Journal of Agricultural Science and Food Research
Open Access
ISSN: 2593-9173
ISSN: 2593-9173
Research Article - (2011) Volume 2, Issue 2
Botanicals are the most promising source and under extensive trials for their biological activity against various pests. The strong inhibition of acetylcholinesterase (AChE: EC 3.1.1.7) is the principal underlying mechanism of action of most of the potential insecticides. Keeping these facts in view, the present study investigated the toxic effects of ethanol extracts of seeds of Annona squamosa (Annonaceae) and Calotropis procera (Asclepiadaceae) in different developmental stages of Musca domestica. L. (Diptera:Muscidae) as well as on the activity of AChE. The third instar larvae of housefly were treated with the different concentrations of all the four extracts by dipping method for 48 h. The LC50 values of the extracts of C. procera and A. squamosa seeds were found to be 870 and 345mgl-1, respectively. The insects were exposed to 5 and 10% concentrations of the LC50 value of each extract along with their control sets to evaluate their inhibitory potential against the activity of AChE. The exposure of these extracts caused significant inhibition of the AChE activity in the developing phases of the insect. The high concentration (10%) of extract from the seeds of A. squamosa exhibited maximum inhibitory effects (56%) on the AChE activity from all the three developmental stages of the insect tested. The results suggest that these plants may prove to be the probable candidates for the development of bioinsecticides to control the population of M. domestica as the safer, ecofriendly and economic alternatives to the synthetic insecticides.
<Keywords: LC50; AchE; Annona squamosa; Calotropis procera; Musca domestica; Bioinsecticide
In recent years, the application of several medicinal plants products has drawn much attention as effective alternatives to the synthetic pesticides and chemical fertilizers. These plant products are reported to be more effective, less expensive, biodegradable and safe for mankind and environment, than their synthetic counterparts, which are environmentally persistent and toxic to non-target organisms including humans eliciting many unidentified diseases after bioaccumulation [1,2]. Therefore, alternatives to conventional pesticides are required to be developed from the active ingredients of plant origin. These compounds have been shown to affect insect populations by reducing their developmental, survival and reproductive rate [3,4]. Several plants belonging to different families and containing a number of phytochemicals such as saponins, tannins, alkaloids, di- and triterpenoids etc., exhibit high pesticidal activities. These molecules have been in use in freshwater bodies to control harmful snails and disease causing insects, such as mosquito larvae and weed fishes [5-9]. The deleterious effect of plant extracts or their pure compounds on insects can be manifested in various ways including toxicity, mortality, antecedent, growth inhibitor, suppression of reproductive behavior and fertility as well as reduction of fecundity [10].
The common housefly, Musca domestica, being the vector of several pathogens such as virus, bacteria, protozoa, helminth eggs etc. is reported to be a menace to the human beings and agronomic livestocks [11]. Different reports suggest that due to prolonged exposure to chemical insecticides worldwide, houseflies have developed resistance against spinasod [12], diflubenzuron [13] and other synthetic insecticides [14,15]. Considering the increasing biomagnifications of these chemical pesticides at each tropic level in the target and non target organisms, it is pertinent to develop ecofriendly and biodegradable insecticides. Products obtained from certain medicinal plants can be used without risk to nontarget organisms. Moreover, consumption of extracts from some of the medicinal plants is even beneficial for human beings [16].
Calotropis procera is a member of the plant family Asclepiadaceae, a shrub widely distributed in West Africa, Asia and other parts of the tropics [17]. The plant is erect, tall, large, much branched and perennial with milky latex throughout. A large quantity of latex can be easily collected from its green parts [17]. The aspect of this natural secretion resembles that of rubber tree, Havea brasiliensis. Local people use it successfully to combat some cutaneous fungal infections. The abundance of latex in the green parts of the plant reinforces the idea that it produced and accumulated latex as a defense strategy against organisms such as virus, fungi and insects [18]. The presence of plant defense related proteins such as hevein, an alpha-amylase inhibitor, has been described to occur in the latex secretion of other plants [19]. However, there are no reports to indicate that Calotropis seed extract may also exhibit insecticidal properties against housefly.
