Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 5, Issue 3
The aim of this study was to investigate the antiangiogenic effect of the Nigella sativa seeds extracts. Twelve extracts were prepared from the N. sativa seeds by supper critical carbon dioxide by varying extraction pressure and temperature. GC-MS analysis revealed the presence of 22 compounds, mainly thymoquinone (TQ), thymohydroquinone and androstane-3,17-diol. The extracts showed significant antioxidant activity, and cell viability study on human umbilical vein endothelial cells (HUVECs) indicated that two extracts (A3 and B3), prepared at higher temperature (60°C) and the lower pressures (2500 and 3000 psi), were the most potent anti-proliferative extracts; the median inhibitory concentrations were 41.5 ± 1.9 and 60.2 ± 2.2 μg/mL, respectively. The extracts down-regulated the expression of vascular endothelial growth factor (VEGF) in endothelia cells causing inhibition of various events of the angiogenesis cascade including; cell proliferation, endothelial cell migration and differentiation on matrigel and consequently inhibited tube formation. The extracts also showed significant inhibition of angiogenesis in vivo by inhibiting the vascularization of the chicken embryo chorioallantoic membrane (CAM). The antiangiogenic activity observed could be due to the anti-oxidant nature of the extract. This study highlights the importance of N. sativa SCCO 2 oil as anti-angiogenic agent.
Keywords: Nigella sativa; Supercritical CO2 extraction; Antiangiogenesis; Antioxidant
Angiogenesis is vitally required for the physiological functions such as reproduction, vascularization and wound healing. However, when angiogenesis gets dysregulated, it leads to numerous lethal diseases such as cancer, ischemia, and macular degeneration, and chronic inflammation, infectious and immune disorders. Activation of endothelial cells is a prerequisite angiogenic factor, which induces angiogenesis by stimulating the genetic expression of a chief proangiogenic mitogen, vascular endothelial growth factor (VEGF) [1]. Endothelial cell proliferation, migration, differentiation and survival activities are mediated by binding of VEGF to its receptor VEGFR-2 through tyrosine kinase signalling pathway [2,3]. As there is a strong interdependence between angiogenesis and malignant pathologies, angiogenesis could be an ideal therapeutic target to deal with the angiogenesis-related disorders. Thus, there is an increasing demand to discover and develop the anti-angiogenic agents to curb the pathological angiogenesis.
Essential oils from medicinal plants have been reported to possess strong antiangiogenic properties and thus essential oils have been used to treat various human ailments caused due to the excessive angiogenesis [4,5]. The widespread use of plant essential oils has recently gathered an intense attention and focus in the basic research on essential oils as phytomedicine [6-8].
Supercritical fluid extraction (SFE) is a useful technique for extraction of volatile oils from natural sources [9]. SFE is preferable method for extraction because it is solvent free and prevents secondary reactions such as oxidation. Supercritical carbon dioxide (SC-CO2) is the most common supercritical fluid used in SFE technology; it is fast, nonexplosive, non-toxic, inflammable, inert to solutes, environmentally safe and results in solvent-free extracts [10,11].
Nigella sativa Linn. (N. sativa) is a very common medicinal plant from the family Ranunculaceae, it is known as black seed [12]. It has been used traditionally as carminatives, condiments and appetizer. Black seeds have also been used for treatment of various diseases such as fever, jaundice, inflammation and [12,13]. Recent studies have shown that N. sativa possesses medicinally active essential oils with various pharmacological activities including anti-cancer and anti-metastasis [14], anti-cestodal [15], antibacterial [16], antiviral [17], antiinflammatory [18], antinociceptive [19], and antioxidant [20] activities. N. sativa fixed oils have showed cytotoxic and immunopotentiating activities in vitro [21,22]. Supercritical CO2 extracts of N. sativa have showed potent antibacterial activities [23], also it has resulted in selective anti-proliferative and induced apoptosis of breast cancer (MCF 7) cells [14].
