, used at 1 mg/mL (w/v), inhibited FOL growth by 17.5 to 73.9% as compared to the ethanol controls whatever the concentration used. Except chloroform extracts from both strains used at 2.5% (v/v), the decrease in FOL growth was higher with the remaining extract-based treatments as compared to Bavistin® (31.3-39.5%) and Bactospeine® (40.9-43.2%) whatever the concentration used (Table 3).
| Solvent/ Concentration (% v/v) | Control | Organic extract from filtrates of endophytic bacterial isolates | Commercial products | ||
|---|---|---|---|---|---|
| Ethanol | ECT16 | ECT43 | F | Bio-F | |
| Chloroform | |||||
| 2.5 | 7.25 a ± 0 | 5.98 b (17.5) | 5.53 c (23.7) | 4.98 d (31.3) | 4.28 e (40.9) |
| 5 | 7.16 a | 3.16 c (55.8) | 2.41 d (66.3) | 4.33 b (39.5) | 4 b (43.2) |
| n-Butanol | |||||
| 2.5 | 7.25 a | 5.05 b (30.3) | 4.36 c (39.8) | 4.98 b (31.3) | 4.28 c (40.9) |
| 5 | 7.16 a | 2.41 c (66.3) | 1.86 c (73.9) | 4.33 b (39.5) | 4 b (43.2) |
Table 3: Effect of chloroform and n-butanol extracts from Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43 tested at two concentrations against Fusarium oxysporum f. sp. lycopersici (FOL) noted after 7 days of incubation at 25°C as compared to controls. ECT16, ECT43: Organic extract from Stenotrophomonas maltophilia CT16 and Bacillus subtilis subsp. inaquosorum CT43, respectively. Control: Ethanol. F: Bavistin® (Chemical fungicide, carbendazim); Bio-F: Bactospeine® (Bacillus thuringiensis-based biopesticide). LSD (Treatments tested × Concentrations used): 0.56 cm at P ≤ 0.05. For each line, values followed by the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05. Numbers in parenthesis indicate the percentage (in%) of FOL mycelial growth inhibition as compared to control.
All organic extracts tested were found to be more active when used at 5% than at 2.5% (v/v). In fact, chloroform extracts from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43 decreased FOL growth by 55.8 and 36.3% when applied at 5% (v/v), compared to 17.5 and 23.7% recorded at 2.5% (v/v), respectively. In addition, when applied at 5% (v/v), n-butanol extracts from S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43, inhibited the pathogen growth by 66.3 and 73.9% compared to 30.3 and 39.8% when tested at 2.5% (v/v), respectively (Table 3).
Whatever the concentration used, the highest inhibition of FOL growth 30.3-73.9% was achieved with n-butanol extracts from both strains compared to 17.5-66.3% recorded with chloroform extracts (Table 3). Metabolites extracted from B. subtilis subsp. inaquosorum CT43 were found to be more active than those extracted from S. maltophilia CT16 whatever the organic solvent used and the concentration applied where the decrease of pathogen colony diameter significantly varied from 23.7-73.9 versus 17.5-66.3%, respectively, as compared to the ethanol controls (Table 3).
Assessment of antifungal mechanisms deployed by endophytic bacteria from Solanum lycopersicum
Hydrogen cyanide production: The eight bacterial strains tested were qualitatively assessed for the production of volatile antibiotic, HCN on NA medium amended with glycine. Only S. maltophilia CT16 was able to produce this antibiotic (Table 4).
