Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Rapid Communication - (2016) Volume 5, Issue 1
Due to limited availability of literature on the spawning activity of Malaysian horseshoe crab, Tachypleus gigas (Müller, 1785), the reproduction behaviour and biology of this arthropod remains poorly understood. Hence, an investigation was carried out from April-July to trail spawning horseshoe crab amplexus at Balok and Cherating, the only known T. gigas spawning grounds on the east coast of Peninsular Malaysia. Through visual tracking during daytime full moon spring tides, the release of air bubbles indicate nest digging by female crabs. While air bubble formation aggravated, flagged aluminium poles were carefully driven into the sediment to mark the nest. Out of the 13 spawning T. gigas amplexus tracked, only one pair was able to dig up to 12 nests and release up to 2,575 eggs within the 2.5 hour spawning period. The female crabs dug nests at the intertidal regions, ranging 1.3-43.2 m from the highest tide marking on the beach. Interestingly after spawning, the horseshoe crabs returned to high seas during the residing tides as amplexus because the male crabs remained clinging onto the female crab’s opisthosoma. The incidence of recapturing a tagged female crab with different attached male crab at Cherating indicates this spawning ground as natal beach. In addition, the presence of 8 horseshoe crab amplexus at Balok compared to only 5 amplexus at Cherating highlight preferable beach settings for spawning and/or the presence of larger spawning crab assemblages at the former beach. Since only 13 spawning horseshoe crab pairs were witnessed at Balok and Cherating, identification of factors that alter beach sediment composition and water chemistry as well as gender-specific poaching and excessive boating activities should be regulated to restore wild T. gigas populations.
<Keywords: Horseshoe crab; Mating Xiphosura; Site-specific spawning; Trailing and tagging; Nesting ground
Horseshoe crabs exist since the Ordovician period and have remarkable evolution history which stopped 250 million years ago [1]. These crabs are survived by Asian horseshoe crabs, Tachypleus tridentatus (Leach), Tachypleus gigas (Müller), Carcinoscorpius rotundicauda (Latreille) and the American relative, Limulus polyphemus (Linnaeus) [2,3]. In Malaysia, all three Asian horseshoe crabs are available, in which, T. gigas and C. rotundicauda are present in the coastal areas of Peninsular Malaysia whereas, T. tridentatus is restricted to East Malaysia at Sabah and Sarawak [4]. Although the co-existence of C. rotundicauda and T. gigas at Balok raised curiosity of researchers yet, the spawning preference of T. gigas at sandy beaches unlike C. rotundicauda at muddy substrate was more practical and this, made the former crab a suitable candidate for research [5-9]. On the other hand, the appearance of spawning T. gigas at Cherating is a new discovery for spawning areas on the east coast of Peninsular Malaysia.
Horseshoe crabs are encountered only when they come ashore to shallow and surf-protected beaches for nesting. A male crab attach to the rear end of one female crab using their pedipalps is distinguished as a mating pair or amplexus [10-12]. A female crab is usually capable of laying 200-300 eggs [13] in varying depths (5-20 cm) below the sand [14,15]. However, the crab’s spawning output is largely influenced by season and local environmental, such as edaphic and hydrology conditions [16,17].
Following the usual method to obtain horseshoe crabs in Malaysia via net catching [18,19], the assessment of spawning crabs by hand collection is only limited to the studies of [15,20,21]. Although covering different sites, estimation of spawning crabs were based on arrivals and random finding of nests during low-tide periods. Thus, the present approach is first of its kind, because horseshoe crab amplexus were tracked and trailed and, every nest dug by the female crab was marked. In addition, this is the first study which assesses the maximum nestingcapability by a spawning T. gigas pair, in relation to the abundances and incidences of egg release by the female horseshoe crab.