The major chemical constituents reported in N. sativa volatile oil are thymoquinone, beta-elemene, nigellone, dithymoquinone and thymohydoquinone [24]. N. sativa fixed oil has been distinguished into saturated and unsaturated fatty acids. Saturated fatty acids include myristic, myristoleic, palmitic, palmitoleic, stearic, arachidic, arachidonic, behenic, lignoceric, and erucic acids. The unsaturated fatty acids contain oleic and linoleic acids [25-27]. Many other compounds have been identified in the oil, such as para-cymene [28], anthenole, carvacrol and 4-terpineol [29], carvone, cycloartenol, d-limonene, hederagenin, indole-3-acetic-acid, rutin and thymol [30].
In present study an attempt was made to optimize the different extraction parameters using supercritical CO2 method to obtain a maximum yield of a fraction of N. sativa seed extract with potent anti-angiogenic activity. Further a series of investigations were conducted using human umbilical vein endothelial cells to elucidate the mechanism of action of the most effective fraction of SC-CO2 extract of N. sativa. Finally, a chemometric correlation between the activity and the chemical composition of the active fraction was done using UV, FTIR and GC-MS analysis.
Pant material and reagents
Seeds of N. sativa were collected from Seiyun city in Yemen. Its herbarium voucher specimen number is (11221- Nigella sativa- 22/3/2011) as authenticated by a Senior Botanist Mr. Shanmugan, School of Biological Sciences, Universiti Sains Malaysia, Penang. The seeds were washed and dried. The dried seeds were ground mechanically and stored in a desiccator. Supercritical carbon dioxide extractor (SFX- 220 SFE system) was purchased from ISCO, USA. Infinite M200 PRO micro-plate reader was obtained from Tecan Group Ltd., Switzerland. Gas chromatograph HP 6890N (G1530N) (HP, China) which accompanied with HP 5973 (G2579A) quadrupole mass spectrometer (Agilent Technologies, USA). Commercial liquid carbon dioxide gas with purity of 99 g/kg in a gas cylinder at a temperature below (-5°C) was supplied locally from Malaysian Oxygen Company, Penang, Malaysia. Dimethyl sulfoxide (DMSO), gallic acid, thymoquinone (TQ), suramin and betulinic acid were obtained from Sigma-Aldrich, USA. MTT (3-(4,5-Dimethylthiazol-2-yl)- 2,5diphenyl tetrazolium bromide) and DPPH (2,2-diphenyl-1-picrylhydrazyl) reagents were purchased from Sigma-Aldrich, Germany. MatrigelTM (10 mg/mL) was obtained from BD Bioscience, USA. The growth medium M199 was obtained from Gibco/Life technology, UK.
Cell line and cell culture
Human umbilical vein endothelial cell line (HUVECs) were purchased from American Type Culture Collection (ATCC), Rockville, MD, USA (Cat No. 8000). They were maintained in ECM medium, supplemented with 5% heat-inactivated fetal bovine serum, 1% penicillin/streptomycin solution and 1% endothelial cell growth factor supplements. ECM and its supplements were also purchased from (ScienCell, USA). Cell culture work was performed in sterilized conditions in a class II Biosafety cabinet and an incubator adjusted with 5% CO2-humidifed atmosphere at 37°C.
Experimental animals
Eight to twelve weeks old Sprague Dawley male rats were obtained from the animal house facility from the School of Pharmaceutical Sciences at Universiti Sains Malaysia. The animals were kept in a clean and quiet transit room with adequate lighting system, temperature and ventilation for few days for acclimatization before euthanasia. The animal work was approved by the Animal Ethics Committee of Universiti Sains Malaysia. The approval reference was USM/Animal Ethics Approval/2010/(59) (228) from Universiti Sains Malaysia.
Supercritical carbon dioxide extraction (SC-CO2)
Twelve extracts were prepared using the SFX-220 SFE system, by varying extraction pressure (2500, 3000, 4500 and 6000 psi) and temperature (32, 45 and 60°C) for 60 min dynamic according to the SC-CO2 extraction method as described before [14]. The extracts were coded as (A1, A2, A3, B1, B2, B3, C1, C2, C3, D1, D2 and D3) and percentage yield was calculated.