Annonaceae (Custard-apple family) is a large family of almost exclusively tropical trees and shrubs comprising about 130 genera and 2300 species. Some plants of this family have been used traditionally as insecticides. For example, the powdered seeds and leaf juices of Annona spp. are used to kill head and body lice, and the bark extract of Goniothalamus macrophyllus is used as mosquito repellents. Annonaceous acetogenins extracted from the tree leaves, bark and seeds have pesticidal and/or insect antifeedant properties [20]. In addition, the insecticidal activity of Annona squamosa L. seed extracts against the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) has been reported [21]. However, there is no report to indicate that A. squamosa seed extract exhibits insecticidal activity against housefly by acting as an antiAChE agent.
Acetylcholinesterase (AChE), a serine hydrolase, catalyzes the breakdown of the neurotransmitter acetylcholine (ACh) into acetate and choline. This process involves the formation of a substrate-enzyme complex, followed by acetylation of the hydroxyl group of the amino acid serine, present within the esteratic site which is finally deacetylated [22]. It facilitates nerve impulse transmission in the organisms. Its inhibition leads to paralysis and death. However, information on the effects of C. procera and A. squamosa on AChE activity in the developmental stages of housefly, M. domestica is inadequate. Hence, an attempt has been made in the present communication to assess the impact of the extracts of leaves and seeds of these two plant species on the AChE activity in three different developmental stages (larva, pupa and adult) of the insect, M. domestica.
Experimental insect
M. domestica was taken as experimental insect because it is a medically important fly due to its role as a vector of several pathogens.
Rearing technique
Adult house flies were collected from near by areas with the help of a sweep net and reared in the laboratory by the method described earlier [23].
Collection and processing of plant samples
The seeds of C. procera and A. quamosa were collected from the Botanical Garden of University of Allahabad. Seeds were properly cleaned and shade dried for 15-20 days at 32-35°C and relative humidity 50-60%. They were powdered mechanically using commercial electrical stainless steel blender (Remi Anupam Mixie Ltd., India). The samples were stored in air tight container at room temperature in dark for further analysis.
Extraction of plant extracts
The dried seeds were extracted with 1L of 90% ethanol in a soxhlet apparatus (Borosil, India). The extracts were concentrated at 50°C and the residue obtained was stored at 4°C.
Preparation of experimental concentrations
The choice of tested concentrations (equivalent to 5% and 10% of 48-h LC50) was based on LC50 values obtained in acute bioassays with Musca domestica. Stock solution was prepared and used for making further dilutions. The extracts were selected for further study following trial runs with various concentrations of the extract.
Method of treatment
For each set of the treatment, twenty late 3rd instar larvae were selected. Seven numbers of glass beakers of 250 ml capacity were taken and labeled for different concentrations in addition to one for check and one for control. In case of control, water and for check, ethanol was added in place of extract. Larvae were dipped into the solution for two minutes and then transferred back in the rearing medium. Each experiment was conducted in triplicates along with the control group. Dose dependent mortality of larvae was observed. Mortality of larvae followed by the exposure was recorded after 24h up to 48h. and LC50 was calculated [24]. In brief, the mortality due to treatment of 3rd instar larvae with different concentrations of the leaf extracts of the two plants was recorded after 24 and 48h. The LC50 value was determined as described in Tables 2 and Table 4.
After determining the LC50 value, third instar larvae (20 in number) were treated with 5 and 10% of LC50 of the extracts for 48h. The experiment was repeated thrice.
Phytochemical analysis of the extract
Qualitative phytochemical analysis of seed extracts of C. procera and A. squamosa was done by the method as followed by Begum et al. [23]. In brief, the phytochemicals such as tannins, alkaloids, saponins, flavonoids, terpenoids and phenols/polyphenols were qualitatively determined.
Assay of AChE activity
The activity of AChE was determined according to the method of Ellman et al. [25]. The tissue homogenate was centrifuged at 1,000g for 15min at 4°C in a refrigerated centrifuge (Model No TLX 361 544). The resultant supernatant was recentrifuged at 20,000g for 20min at 4°C and used as an enzyme source. The reaction mixture (3ml) contained 1.5 ml of 100mM sodium phosphate buffer (pH 8.0), 0.3 ml of 5 mM DTNB [5,5'-dithiobis-(nitrobenzoic acid) prepared in 10 mM sodium phosphate buffer, pH 7.5 containing 15 mg sodium bicarbonate added per 10 ml of solution], 0.3 ml of 5 mM acetylthiocholine iodide (ATI), 0.1 ml of 10% homogenate, and 0.8 ml of distilled water. The increase in absorbance was monitored at 412 nm and 28°C for 3 min in a UV-visible double beam spectrophotometer (ELICO, model: SL-160) with quartz cuvettes (1 cm light path) against distilled water as blank. Measurements were made in triplicate for each tissue homogenate. Simultaneously, two blanks were also used. One blank contained phosphate buffer, DTNB, and ATI but not enzyme protein to determine the spontaneous hydrolysis of ATI, and the second blank contained phosphate buffer, DTNB, and enzyme protein but no substrate (ATI) to correct for any non-AChE-dependent formation of thionitrobenzoic acid (TNB). One unit of AChE activity was expressed as nanomoles of substrate hydrolyzed/min/mg protein under experimental conditions.