Study site description
Balok (N 03°56.009’ E 103°22.584’) and Cherating (N 04°07.261’ E 103°23.353’) is known for horseshoe crab spawning on the east coast of Peninsular Malaysia (Figure 1). Since both locations are only 28 km apart, these sites have similar environmental patterns such as hot and humid climate with temperatures varying at 24-36°C, annual average rainfall of 1567.5 mm and mixed tides that range from 0.1-3.5 m [22,23]. In addition, both, Balok and Cherating are estuaries that receive human influences via boating, fishing and recreation activities.
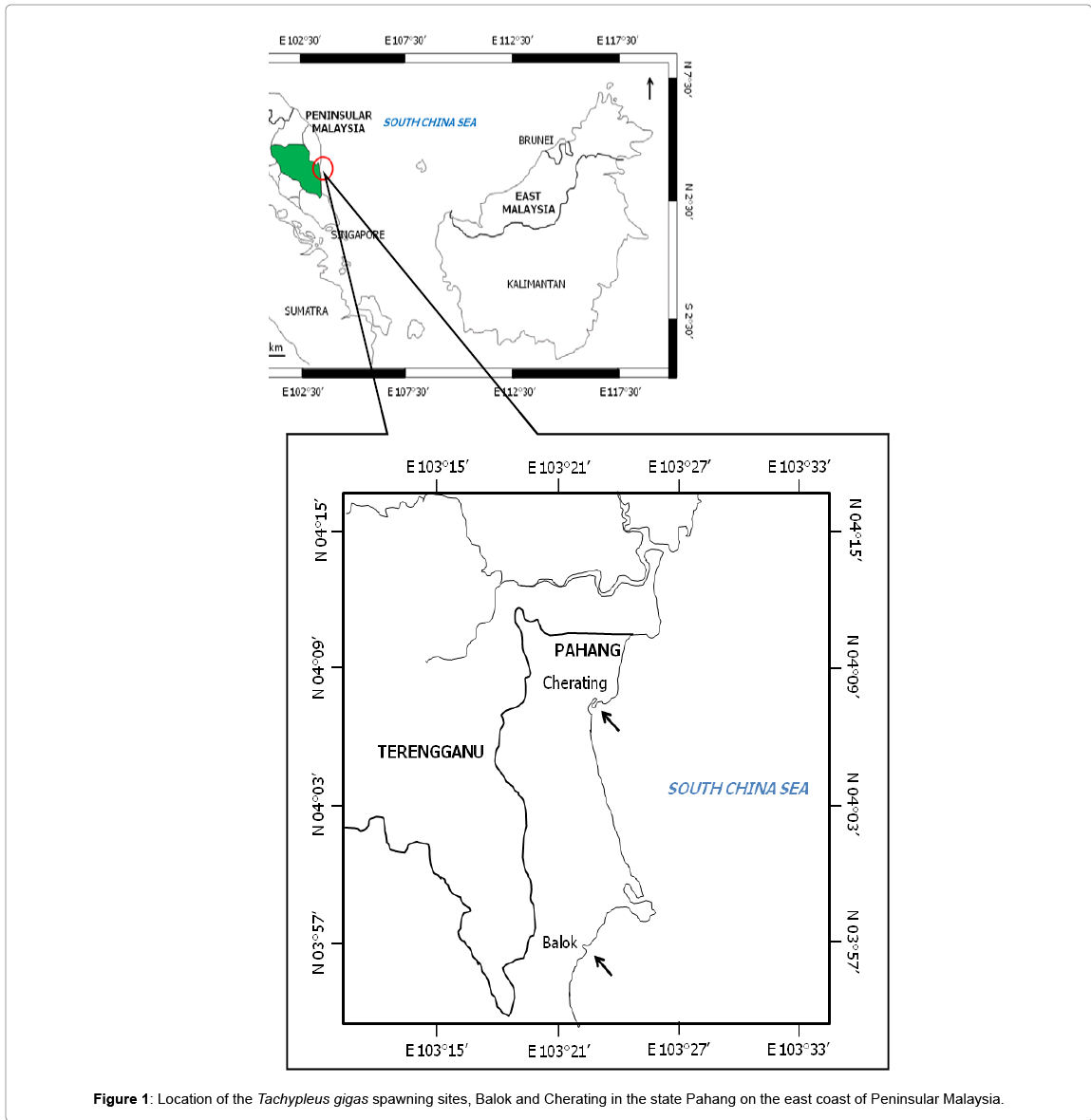
Figure 1: Location of the Tachypleus gigas spawning sites, Balok and Cherating in the state Pahang on the east coast of Peninsular Malaysia.