Anti-proliferation assay
MTT assay [31] was used to evaluate the anti-proliferative effect of the SC-CO2 extracts of N. sativa. Briefly, HUVECs were treated with various concentrations (5 to 100 μg/mL) of the extracts for 48 h. Betulinic acid and thymoquinone were used as positive controls. DMSO (0.1%) was used as the negative control. After 48 h MTT was added and the optical densities (OD) were measured at 570 nm and reference was 620 nm. The results were presented as mean ± SEM percent inhibition of cell proliferation.
Rat aortic ring assay
This assay was carried out as previously described Goodwin with minor modifications [32]. Thoracic aortas were removed from euthanized male rats, rinsed with serum free medium and cleaned from fibroadipose tissues. The aortas were cross sectioned into small rings (approximately 1 mm thickness) and seeded individually in 48-wells plate in 300 μL serum free M199 media containing 3 mg/mL fibrinogen and 5 mg/mL aprotinin. Ten microliters of thrombin (50 NIH U/mL in 1% bovine serum albumin in 0.15 M NaCl) was added into each well and incubated at 37°C for 90 min to solidify. A second layer (M 199 medium supplemented with 20% HIFBS, 0.1% έ-aminocaproic acid, 1% L-Glutamine, 2.5 μg/mL amphotericin B, and 60 μg/mL gentamicin) was added into each well (300 μL/well). Various concentrations (25- 250 μg/mL) were used for calculating IC50 of the active extracts (A3, B3 C3 and D3). Suramin (100 μg/mL) and thymoquinone (5-50 μg/ mL) were used as positive controls whereas, 0.1% DMSO was used as a negative control. On day four, the medium was replaced with a fresh one containing the test materials. On day five, aortic rings were photographed using EVOS f1 digital microscope (Advanced Microscopy Group, USA) (40X magnification) and subsequently the length of blood vessels outgrowth from the primary tissue explants was measured using Leica Quin software.
The inhibition of blood vessels formation was calculated using the formula
% blood vessels inhibition=(1-(A0/A)Z) × 100, Where; A0=distance of blood vessels growth in treated rings in μm, A=distance of blood vessels growth in the control in μm.
The results are presented as mean percent inhibition ± SD, (n=8). % inhibition was plotted against the concentrations and IC50 was calculated.
The significant difference between the micro vessels out growth in treated versus untreated aortic rings was calculated using Student’s t test. Based on the results of this assay, the most active extracts, A3 and B3 were chosen for the subsequent investigations for the antiangiogenic property.
Evaluation of antiangiogenic properties of A3 and B3
Colony formation assay: The assay was performed according to Franken et al. [33], HUVEC cells were cultured in 6 well plate using ECM medium (500 cell/mL) for 12 h. The cells were treated with 10, 20, 40 and 60 μg/mL of the extracts A3 and B3. Betulinic acid (20 μg/ mL) and DMSO (0.1%) were used as positive and negative controls, respectively. The treatment was carried out for 48 h by incubating the plates in incubator with 5% CO2-humidifed atmosphere at 37°C. The cells were maintained by replenishing the old medium with fresh ECM for every 3 to 4 days until sufficient number of colonies (≥ 50 cells) were recorded in the negative control group (usually 9-10 days). Eventually, the colonies were fixed, stained with 0.2% crystal violet and counted under stereomicroscope. Percentage of plating efficiency (PE%) and percentage of surviving fraction (SF%) was calculated.
Endothelial cell migration assay: The assay was conducted as described by Ref. [34]. The HUVECs were seeded in ECM and incubated for 48 h. The monolayer was scratched with a sterile 200 μL micropipette tip and then washed with PBS to remove the unattached cells and to smoothen the edges of the scratch. ECM (2 mL) with low serum concentration (2.0%) was added to each well followed by addition of two different concentrations (20 and 40 μg/mL) of A3 and B3. Whereas, 0.1% DMSO and betulinic acid (20 μg/mL) were used as the negative and positive controls, respectively. For each test group, 10 microscopic fields per well were photographed at 0, 12 and 24 h using AMG EVOS fI inverted microscope. The width of the wounded area was measured using Leica Quin software. The percent inhibition of cell migration was then calculated relative to wound closure at zero time using the formula:
% wound closure=(1–the width at the indicated times (h)/the width at zero time) × 100%
The results are displayed as average ± SD, (n=6).