| Endophytic bacterial isolates | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene | CT12 | CT13 | CT16 | CT19 | CT32 | CT43 | SV4 | SV5 |
| Chita | + | + | +* | + | - | - | + | + |
| Protb | + | + | - | + | + | + | + | + |
| HCNc | - | - | + | - | - | - | - | - |
| Lpsd | n.t | n.t | n.t | n.t | n.t | + | n.t | n.t |
| SAe | n.t | n.t | + | n.t | + | n.t | n.t | n.t |
| Sdf | n.t | n.t | + | n.t | + | n.t | n.t | n.t |
Table 4: Antifungal properties of endophytic bacterial isolates recovered from Solanum lycopersicum (L.) CT12, CT13, CT16: Stenotrophomonas maltophilia , CT19: Pseudomonas geniculata, CT32: Bacillus amyloliquefaciens, CT43: B. subtilis subsp. inaquosorum, SV4: B. licheniformis , SV5: B. subtilis . aChitinase activity: Tested on chitinagar (0.5% w/v) medium and incubated at 28 ± 2°C for 72 h; +: Presence of clear zone; -: Absence of clear zone; *: Detection of ChiA gene by PCR using 5'-GATATCGACTGGGAGTTCCC-3 'and 5'- CATAGAAGTCGTAGGTCATC-3’ primers. bProtease activity: Tested on skim milk agar (3% v/v) medium and incubated at 28 ± 2°C for 48 h; +: Presence of clear zone; -: Absence of clear zone. cHydrogen cyanide production on glycine-agar (4.4 g/L w/v) medium and incubated at 25°C for 4 days; +: Modification on the filter paper color (light-reddish color); -: No modification on the filter paper color (yellow). dLipopeptide antibiotics gene: Detection of Sfp gene by PCR using 5’-ATGAAGATTTACGGAATTTA-3’ and 5’- TTATAAAAGCTCTTCGTACG-3’ primers; n.t: Not tested. eSalicylic acid production after 48 h incubation at 28 ± 2°C in succinate medium; +: Production of salicylic acid. n.t : Not tested. fSiderophore production: Tested on Chrome Azurol Sulphonate (CAS) agar medium and incubated at 28 ± 2 °C for 5 days; +: Presence of zone of siderophore activity (yellow color); n.t: Not tested.
Siderophores production: B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16 were found able to produce siderophores in CAS agar medium (Table 4) as indicated by the presence of activity zones (yellow color) of about 11 and 13.5 mm in diameter around their colonies, respectively (Supplementary Table 1).
Enzymatic activity: protease activity S. maltiophilia CT12, S. maltiophilia CT13, P. geniculata CT19, B. amyloliquefaciens CT32, B. subtilis subsp. inaquosorum CT43, B. licheniformis SV4 and B. subtilis SV5 were found able to produce protease on skim milk agar medium while S. maltophilia CT16 did not (Table 4).
Chitinase activity and detection of ChiA gene and sequence analysis
Except B. subtilis subsp. inaquosorum CT43 and B. amyloliquefacines CT32, the six remaining strains formed clear zones around their colonies when grown on chitin-agar medium. This indicates that S. maltophilia CT12, S. maltophilia CT13, S. maltophilia CT16, P. geniculata CT19, B. licheniformis SV4 and B. subtilis SV5 are able to produce chitinase (Table 4).
Only the two most active strains in suppressing Fusarium wilt disease (S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43) were selected for detection of chitinase (ChiA ) gene by PCR. S. maltophilia CT16 gave a 225 bp product specific for the ChiA gene using the two tested primers. However, no PCR product was detected in B. subtilis subsp. inaquosorum CT43. The PCR results confirm those obtained with the qualitative test in chitin-agar medium. PCR product for ChiA gene was cloned and sequenced. Two ChiA sequences (CT16_1 and CT16_2) were submitted to Gen Bank and have the following accession numbers: KX087371 and KX087372, respectively. Phylogenetic tree inferred using chitinase nucleotide sequences with closely related sequences revealed a short distance with ChiA gene of Serratia marcescens Bn10 (GenBank accession number DQ165083) and S. marcescens AUDS227 (HQ699804) (Figure 5A). Phylogeny analysis of chitinase amino acid sequences data revealed a short distance with chitinase of S. marcescens (AAZ86539 and ACE78180), Serratia sp. FS14 (WP_044030235), S. marcescens subsp. marcescens (KPQ73123) and uncultured bacterium (CAH23691) (Figure 5B).