Field visits at Balok and Cherating
Field visits to Balok and Cherating were carried out in four consecutive days during the 15th lunar day or full moon periods from April-July 2013. Tracking of spawning horseshoe crabs were carried out one hour before day-time full moon spring tides and the survey lasted up to 2.5 hours daily. The crabs moved along the spawning beach and release bubbles while searching for suitable substrate. During vigorous release of bubbles by the horseshoe crab amplexus for more than two minutes, an indication of sand digging to lay eggs, flagged aluminum poles were carefully driven on each side of the female crab’s prosoma-opisthosomal hinge to mark egg deposition locations (nests). Coordinates of potential nests made by every spawning amplexus were retrieved via Global positioning System (Garmin 5.0). This was followed by acquisition of physicochemical parameters of the water such as water temperature, salinity, dissolved oxygen (DO), pH and total dissolved solid (TDS) at 10 cm depth using a multi-sensor probe (YSI 556 MPS, Ohio, USA).
The spawning activity of T. gigas is ended when the crab moves towards deeper water. These horseshoe crab amplexus were gently handpicked and separated to acquire their prosomal width using a measuring tape. Then, the carapace-tip of each crab was punctured (using scalpel) to fit the unique identification (ID) button tags. Identification of crabs via button tags eases data collection, prevents recounting and identifies returning crabs (if any). After tagging, the horseshoe crabs were released into the water. Low tide re-visitations to Balok and Cherating were carried out to identify potential nests, retrieve the flagged aluminium poles and to unearth the newly fertilized horseshoe crab eggs using a plastic hand shovel. It followed with counting of the eggs after sand removal using 2 mm mesh size plastic sieve [24]. All horseshoe crab eggs were returned to their nest and covered with the sand. The distance between nests and location of nests from the highest tide marking on the beach were also estimated using measuring tape.
Data analysis
Only spawning horseshoe crabs that released eggs were considered as research model in this study. The crab’s eggs and nests were identified as spawning output. Statistical Package for the Social Sciences (SPSS v.17) was used for T-test (defined ‘T’) and One-way ANOVA (defined ‘F’) analyses to estimate significance ‘P’ values and, to correlate via Spearman’s test (defined ‘X’) the results obtained. Step-wise analysis, defined ‘BEST’, was carried out using Primer v.6 to determine important relationships between the horseshoe crab and their pattern of spawning at Balok and Cherating.
Spawning yield at Balok and Cherating
Throughout the spanning field work, three T. gigas in amplexus were observed at Cherating whereas another six were observed at Balok. Interestingly, no crabs were present at Balok during May 2013 although comparatively one T. gigas amplexus arrived to spawn at Cherating beach during that time. Prosomal width disparities exist among these crabs where it ranged 22.9-25.1 cm for Balok crabs and it ranged 18.3- 23.2 cm for Cherating crabs. Despite the poor abundance of spawning crabs at Cherating, the highest abundance for eggs (2575 eggs/attempt) and nests (up to 12/attempt) were recorded from this beach (Table 1). Also, female crabs at Cherating were more productive, with 16 nests containing 4118 eggs, than at Balok where, only 9 nests containing 2624 eggs were discovered.