Tube formation assay: Inhibitory effect of A3 and B3 on tube-like networks of HUVECs on matrigel matrix was investigated as previously described [35]. In brief, a layer of matrigel (150 μL/well) was added to 48 well plate and allowed to solidify for 45 min in an incubator at 37°C and 5% CO2. Then, HUVECs were seeded onto the matrigelcoated plate and treated with three concentrations (20, 40 and 60 μg/ mL) of A3 and B3 for 24 h. The plate was incubated in CO2 incubator at 37°C and 5% CO2. Betulinic acid (10 μg/mL) and (0.1%) DMSO was used as positive and negative controls, respectively. Finally, the tubular networks in the wells were photographed using AMG EVOS fI inverted microscope and the network length and width was quantified [36].
Measuring of VEGF expression:Vascular endothelial growth factor (VEGF) activity was measured using human VEGF-165 ELISA kit (Raybio, USA) according to the manufacturing instructions. HUVECs were treated over night with the extracts A3 (20 μg/mL) and B3 (20 μg/ mL). DMSO (0.1%) was used as a negative control. The concentration of VEGF in the HUVECs was calculated from the regression equation as pg/mL of cell lysates.
Chick embryo chorioallantoic membrane (CAM) assay: Inhibitory effect of A3 and B3 was investigated in vivo using fertilized chicken embryo [37]. Fertile eggs were cleaned and kept horizontally with rotation for 3-5 days at 37°C in a humidified incubator to ensure formation of chorioallantoic membrane (CAM) vessel network. Subsequently, a small window was made gently with sterilized seizer on the upper face of the eggshell. About 5 mL of albumin was aspirated carefully and the eggs were incubated horizontally to allow the CAM to detach from the shell. The SC-CO2 extracts A3 and B3 were prepared in agarose 1.2% discs at 100 μg/disc and applied directly onto the CAM. Discs containing the vehicle only (1.2% agarose) were used as negative control, while 50 μg/disk of Suramin was used as a positive control). The opening was covered with sterilised surgical tape and the embryos were incubated for 48 h. At the end, the CAMs at the disk area were photographed under a dissecting microscope and blood vessels in each CAM were counted. The results are presented as a mean blood vessel count per CAM ± SD, (n=6).
Phytochemical analysis
The twelve extracts were analysed using FTIR, UV-Vis spectrophotometry, total phenolic and flavonoid contents. These assays and their results were published in our previous article [14].
GC-MS
GC-MS analysis of the extracts was carried out on a gas chromatograph HP 6890N (G1530N) which was accompanied with HP 5973 (G2579A) quadrupole mass spectrometer, at 70 eV. Non polar capillary column HP-5MS 19091S-433 was used as a stationary phase. The initial oven temperature was 70°C (two min) and then increased to 285°C at a rate of 20°C/min. The total run time was 32.75 min. Helium flow rate was 1.2 mL/min. The source temperature was 230°C while the quadrupole temperature was 150°C. Then, mass spectrometer recorded mass spectra and mass/charge ratios (m/z) of molecular ions and compared them to the referenced data of NIST02 library of MSD ChemStation Data analysis application [38].
Statistical analysis
The results were expressed as means ± SD. They were analysed statistically using SPSS 16.0 package by One-way ANOVA test. Significant level was considered as (0.05). Correlation was calculated by bivariate, 2-taild pearson test; R2 values were calculated by linear regression test.
The dried powder of N. sativa seeds were extracted with SC-CO2 extraction method. Totally twelve (12) extracts (A1, A2, A3, B1, B2, B3, C1, C2, C3, D1, D2 and D3) were resulted from different experimental conditions in accordance with the full factorial design with variables such as extraction time, pressure and temperature. The details of the extracts are given in Table 1. All the extracts were first screened for antiangiogenic property using anti-proliferative assay on HUVECs and anti-neovascularization assay on rat aortic explants. Figure 1 illustrates the GCMS chromatograms.
| Extract | SC-CO2 extraction parameters | |
|---|---|---|
| Pressure (psi) | Temperature (°C) | |
| A1 | 2500 | 32 |
| A2 | 2500 | 45 |
| A3 | 2500 | 60 |
| B1 | 3000 | 32 |
| B2 | 3000 | 45 |
| B3 | 3000 | 60 |
| C1 | 4500 | 32 |
| C2 | 4500 | 45 |
| C3 | 4500 | 60 |
| D1 | 6000 | 32 |
| D2 | 6000 | 45 |
| D3 | 6000 | 60 |
Table 1: Codes of the SC-CO2 extracts of N. sativa according to the extraction parameters of pressure and temperature.