Statistical analysis
All values are given as mean ± standard error of mean (SEM) of three independent experiments. All data were examined using Students 't' test. Graph pad software was used to obtain level of significance.
The results presented in Tables 1 and 3 exhibits the toxicity of Calotropis and Annona seed extracts against M. domestica larvae, respectively. The treatment of 3rd instar larvae of M. domestica with different concentrations of the seed extracts of these two plants exhibited relatively lower percent mortality after shorter duration (24h) than that at longer duration (48h). The exposure of the flies to both of the ethanol extracts caused significant mortality in a dose dependent manner. The ethanol extract of C. procera was found to be quite effective against M. domestica larvae as nearly 100% mortality was observed at 500ppm. The mortality curves for the determination of LC50 values for the ethanol extracts of C. procera and A. squamosa are shown in Figures 1 and 2, respectively. The LC50 values of C. procera and A. squamosa extracts were calculated to be 870 and 345ppm, respectively (Tables 2 and 4).

Figure 1: Mortality curve of Musca domestica larvae for the determination of LC50 of C. procera seed extract. The graph was plotted using the data presented in table 1.

Figure 2: Mortality curve of Musca domestica larvae for the determination of LC50 of Annona squamosa seed extract. The graph was plotted using the data presented in table 3.
|
Concentration (ppm) |
No. of live Larvae |
% Alive at |
% Mortality at |
|
|
24 h |
48 h |
|||
|
0 (control) 0 (check) 200 400 600 800 1000 1200 1400 |
20 20 20 20 18 17 12 10 6 |
20 20 20 18 14 12 8 5 0 |
100 100 100 100 100 100 100 90 90 70 85 60 60 40 50 25 30 0 |
0 0 0 0 5 0 100 10 10 30 15 40 40 60 50 75 70 100 |
The larvae of M. domestica (20 in each set) were treated with different concentrations of ethanol extract of C. procera seed for 24 and 48 h as shown in Materials and Methods. The control and check represent larval treatment with water and ethanol, respectively. The experiments were conducted in triplicate.
Table 1: Toxicity testing of ethanol extract of Calotrpis procera seed against Musca domestica.
|
Concentration (ppm) |
Concentration difference |
No. of alive larvae |
No. of dead larvae |
Mean death |
Mean death Concentration difference |
|
0 (control) 0 (check) 200 400 600 800 1000 1200 1400 |
0 0 200 200 200 200 200 200 200 |
20 20 20 18 14 12 8 5 0 |
0 0 0 2 6 8 12 15 20 |
0 0 0 1 4 7 10 13.5 17.5 |
0 0 0 200 800 1400 2000 2700 3500 |
|
Total 10,600 |
|||||
The LC50 value of the seed extract of Calotrpis procera for 48 h has been determined according to the arithmetic method of Karber (1931). The calculation was done as following:
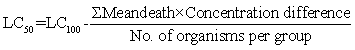
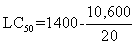
LC50 = 1400 – 530
LC50 = 870ppm
Table 2: The LC50 value of ethanol extract of Calotrpis procera seed for 48h.
|
Concentration (ppm) |
No. of live Larvae |
% Alive at |
%Mortality at |
|
|
24 h |
48 h |
|||
|
0 (control) 0 (check) 100 200 300 400 500 |
20 20 20 18 16 15 08 |
20 20 19 15 12 9 0 |
100 100 100 100 100 95 90 75 80 60 75 45 40 0 |
0 0 0 0 0 5 10 25 20 40 25 55 60 100 |
The larvae of M. domestica (20 in each set) were treated with different concentrations of ethanol extract of seeds of A. squamosa for 24 and 48 h as shown in Materials and Methods. The control and check represent larval treatment with water and ethanol, respectively. The experiments were conducted in triplicate.