Figure 5: Neighbor-joining phylogenetic trees of chitinase nucleotide (A) and amino acid (B) partial sequences obtained from Stenotrophomonas maltophilia CT16 with respect to closely related sequences available in GenBank. Sequences obtained from Genbank database under the following accession numbers: A. AF059494 (Aeromonas hydrophila chitinase A (ChiA ) gene), DQ013365 (Enterobacter sp . NRG-4 chitinase (ChiA ) gene), DQ282126 (Sanguibacter sp. C4 chitinase (Chit58 ) gene), EF451957 (Serratia proteamaculans 18A1 endochitinase (ChiA ) gene), HQ699804 (S. marcescens AUDS227 chitinase (ChiA ) gene), DQ165083 (S. marcescens Bn10 endochitinase (ChiA ) gene), EF151930 (Serratia sp. KCK (ChiA ) gene), AJ812551 (Uncultured bacterium D5 chitinase); B. ACH89423 (chitinase, Aeromonas hydrophila ), AAZ86539 (endochitinase, S. marcescens), WP_046374766 (chitinase, Serratia liquefaciens), WP_020453664 (chitinase A, Serratia plymuthica ), WP_044030235 (chitinase, Serratia sp . FS14), ACE78180 (chitinase A, S. marcescens ), KPQ73123 (chitinase, S. marcescens subsp. marcescens ), CAH23691 (chitinase, uncultured bacterium), and for the chitinase of S. maltophilia CT16 tested (CT16_1 and CT16_2; KX087371 and KX087372, respectively). The tree topology was constructed using ClustalX (1.81).
Only the two most active strains in suppressing Fusarium wilt disease (S. maltophilia CT16 and B. subtilis subsp. inaquosorum CT43) were selected for detection of chitinase (ChiA ) gene by PCR. S. maltophilia CT16 gave a 225 bp product specific for the ChiA gene using the two tested primers. However, no PCR product was detected in B. subtilis subsp. inaquosorum CT43. The PCR results confirm those obtained with the qualitative test in chitin-agar medium. PCR product for ChiA gene was cloned and sequenced. Two ChiA sequences (CT16_1 and CT16_2) were submitted to Gen Bank and have the following accession numbers: KX087371 and KX087372, respectively. Phylogenetic tree inferred using chitinase nucleotide sequences with closely related sequences revealed a short distance with ChiA gene of Serratia marcescens Bn10 (GenBank accession number DQ165083) and S. marcescens AUDS227 (HQ699804) (Figure 5A). Phylogeny analysis of chitinase amino acid sequences data revealed a short distance with chitinase of S. marcescens (AAZ86539 and ACE78180), Serratia sp. FS14 (WP_044030235), S. marcescens subsp. marcescens (KPQ73123) and uncultured bacterium (CAH23691) (Figure 5B).
Lipopeptide antibiotics genes detection and sequence analysis
Detection of genes encoding for LPs antibiotics was assessed by PCR in B. subtilis subsp. inaquosorum CT43 genome. LPs genes were generally present in the genus of Bacillus. In this case, the most active strain of Bacillus spp. in reducing Fusarium wilt severity was selected for this test. B. subtilis subsp. inaquosorum CT43 yielded a 675 bp PCR product specific to the surfactin (Sfp) gene. The iturin D (ItuD ), bacillomycin D (BamD ) and fengycin D (FenD ) genes were not detected in the Bacillus strain tested.
PCR product for Sfp gene was cloned and sequenced to verify product was indeed specific for the expected gene. Two Sfp sequences (CT43_1 and CT43_2) were submitted to GenBank and have the following accessions numbers: KX099383 and KX099384, respectively. Phylogeny analysis of surfactin nucleotide (Figure 6A) and amino acid sequences data (Figure 6B) revealed a short distance with Sfp gene of B. subtilis 96-41 (Genbank accession number EU882341) and surfactin biosynthesis protein of B. subtilis (AIY62581), respectively.