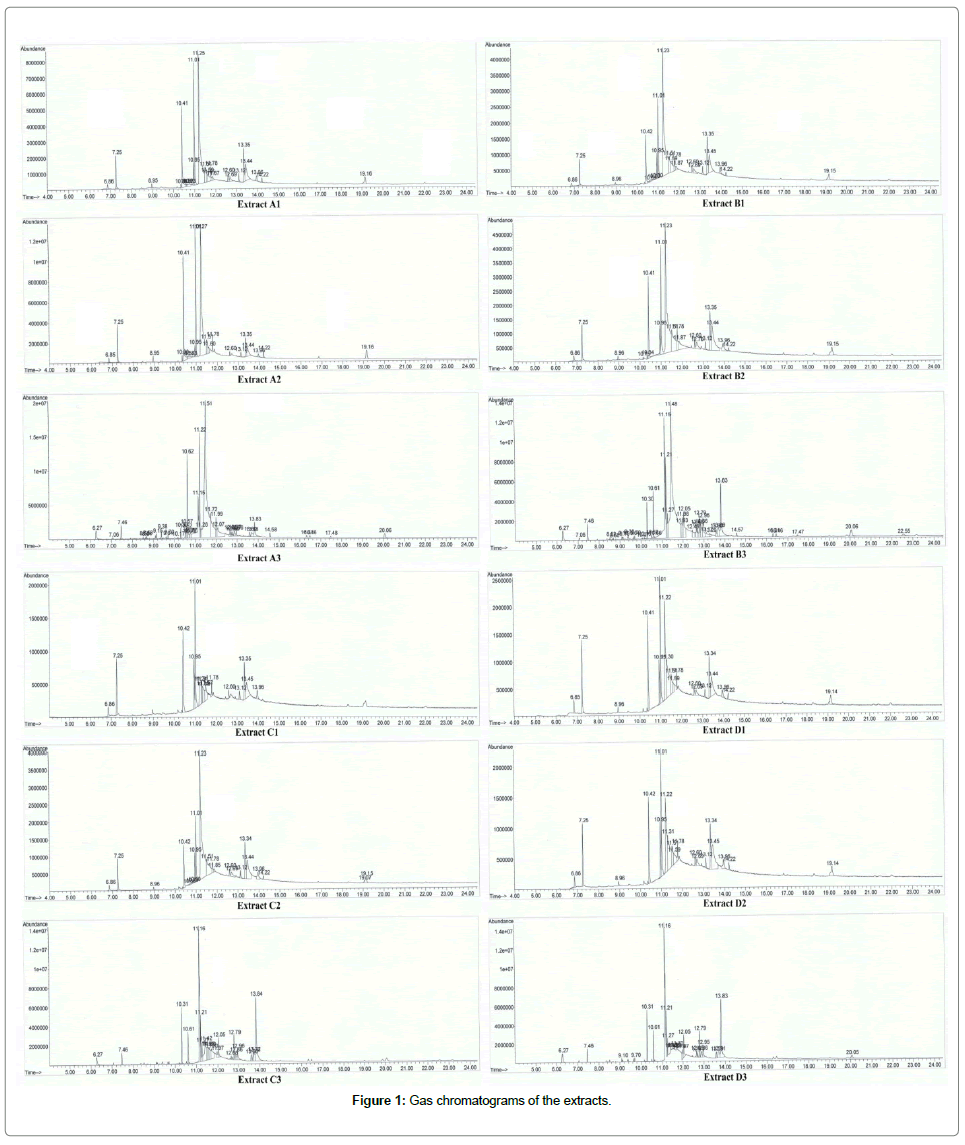
Figure 1: Gas chromatograms of the extracts.