| Site | Month | Sw pair (no.) | FPW (cm) | Nt (nos.) | SwT (min.) | DHTL (m) | NtD (cm) | Eggs (nos.) | SDt (m) | Temp (ºC) | Salinity (‰) | pH | DO (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cherating | May | 1 | 18.3 | 2 | 7.5 ± 3.5T (5-10) ∑=15 |
5.3 ± 1.2D (4.5-6.2) |
19.5 ± 0.7N (19.0-20.0) |
263 ± 240E (93-433) ∑=526 |
76.9* | 32.2 | 30.3 | 7.8 | 78.5 |
| June | 1 | 23.1 | 4 | 9.5 ± 1.9T (8-12) ∑=38 |
5.4 ± 0.3D (5.0-5.7) |
20.3 ± 3.9N (16.5-25.5) |
254 ± 47E (212-306) ∑=1017 |
15.9 ± 14.6S (0.3-29.4) ∑=63.8 |
31.9 | 31.6 | 7.8 | 79.7 | |
| 2 | 23.1 | 12 | 9.8 ± 1.7T (7-12) ∑=118 |
6.4 ± 0.3D (5.9-6.9) |
18.5 ± 2.6N (15.0-23.0) |
215 ± 43E (135-277) ∑=2575 |
0.10 ± 0.05S (0.03-0.20) ∑=1.2 |
32.9 | 31.4 | 8.2 | 79.5 | ||
| Balok | June | 1 | 22.3 | 1 | 6* | 43.2* | 14.5* | 309* | 0 | 32.8 | 31.3 | 7.9 | 80.9 |
| 2 | 24.0 | 2 | 4.5 ± 0.7T (4-5) ∑=9 |
27.4 ± 4.7D (24.1-30.7) |
26.5 ± 1.4N ( 23.0-30.0) |
426 ± 28E (406-446) ∑=852 |
3.4* | 33.4 | 31.0 | 7.8 | 82.4 | ||
| 3 | 25.1 | 1 | 2* | 20.4* | 20.5* | 251* | 0 | 33.4 | 31.0 | 7.8 | 82.4 | ||
| 4 | 24.5 | 3 | 5.3 ± 0.6T (5-6) ∑=16 |
15.5 ± 16D (15.4-15.6) |
19 ± 1.7N (17.0-20.0) |
364 ± 8E (360-373) ∑=1093 |
7.2 ± 4.5S (4.0-10.4) ∑=21.7 |
33.2 | 31.0 | 8.0 | 82.9 | ||
| 5 | 22.9 | 1 | 5* | 19.6* | 17.5* | 199* | 0 | 33.4 | 31.1 | 7.8 | 82.4 | ||
| 6 | 23.1 | 1 | 10* | 15.9* | 15.5* | 171* | 0 | 33.2 | 31.0 | 8.0 | 82.9 |
Sw = spawning, FPW = female prosomal width, Nt = nest, SwT = spawning duration/time, DHTL = distance from highest tide level, NtD = nest depth, SDt = spawning distance (first to last nest distance), Temp. = water temperature and, DO = dissolved oxygen. The prefix, * = single value, T = time per nest, D = average distance of nests from highest tide, N = average nest depth, E = number of eggs per nest and, S = average spawning distance. Units, (no.) = count, (m) = meter(s), (cm) = centimetre(s), (min.) = minute(s) and, (nos.) = amount/quantity.
Table 1: Tachypleus gigas spawning characteristics and yield in May-June 2013
While horseshoe crab nest and egg abundance was related to the female crab’s prosomal width (X = 0.374; T =2.846, P =0.036), the spawning output per female crab was lower at Cherating (257 eggs/ nests) than at Balok (291 eggs/nest) (F = 10.2, P = 0.02). Although the female horseshoe crabs nested at almost similar depths, 18.9-19.2 cm, the female horseshoe crabs from Cherating required 8.9 minutes to nest. Comparatively, female crabs at Balok only needed 5.6 minutes to nest. With regard, there is strong correlation between eggs released by the female crab and the time spent to nest during a spawning event (Best = 0.702; X = 0.206; T = 13.3, P = 0.001).