Table 3: Toxicity testing of the ethanol extract of Annona squamosa seed against Musca domestica.
|
Concentration (ppm) |
Concentration difference |
No. of alive larvae |
No. of dead larvae |
Mean death |
Mean death Concentration difference |
|
00 (control) 00 (check) 100 200 300 400 500 |
0 0 100 100 100 100 100 |
20 20 19 15 12 9 0 |
0 0 1 5 8 11 20 |
0 0 0.5 3 6.5 9.5 11,5 |
0 0 50 300 650 950 1150 |
|
Total 3100 |
|||||
The LC50 value of the seed extract of Annona squamosa for 48 h has been determined according to the arithmetic method of Karber (1931). The calculation was done as following:
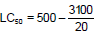
LC50 = 500 – 155
LC50 = 345ppm
Table 4: LC50 value of the seed extract of Annona squamosa for 48 h.
The phytochemical analysis of the extract (Table 5) showed the presence of alkaloids in maximum amount in ethanol extract of C. procera seed, which was tracely found in A. squamosa. The flavonoid contents were found in good quantity in A. squamosa extract. In addition, the terpenoids and alkaloids were also found to be present in ethanol extract of A. squamosa in traces. These two compounds were found to be missing from C. procera extract as determined by the method used in this study. Tannins and Saponins could not be detected in both of these extracts.
| Chemicals | Calotropis procera | Annona squamosa |
| Tannins | _ | _ |
| Flavonoids | _ | ++ |
| Tarpenoids | _ | + |
| Phenols | _ | _ |
| Alkaloids | + + | + |
| Saponins | _ | _ |
The phytochemicals in the ethanol extracts of the Calotropis procera and Annona squamosa leaves were determined as described in Materials and Methods.
- absent; + moderately present; ++ highly present
Table 5: Phytochemical analysis of ethanol extracts of Calotropis procera and Annona squamosa seed.
The present study also revealed the effect of in vivo exposure of the third instar larvae for 48 h to 5% and 10% of LC50 concentrations of the ethanol extracts from seeds of the plants, C. procera and A. squamosa on the the level of AChE activity in three developmental stages (larva, pupa and adult) of M. domestica.
The results presented in the Tables 6 reflected that the exposure to both the extracts caused significant reduction in the activity of AChE in different developmental stages of M. domestica as compared to control. Both, the low (5%) and high (10%) concentrations of these extracts were able to significantly inhibit the AChE activity from the insect larvae (Table 6). In the larval stage, the extract from A. squamosa seed caused maximum inhibition (48.01%) while seeds extract C. procera of was relatively less (37%) effective. Similar trend of inhibition was displayed by the extracts from seeds of both the plants for other two stages (pupa and adult) of the insect. The A. squamosa extract appear to be more effective for the larvae and adult stages of the insect than the pupa stage at both of the concentrations tested. The trend of sensitivity to the high concentration of the C. procera extracts on the insects developmental stages was as following: adults>larva>pupa. The effect was, however, in concentration dependent manner.
| Developmental stage | Control (%) | Seed extracts | |||
| Annona squamosa 5% 10% | Calotropis procera 5% 10% | ||||
| Larva | 46.47 ±0.05 | 30.12**±0.16 (-35.17) | 24.15*±0.65 (-48.01) | 37.69*±0.47 (-18.89) | 29.23* ±0.32 (-37.08) |
| Pupa | 67.56 ±0.29 | 58.87*±3.46 (-12.85) | 41.79*±4.21 (-38.14) | 61.23ns±1.47 (-9.36) | 49.34 ns±5.05 (-26.97) |
| Adult | 195.98 ±0.93 | 157.65*±0.84 (-19.56) | 85.57**±1.26 (-56.34) | 162.56*±0.63 (-17.05) | 112.76**±0.88 (-42.47) |
The AChE activity in three developmental stages of the insect was assayed as indicated in Materials and Methods. Data were analyzed by Graphpad software, values given as mean ± SEM Values in parentheses are percent change over control. Significance (*) of data is shown in superscripts, nsP>0.05(non significant), *P<0.05(significant), **P<0.01(very significant), C-Control (0%), 5% & 10% are 1/20 and 1/10 of LC50.
Table 6: Effect of Annona squamosa and Calotropis procera ethanol extracts on AChE activity (nanomoles of ATI hydrolyzed/min/mg protein) of Musca domestica.