Rat aortic ring assay
Table 2 shows that A3 and B3 showed significant (P<0.05) antiangiogenic effect with IC50 90.75 ± 4.75 and 98.60 ± 2.91 μg/mL, respectively whereas, C3 and D3 demonstrated a moderate activity with IC50 109.40 ± 3.53 and 125.44 ± 7.93 μg/mL, respectively. The dose-response curves of the active extracts were depicted in Figure 2c.
| Extract | IC50 (μg/mL) |
|---|---|
| A3 | 90.75 ± 4.75* |
| B3 | 98.60 ± 2.91* |
| C3 | 109.40 ± 3.53 |
| D3 | 125.44 ± 7.93 |
| TQ | 12.69 ± 1.44* |
Table 2: IC50s (μg/mL) of A3, B3, C3, D3 and TQ on inhibition of neovascularization (rat aortic ring assay).
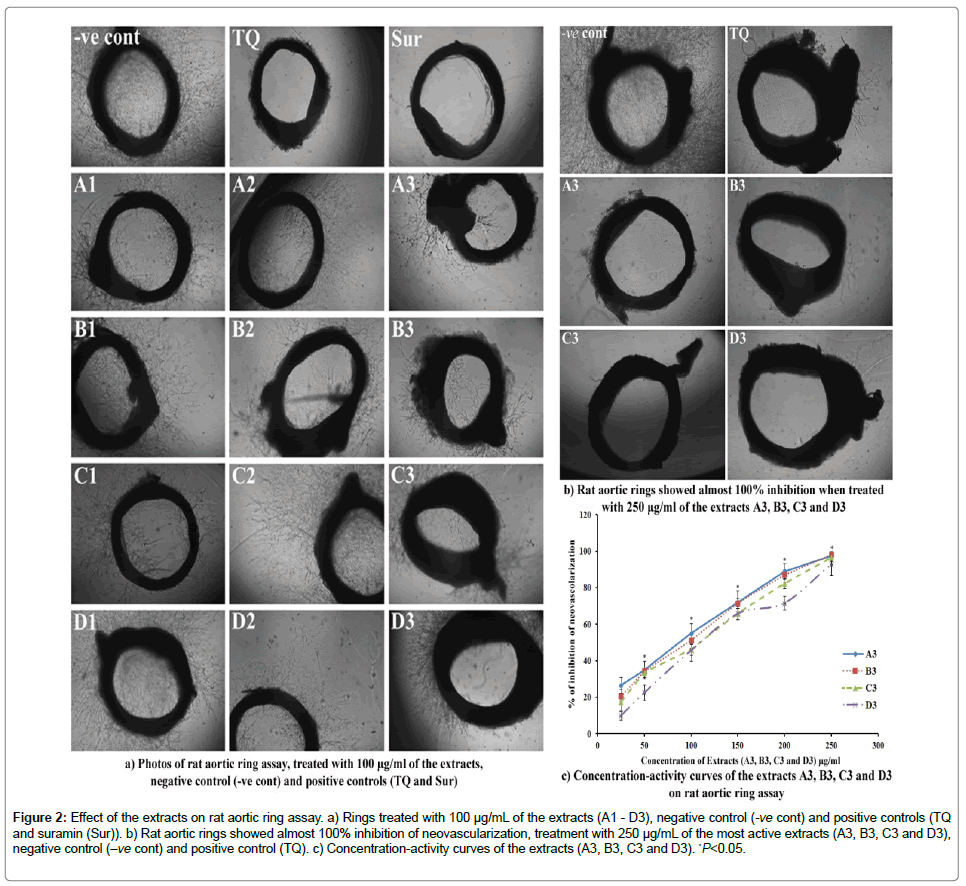
Figure 2: Effect of the extracts on rat aortic ring assay. a) Rings treated with 100 μg/mL of the extracts (A1 - D3), negative control (-ve cont) and positive controls (TQ and suramin (Sur)). b) Rat aortic rings showed almost 100% inhibition of neovascularization, treatment with 250 μg/mL of the most active extracts (A3, B3, C3 and D3), negative control (-ve cont) and positive control (TQ). c) Concentration-activity curves of the extracts (A3, B3, C3 and D3). *P<0.05.