Figure 6: Neighbor-joining phylogenetic trees of surfactin (Sfp) nucleotide (A) and amino acid (B) partial sequences obtained from Bacillus subtilis subsp. inaquosorum CT43 with respect to closely related sequences available in GenBank. Sequences obtained from Genbank database under the following accession numbers: A. KJ652452 (Sfp gene, B. amyloliquefaciens VB8); JQ966544 (Sfp gene, B. axarquiensis GS3; KC846090 (Sfp gene, B. licheniformis NIOT-ARMKVK06); AY185905 (Sfp -like gene, B. megaterium PV361); JQ966542 (Sfp gene, B. mojavensis GS1); AY185904 (Sfp - like gene, B. sphaericus ATCC 14577); HQ711611 (Sfp -like gene, B. subtilis EPC8); HQ711610 (Sfp gene, B. subtilis EPC5); EU146075 (Sfp gene, B. subtilis subsp. subtilis NCIB 3610); EU882341 (Sfp gene, B. subtilis 96-41); JX025778 (Sfp gene, B. tequilensis NIOS11); B. ADZ23658 (B._subtilis _4: surfactin, B. subtilis ), CUB44860 (B._subtilis _3: Sfp B. subtilis ), AAO74611 (Sfp -like, B. megaterium ), ABW74629 (Sfp , B. subtilis subsp. subtilis ), AIY62581 (surfactin biosynthesis protein, B. subtilis ), WP_015715234 (Sfp , B. subtilis group ), AEK64474 (B. subtilis _2: B. subtilis ), WP_049141355 (B. amyloliquefaciens _1: B. amyloliquefaciens ), ACF76869 (B._subtilis _1: biosurfactant protein, B. subtilis ), WP_024713950 (Sfp , B. tequilensis ), ACG68436 (Sfp , B. amyloliquefaciens ), and for the surfactin of B. subtilis subsp. inaquosorum CT43 tested (CT43_1 and CT43_2; KX099383 and KX099384, respectively). The tree topology was constructed using ClustalX (1.81).
Salicylic acid production
Salicylic acid production, inducers of systemic plant resistance, was assessed for the two best active strain B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16 were found able to produce salicylic acid after 48 h of growth in succinate medium (Table 4). Salicylic acid content produced in cultures was of about 3.74 and 2.21 μg/mL, respectively (Supplementary Table 1).
This study highlighted the effectiveness of eight endophytic bacteria namely S. maltophilia CT12 (KR818058), S. maltophilia CT13 (KR818059), S. maltophilia CT16 (KR818060), P. geniculata CT19 (KR818061), B. amyloliquefaciens CT32 (KR818062), B. subtilis subsp. inaquosorum CT43 (KR818063), B. licheniformis SV4 (KR818064) and B. subtilis SV5 (KR818065) associated to healthy tomato plants in suppressing Fusarium wilt and promoting growth in plants challenged with the pathogen.
Assessed for their ability to suppress Fusarium wilt disease, the eight bacterial strains and their extracellular metabolites have successfully decreased yellowing and wilting in FOL-inoculated plants and reduced pathogen mycelial growth. B. subtilis subsp. inaquosroum CT43 and S. maltophilia CT16 have been shown most active in decreasing disease severity using both whole cell suspensions and cell-free culture filtrates. This inhibitory effect against FOL is due in part to the production of bio-active metabolites. Diffusible and/or volatile compounds secreted by endophytic Bacillus spp ., recovered from wild Solanaceae species, exhibited an antifungal activity towards FOL [38]. Metabolites from endophytic B. megaterium , Burkholderia cepacia, P. chlororaphis , P. putida and Serratia marcesens , recovered from healthy tomato plants have shown able to reduce FOL and F. oxysporum f. sp. radicis-lycopersici mycelial growth [17]. Furthermore, FOL sporulation ability has been significantly decreased using whole-cell suspension of B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16. Various secondary metabolites synthesized by antagonistic bacteria and/or fungi decrease the formation and germination of fungal spores [20]. Antimicrobial metabolites produced by S. maltophilia UPMKB9, have been shown to inhibit spores germination of F. oxysporum and Collectotrichum gloeosporioides and to induce hyphal alteration [39].