Tachypleus gigas spawning pattern
During the Southwest (SW) monsoon (May-June) survey, salinity and water temperature at Balok and Cherating were at 31‰ and 31- 33ºC (Table 1). While pH remained the same at 7-8, the dissolved oxygen was increased at Balok (80-82%) compared to Cherating (79- 80%). These water properties made the SW monsoon important for the horseshoe crab spawning at Balok and Cherating (BEST = 0.466; X = 0.098; T = 25.5, P = 0.02). In addition, there were demarcations for the horseshoe crab spawning preference on the beach where, at Balok, female crabs made their nests 15-27 m away from the highest tide mark whereas at Cherating, it was only 5-6 m away (Table 1).
An interesting discovery was made at Cherating where, a spawning horseshoe crab amplexus laid 2575 eggs in 12 nests. The maximum distance per nest of this spawning horseshoe crab amplexus was only 0.2 m apart and it covered a total distance of 1.2 m on the beach. Since this is the highest number of nests produced by a horseshoe crab in amplexus, its nesting behaviour was used to describe the spawning pattern of T. gigas (Figure 2). The nests 1-3 (A-C) were made by the spawning horseshoe crab amplexus during the highest tide period whereas the remaining nests (nest 4-12, D-L) were constructed by the amplexus during the residing tide. From nest 4 (D) onwards, the crab begun nesting at the water edge and its nest placement on the beach increased in distance from the highest-tide mark (Figure 2). In addition, this spawning horseshoe crab amplexus took from 7-12 minutes to construct a nest and their spawning activity lasted 118 minutes before they decided to move into deeper water (Table 1).
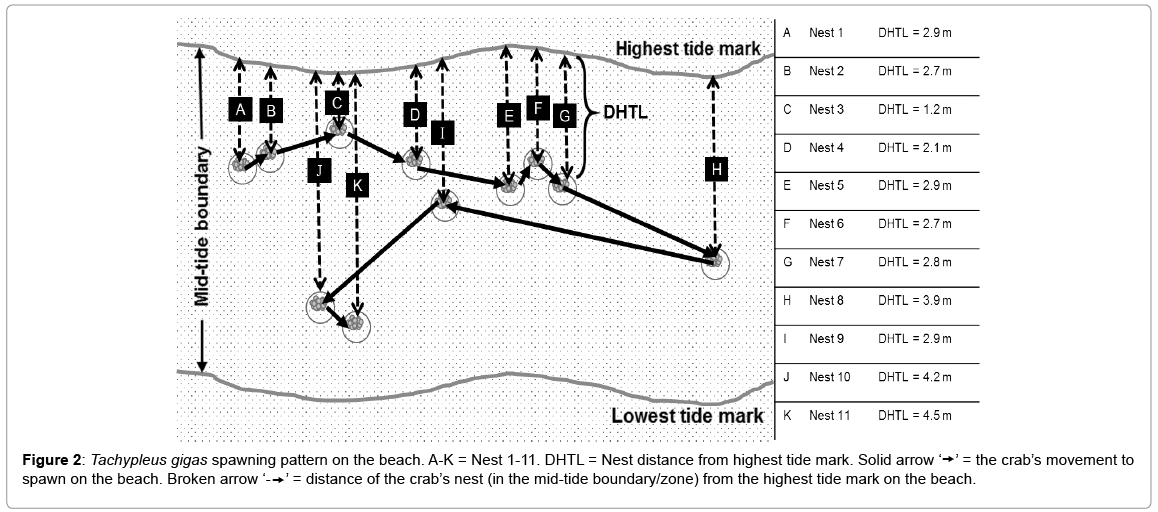
Figure 2: Tachypleus gigas spawning pattern on the beach. A-K = Nest 1-11. DHTL = Nest distance from highest tide mark. Solid arrow ‘’ = the crab’s movement to spawn on the beach. Broken arrow ‘- ’ = distance of the crab’s nest (in the mid-tide boundary/zone) from the highest tide mark on the beach.