Table 3 illustrates that at a concentration of 100 μg/mL, the extracts A3, B3, C3 and D3 showed the more pronounced inhibitory effect on neovascularization in rat aortic explants. The percentage inhibition of A3, B3, C3 and D3 at 100 μg/mL was 54.38 ± 5.17, 46.88 ± 5.18, 49.10 ± 6.32 and 40.27 ± 8.35%, respectively. However, TQ and suramin showed almost complete inhibition of sprouting of micro vessels from the aortic explants. The inhibitory effect of the test samples on the sprouting of micro-vessels from the aortic rings are depicted in Figure 2a and 2b.
| Extract | EC50 on DPPH (mg/mL) | % of HUVECs cell viability (MTT) | % of inhibition on rat aortic ring assay |
|---|---|---|---|
| A1 | 1.93 ± 0.05 | 89.77 ± 4.39* | 14.48 ± 4.31 |
| A2 | 2.11 ± 0.13 | 67.86 ± 3.43* | 21.74 ± 6.99 |
| A3 | 4.47 ± 0.16 | (-0.22) ± 0.70* | 54.38 ± 5.17* |
| B1 | 1.86 ± 0.02 | 101.03 ± 2.31 | 12.68 ± 4.79 |
| B2 | 1.90 ± 0.04 | 112.69 ± 0.55 | 20.42 ± 4.37 |
| B3 | 4.29 ± 0.07 | (-0.14) ± 0.24* | 46.88 ± 5.18* |
| C1 | 1.58 ± 0.04 | 106.24 ± 1.64 | 13.71 ± 4.33 |
| C2 | 1.67 ± 0.03 | 105.66 ± 0.78 | 9.63 ± 5.62 |
| C3 | 4.13 ± 0.09 | 103.18 ± 0.82 | 49.10 ± 6.32* |
| D1 | 0.86 ± 0.04 | 98.97 ± 1.24 | 17.61 ± 6.76 |
| D2 | 1.50 ± .04 | 85.23 ± 1.14* | 26.16 ± 8.33 |
| D3 | 3.27 ± 0.07 | 89.20 ± 2.56* | 40.27 ± 8.35* |
| TQ | (17.46 ± 1.40) × 10-3 | - | - |
| GA | (1.08 ± 0.01) × 10-3 | - | - |
| TQ (50 μg/mL) | - | - | 100.04 ± 3.73 |
| Suramin (100 μg/mL) | - | - | 99.80 ± 4.24 |
Table 3: Activities of the extracts on DPPH free radical as EC50 (mg/mL), percentages of cell viability of HUVEC cells treated with 100 μg/mL of the extracts, and the potencies of the extracts on inhibition of sprouting of microvessel growth on rat aortic ring assay after treatment with 100 μg/mL of extract.
The extracts, A3 and B3 were selected to elucidate the mechanism of action with a series of in vitro and in vivo antiangiogenic assays.
Antioxidant activity
Effect of the SC-CO2 extracts on DPPH scavenging effect is depicted in Table 3. Figure 3 illustrates the effect of different extraction pressure and temperature on the antioxidant effect.