Our findings clearly demonstrated that the two bacterial strains tested, B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16, produced the maximum of their naturally bioactive metabolites against FOL growth at 4 days of incubation. This is in accordance with Aydi Ben Abdallah results using Bacillus species from wild Solanaceae for FOL biocontrol [38]. In this way, organic extracts (chloroform and nbutanol) obtained from 4-day old cultures of these two bacteria were efficient in reducing pathogen growth compared to the ethanol controls. This growth suppressive effect of chloroform and n-butanol extracts against FOL was previously reported from Bacillus spp . and Serratia sp . C4 recovered from Datura metel , Solanum nigrum , S. elaeagnifolium , N. glauca and C. nocturnum [38,40]. Our study indicated also that the antifungal activity displayed by B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16 decreased due to heating at 100°C, filtrate pH adjustment at pH 2 and pH 12 and proteinase K treatment as reported in many other studies [41,42]. This antifungal activity may, thus, be due in part to heat labile proteins such as extracellular hydrolytic enzymes and in other part to chemical compounds and/or peptide antibiotics.
In fact, synthesis of hydrolytic enzymes, such as chitinase, glucanase, cellulose, pectinase and protease, has been reported for various genera of endophytic bacteria including Stenotrophomonas , Bacillus , Paenibacillus , Erwinia , Pseudomonas and Serratia [19,43]. Therefore, in the current study, we have focused on searching cell-wall degrading enzymes and/or antibiotics used by the endophytic bacteria tested to exert their antifungal activity. Indeed, chitinase gene (ChiA ) was detected in S. maltophilia CT16 but not in B. subtilis subsp. inaquosorum CT43 strain. This lack of ChiA gene for the later strain was also confirmed by negative chitinolytic activity on agar plate. In the same sense, the ChiA gene, encoding for chitinase, have been detected in S. maltophilia MUJ, recovered from wheat rhizosphere and exhibited antifungal activity against Alternaria alternata , A. radicina , Rhizoctonia solani , F. solani , F. oxysporum , F. avenaceum and B. cinerea [44]. Kalai-Grami et al. [5] showed also that endophytic Bacillus spp. (B. velezensis , B. mojavensis , B. amylolequifaciens and B. methylotrophicus ) are unable to produce chitinase on chitin-agar medium. Conversely, ChiA gene was expressed in endophytic Bacillus spp . (B. tequilensis SV39, B. methylotrophicus SV44, B. amylolequifaciens subsp. plantarum SV65, B. tequilensis SV104, B. subtilis SV41 and B. cereus S42) using the same primers [42,45]. Thus, the absence of chitinolytic activity in B. subtilis subsp. inaquosorum CT43, despite its potential ability to suppress Fusarium wilt using both whole cells and filtrates, may be explained by its production of lipopeptide antibiotics and/or other bioactive secondary metabolites belonging to phthalic acid families [46,47]. In this context, B. subtilis subsp. inaquosorum CT43 was assessed for the presence of LPs genes including surfactin (Sfp), bacillomycin (Bam C), iturun (Itu D ) and fengycin (Fen D ) genes and only Sfp gene encoding for surfactin was detected. In Aydi Ben Abdallah et al. previous studies, two strains of B. tequilensis , SV39 and SV104, recovered respectively from D. metel and S. elaeagnifolium , were found to be positive for Sfp gene [42]. However, the strain SV41 of B. subtilis obtained from D. metel stem did not arbor any LP s genes using the same primers. Surfactin, detected in the culture filtrate of endophytic B. subtilis EPC016, recovered from cotton plants, exhibited an important role in Fusarium wilt suppression [10]. In our study, S. maltophilia CT16 has not been assessed for the presence of LPs gene because lipopeptide antibiotics are commonly known only for Bacillus genus. Indeed, when tested for their ability to produce the volatile antibiotic, HCN, only S. maltophilia CT16 strain was shown positive. This secondary metabolite commonly produced by Gram-negative bacteria was shown to be active against several soilborne pathogens such as Sclerotium rolfsii and R. solani [36,48-50]. The production of HCN seemed to vary depending upon strains as shown for S. maltophilia CT12 and CT13 that are unable to produce HCN.