Low population of crabs
Unlike observations that focused on horseshoe crab assemblages [16,25], the present investigation observed the spawning activity of every T. gigas mating pair. While acoustic transmitters were sensitive to wind and time consuming to locate crabs [26,27], manual tracking of spawning horseshoe crabs by observing the release of bubbles before and after spring tide is effective, feasible and low cost. Presently, the emergence of T. gigas to Balok was lower than previous records [15,18, 20]. Past surveys at Tanjung Selangor, the only T. gigas spawning site with long term investigations in Malaysia showed that beach conditions during SW monsoon was most favourable to the horseshoe crab for spawning [21]. Although field visits covered the most productive spawning season, as claimed by [5,6], spawning crabs were only spotted at Balok beach during June. Moreover, there is disagreement with [28] because T. gigas does not spawn year-round, unless this animal also spawns during the period which was not covered in this study.
Highlighting the interesting discovery of only male T. gigas entangled in crab nets during 2009 and 2013 [29], gender-specific poaching was thought to occur at Balok. In fact, there was demand for female horseshoe crabs at Balok. Also, size demarcation made T. gigas more preferable than C. rotundicauda [30]. Perhaps, the horseshoe crab’s low market value (USD 0.55/RM 2.22 per crab) made them a cheaper ‘protein source’ than fish, decapod crabs, mussels and prawns. Although Cherating supports the spawning activity of T. gigas, due to limited research and field visits, the fate of horseshoe crabs there remains unknown. However, since there is a fishing village at Cherating, the poor abundance of horseshoe crabs there was thought to be consequence of demands, the similar problem faced at Balok.
The arrival of three spawning amplexus at Cherating and another six at Balok is evident to small T. gigas populations in Malaysia [15,18,21] that consist of assemblages [31,32]. While dissimilar operational sex ratio (OSR) were observed in small population groups such as 1.07:1 at Maine [33] and up to 9:1 in larger groups [34], the absence of satellite male and solitary female produced OSR 1:1 at Balok and Cherating. Monogamous OSR is unique to small horseshoe crab groups because incidences of strategic mating such as satellite males or solitary partner-selective female are often seen [35,36]. Since poaching was thought to occur around Balok and Cherating, disturbances to the horseshoe crab’s population structure highlight large-scale harvesting to meet demands locally at Penang and Johore as well as abroad in Vietnam and Thailand [37].
Beach conditions that favour Tachypleus gigas spawning activity
From the first to the last nest placed on the beach by spawning horseshoe crab amplexus, the nest distance from the highest tide mark varied more than three-folds at Balok and Cherating. Although this horseshoe crab spawning trend coincides with residing tides, the crab’s nest placement on the beach was also thought to be influenced by steepness of the beach itself [38,39]. In this case, T. gigas spawning activity at Balok and Cherating correspond to beach slope that offered water cover (Figure 2). Balok beach inclined ±10% from mean sea level (MSL) [40]. As it was uncertain for Cherating, the distance (beach length) required to reach 1 m depth was estimated in-situ.
To achieve 1 m water depth, only 15 m beach length was required at Cherating, compared to Balok which required up to 40 m. Hence, the horseshoe crabs of Cherating were able to spawn 5-6 m from the highest tide mark (narrow mid-tide boundary) on the beach. In the case of Balok, the mid-tide boundary was broad and the crabs spawned more than 15 m from the highest tide mark. Despite Balok having the advantage of reduced beach steepness compared to Cherating, the crab’s spawning output (nest and eggs) there was poor. Perhaps, drawbacks were caused by newly trained field workers who failed to notice the spawning crabs earlier and this reduced the disclosure of nests and eggs at Balok. However, topography comparisons show Cherating to have sand bar shelter compared to Balok, a river mouth with direct opening to the sea. Horseshoe crabs are known to favour beaches with negligible tides/low wave current during spawning [41], a major setback at Balok during high tide.