When these developmental stages of the insect were exposed to the low (5%) and high (10%) concentrations of the extracts from seeds of the plant, A. squamosa, the seeds extract inhibited the enzyme activity to greater extent as compared to C. procera. However, in the adult stage the inhibition caused due the treatment with both extracts at low concentration (5%) was similar (about 19 and 17 %), whereas there was marked difference in their effects at high concentration (10%); the C. procera and A. Squamosa seed extracts exhibiting 42 and 56.34% inhibition, respectively. All the three stages of the insect exhibited higher sensitivity to both concentrations (5 and 10%) of the extract from seeds of A. squamosa than C. procera. The pattern of sensitivity to the high concentration of the A. squamosa extracts on the insect's developmental stages was as following: adults>larva>pupa. The effect of the plant extracts was concentration dependent (Table 6).
The results presented in Tables 6 demonstrate that the extract from the seeds of A. squamosa was comparatively more inhibitory to the enzyme activity in all the stages of the insect than that of C. procera. It is note worthy that the extracts from both of the plants displayed less inhibitory influence on the pupa stage of M. domestica. The negative effect of the extracts from both of the plants was concentration dependent on all the three developmental stages of the insect.
The accumulation and biomagnifications of synthetic compounds into different non-target organisms, including humans through food chain with increased risk of the development of diseases or disease syndromes has prompted to explore for relatively safer and more potential molecules for better insects-pests management.
The results obtained from the present study with the extracts of seeds of two different plant species indicated that the A. squamosa seed extract exhibited higher insecticidal potential. The LC50 values of both of these extracts recorded in this study may be due to the crude nature of the ethanol extracts used in the present study. The crude plant extracts contain many active as well as inactive compounds which may act synergistically to enhance a specific bioactivity or antagonistically to mask certain activities.
Previous investigations on annonaceous acetogenin, the bioactive principle of the plant family Annonaceae, have shown to possess pesticidal or antifeedant properties [26,27]. Seed oil of A. squamosa has been reported to reduce survival of leaf hopper, Nephotettix virescens (Hemiptera: Cicadellidae) and transmission of rice tungro virus [28,29].
Similarly, although reports on nematicidal [30], antimicrobial and antihelminthic [31] activities of C. procera extract and its use in the treatment of toothache, cough and subcutaneous diseases [32] exist, there is no report at all regarding the LC50 for the alcoholic extract of C. procera seed against M. domestica. The laboratory study on larvicidal properties of leaf extract of C. procera against housefly [23] and mosquito larvae is known [33].
Death of the larvae exposed to the ethanol extracts of these two plants in present study may be due to the inability of the moulting bodies to take up sufficient volume of air to split the old cuticle and expand the new one during ecdysis or metamorphosis. The inhibitory effects of plant extracts are possibly due to the perturbations in the hormonal regulation in the insect [34].
Antifeedant, growth inhibitory and toxic effects of crude methanolic seed extracts of Annona squamosa and Annona atemoya (LC50 = 167.5 ppm and 382.4 ppm respectively) against the cabbage looper, Trichoplusia ni (Hübner) (Lepidoptera: Noctuidae) has been earlier evaluated in a greenhouse experiment using different bioassays [35].
AChE is a key enzyme in the nerve impulse transmission system of any living organisms. It catalyzes the hydrolysis of the neurotransmitter, acetylcholine, at the synaptic cleft (the space between the two axonic ends of nerve cells) so that the next nerve impulse can be transmitted across the synaptic gap. Inhibition of AChE causes accumulation of AChE at the synapses, so that the post-synaptic membrane is in a state of permanent stimulation, which results in paralysis, ataxia, general lack of co-ordination in the neuromuscular system and eventual death [36]. Several pesticides or insecticides of the organophosphate and carbamate groups are known to act to paralyze and kill insects by inhibiting the activity of this enzyme [37].