Furthermore, bacterial strains tested are not only active in suppressing disease symptoms and/or in inhibiting the pathogen growth but were also shown effective in enhancing tomato growth. Pearson’s correlation analysis indicated that the reduction of Fusarium wilt severity was related to the decrease in pathogen colonization of vascular tissues, leading to promotion of plant growth. Similarly, Fusarium wilt-suppressive effects displayed by an endophytic bacterium B. subtilis EPC016, isolated from cotton plants, lead to an increase in plant growth and fruit yield of tomato compared to control [10]. In the same sense, Algam et al. found that endophytic Brevibacillus brevis B2 and B. subtilis strains, originally isolated from tomato rhizosphere, had enhanced growth of tomato and had successfully controlled the bacterial wilt disease caused by Ralstonia solanacearum [51]. In Aydi Ben Abdallah et al. findings, endophytic bacteria B. mojavenis S40, S. maltophilia S37, Stenotrophomonas sp. S33, Pseudomonas sp. S85, obtained from surface-sterilized tissues of Datura spp. plants, were shown able to suppress Fusarium wilt disease and to enhance tomato growth in plants challenged or not with FOL [52,53].
Indirectly, plant growth-promoting bacteria enhance the plant growth by reducing the disease severity via the inhibition of pathogens progress and/or by inducing systemic resistance [54]. In the current study, the two most active strains, B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16, in reducing Fusarium wilt severity and enhancing growth of FOL-inoculated plants have shown able to produce salicylic acid and/or siderophores. In fact, among the most commonly tested chemical elicitors for inducing systemic resistance and/or expressing local resistance, salicylic acid play an important role in both effects [55]. In Aydi Ben Abdallah et al. [42] study, the five bioactive Bacillus spp. in reducing tomato Fusarium wilt severity were found able to produce salicylic acid and the highest production was recorded with B. subtilis SV41. Furthermore, application of salicylic acid in various crops successfully reduce severity of Fusarium wilt in tomato, chickpea, asparagus, and crown and root rot of tomato and Verticillium wilt of eggplant [56-59]. Indeed, siderophores production ability has been involved in antagonism through competition for iron with plant pathogenic agents and/or stimulation of plant growth through iron supply [46,60]. Endophytic Bacillus sp ., B. cereus , B. pumilus , B. licheniformis , B. megaterium and S. marcescens , obtained from tomato plants, has been demonstrated by Amaresan et al. as siderophore-producing agents. Kumar and Audipudi reported that S. maltophilia , isolated from chilli pepper rhizosphere, was able to produce siderophores [61,62].
Healthy tomato plants were found to be natural potential sources for isolation of plant growth-promoting bacteria and biocontrol agents. The whole-cell suspensions and cell-free filtrates of the eight endophytic bacteria successfully decrease Fusarium wilt severity, enhance tomato growth in challenged pathogen plants, and inhibit FOL mycelial growth and sporulation. The most bio-active bacteria are B. subtilis subsp. inaquosorum CT43 (KR818063) and S. maltophilia CT16 (KR818061). The decline recorded in the in vitro antifungal activity displayed by their filtrates, treated with proteinase K or heated at 100°C, was explained by the expression of chitinase among other hydrolytic enzymes and/or other heat-labile compounds that need to be more elucidated. Chloroform and n-butanol extracts from the two strains confirm the presence of bio-active metabolites towards FOL that need to be identified using GC-MS or HPLC. B. subtilis subsp. inaquosorum CT43 and S. maltophilia CT16 were found able to produce siderophores and salicylic acid. S. maltophilia CT16 was shown to be HCN-producing agent. B. subtilis subsp. inaquosorum CT43 was found to be new source of antifungal metabolites especially surfactin that can be act as elicitor for inducing systemic resistance of plant.
This work was funded by the Ministry of Higher Education and Scientific Research of Tunisia through the funding allocated to the research unit UR13AGR09-Integrated Horticultural Production in the Tunisian Centre-East, Regional Research Centre on Horticulture and Organic Agriculture of Chott-Mariem, Tunisia. Financial support was also provided by the Postgraduate Programmes 3439 and 3817 (DBB) to KKP. We are kindly grateful to all the team of the Department of Biochemistry and Biotechnology, University of Thessaly, Larissa, Greece for their warm welcome and pleasant working conditions.