In the present study, a female T. gigas could release up to 446 eggs. This opposed the previous suggestion by [42] that female T. gigas was only capable of releasing 300 eggs per clutch/nest. In fact, [15] also observed the female crab’s potential to lay up to 393 eggs. Major drawbacks in the population structure were caused by limited T. gigas amplexus abundance compared to L. polyphemus (>50 crabs arrive to spawn) [43-45] and, limited spawning sites along the east coast of Peninsular Malaysia [15,21,29,40,46]. Tachypleus gigas spawning output in terms of nest and eggs were very reduced, after comparing its spawning output with L. polyphemus because, the batch laying behaviour (several clutches) and unearthing-replacing of nest does not occur among horseshoe crabs from Malaysia [21,29], India [42,47-49] and Singapore [2].
At Cherating, T. gigas made their nests 0.03-29 m apart whereas at Balok, it was 3.4-10.1 m apart. At both sites, the female crabs only released eggs in one clutch per nest. While female horseshoe crab size and their spawning duration influenced the abundance and incidence of eggs released during a spawning act, environmental influences such as dissolved oxygen and pH influenced the horseshoe crab’s site selection on the beach. However, compared to the biology (size) of female horseshoe crab [50], environmental influences (duration of high tides) play a greater role for horseshoe crab spawning output such as number of eggs and nests [51].
Since, horseshoe crabs seek for well oxygenated sediments to nest at Balok [21,29,39,52] and Cherating, their spawning output pertains to how long the crab takes to find the ‘perfect nesting spot’. Hence, while some horseshoe crabs spend 40 minutes to spawn and make only 5 nests, an incidence of 2-3 hours spawning and making up to 12 nests by a T. gigas amplexus coincide with nesting site selection. Inevitably, horseshoe crabs risk various obstacles such as poaching and boating disturbances during their spawning. It follows with a long embryogenesis, up to 42 days, and long larvae-juvenile stages that lasts up to 11 years to before maturing into adult. Thus, these drawbacks at Balok and Cherating result to poor abundances of spawning horseshoe crab amplexus, which was seen as a major setback in the present study especially at active T. gigas spawning grounds.
Though the field visits spanned from April-July 2013, Tachypleus gigas only came to spawn at Balok and Cherating in May-June, during the Southwest monsoon period, because of favourable salinity and dissolved oxygen during that time. In addition, the horseshoe crab’s spawning activity at Balok and Cherating was influenced by beach elevation and locations on the beach that receive negligible tidal waves. While discrepancies of female prosomal width was thought to influence the abundance and incidence of egg release, the crab’s behaviour to spawn during high and residing tides made them have limited spawning time. In addition, T. gigas initiate their spawning near to the highest tide mark, at the mid-tide region of the beach while remaining fully submerged. Consecutive nests are made at the mid-tide region of the beach but it pertains to residing tides that offer full inundation. While the availability of spawning horseshoe crabs at Cherating is a new record for spawning grounds on the east coast of Peninsular Malaysia, major hindrance to fully understand Cherating and its capability as horseshoe crab spawning site was hampered by limited work time and budget. Hence, we insist on additional observations such as beach topography and sediment analysis as future perspectives to establish reasons for the shifting horseshoe crab spawning seasons at not only Cherating but, as well as at Balok. Perhaps, longer observation periods and additional statistics such as population size estimation could predict the actual population size instead of sole estimation on spawning crabs at both, Balok and Cherating.
We greatly appreciate Professor H. Jane Brockmann, from Department of Biology, University of Florida, Gainesville, Florida, USA for her valued comments and suggestions on our original manuscript. We are very grateful for the assistance of Izzatul Huda, Fairuz, Zahran, Azwa and Nazirul Amin, Biodiversity students from Universiti Malaysia Terengganu who helped with fieldwork and data collection.