The results clearly indicated that among the seed extracts of two different plant species tested, A. squamosa seed extract exhibited the greater potential to inhibit the AChE activity, as only 44 % activity remained in adult housefly after 48h of treatment at higher concentration (10% of LC50). However, in larva and pupa stages, the enzyme activity was reduced by about 48 and 38%, respectively, at this concentration. However, the seed extract from another plant, C. procera, registered lesser AChE inhibition potential, inhibiting nearly 42% activity in adult. Whereas in larvae and pupae it was reduced by 37 and 27% respectively. The varying pattern of reduction in the enzyme activity by the application of extracts from seeds of these two plants indicates the different chemical nature of the extracts acting as AChE inhibitors. The death of the insects due to treatment with higher concentrations of these extracts suggest that the molecules present in these preparations possibly interfere at the cholinergic synapse and destroy the communication network from one exonic end to the another thereby blocking the nerve impulse transmission.
In a similar study, a significant decrease in AChE activity in brown planthopper Nilaparvata lugens due to the treatment with azadirachtin (AZA), a tetranortriterpene from neem has been shown [38]. Some other workers have reported that the exposure of nymph of Blattella germanica L to reserpine and neem extract caused 52.68 and 38.72 % reduction in the activity of AChE, respectively [39]. The significant inhibitory effect of plant extracts on AChE activity at low concentrations in the present study indicates that even a smaller quantity of the extract is able to enter the insect body tissues, cross the blood brain barrier and inhibit the AChE activity. The lethal effect on the larvae exposed to different extracts tested may be due to the accumulation of acetylcholine (ACh), a neurotransmitter, at synaptic junctions, which interrupts the coordination between the nervous and muscular junctions (neurotoxicity).
AChE has been reported to be the site of action of plant based anticholinesterase insecticides and their active intermediates. The inhibition of the AChE activity at higher doses of Azadirachtin treatment was observed in the cockroach, Periplaneta americana L [40] and the snail, Limnaea acuminata Lamarck, at 40% and 80% concentrations of neem oil [36]. It was also observed that 25g distilled water extracts of the botanicals Punica granatum L., Thymus vulgaris L., and Artemisia absinthium L., significantly inhibited the AChE activity of nematodes at very high (100%) concentrations [41].
In addition, several essential oils from aromatic plants, monoterpenes, and natural products have been shown to act as inhibitors of AChE [42]. Pulegone-1,2-epoxide, isolated from the Verbenaceae medicinal plant, Lippia stoechadifolia L. (Poleo), showed an irreversible inhibition of the AChE in house fly and Madagascar roach [43]. Similarly six monoterpenes, Pulegone (0.12×104 ppm), gossypol (0.45 × 104 ppm), citral (1.5 × 104 ppm), linalool (2.5×104 ppm), bornyl cetate (2.7×104 ppm), and cineole (4.3×104 ppm), representing characteristic constituents of secondary plant metabolites were tested on AChE from the eel in vitro and on toxicity in vivo in a stored product pest, Tribolium castaneum Herbst. All six terpenoids inhibited activity of AChE [44]. Also, it has been demonstrated that the essential oils from the plants belonging to the Labiatae family caused inhibition of AChE activity in similar manner. This enzyme was isolated from the stored product insect, Rhizopertha dominica Fab [45].
As compared to other developmental stages of house fly, minimum percentage change in inhibition of AChE activity has been found in pupae with all the treatments which probably could be because of this stage being less sensitive to these plant extracts. This is in accordance with earlier reports showing that pupae exhibit a higher tolerance to chemical agents than active stages [46]. Acanthoscelides obtectus (Say) larvae were found to be more susceptible to the fumigant toxicity of the essential oils from Lavandula hybrida (Rev), Rosmarinus officinalis L. and Eucalyptus globules (Lab) than pupae [47]. In addition, pupa of Leptinotarsa decemlineata (Say) has also been found less sensitive to the Piper nigrum L. extracts [48].
In conclusion, the extracts from the seeds of C. procera and A. squamosa at high concentration (10%) exhibited maximum toxicity and AChE inhibition potential in all the three developmental stages of the insect tested. Their effects were concentration dependent. The extracts from both plants were less effective to the pupa stage of M. domestica. The results of this study raises the possibility that the insecticidal and antiAChE properties of the active compound(s) present in the tested plant extracts could be exploited as an alternate of many synthetic chemical insecticides being indiscriminately used for control of insects pests. The extracts of these plants may be useful in the management of M. domestica and the transmission of different pathogens associated to it.
The authors express their gratitude to Prof. P. Gaur, Ex-Head, Department of Zoology, University of Allahabad for providing necessary facilities. Financial assistance to one of the authors (NB) from the University of Allahabad (UGC) and UGC-SAP (RFSMS) to the Department of Zoology is gratefully acknowledged.