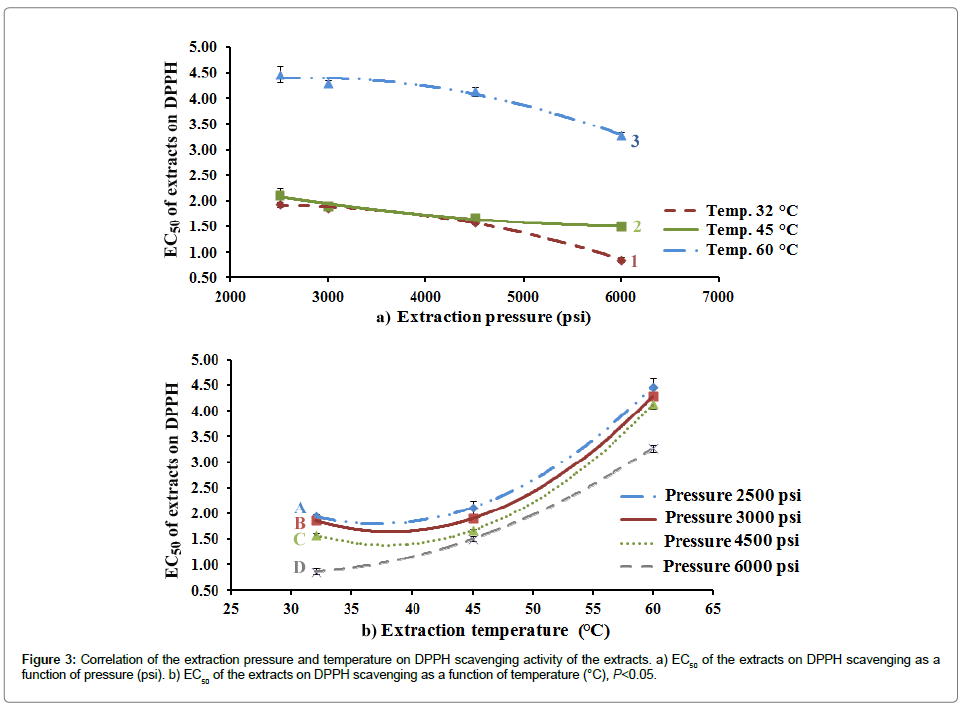
Figure 3: Correlation of the extraction pressure and temperature on DPPH scavenging activity of the extracts. a) EC50 of the extracts on DPPH scavenging as a function of pressure (psi). b) EC50 of the extracts on DPPH scavenging as a function of temperature (°C), P<0.05.
Cell viability assay
The SC-CO2 extracts of N. sativa seeds were tested for their inhibitory effect on HUVECs proliferation using MTT assay. The extracts that were prepared at 60°C with different pressures A3 and B3 (2500 and 3000 psi, respectively) displayed most potent activity as compared to the other extracts tested. The extracts A3 and B3 inhibited proliferation of HUVECs completely at dose of 100 μg/mL. The extracts A3 and B3 exhibited dose-dependent anti-proliferative effect with IC50 41.50 ± 1.92 and 60.21 ± 2.19 μg/mL, respectively (Figure 4). The IC50s of the positive controls, TQ and betulinic acid were 2.39 ± 0.18 and 3.73 ± 0.20 μg/mL, respectively. The result obtained in MTT assay for the SC-CO2 extracts of N. sativa seed is given in the Table 3.
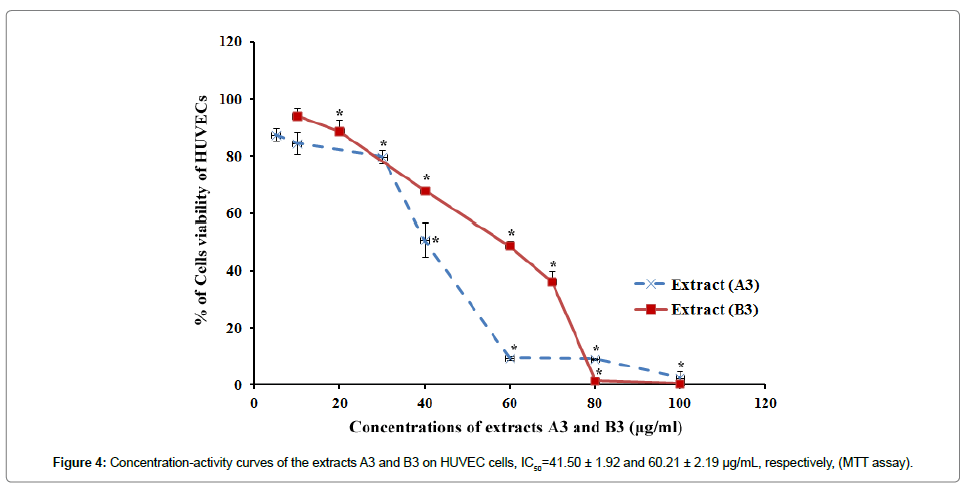
Figure 4: Concentration-activity curves of the extracts A3 and B3 on HUVEC cells, IC50=41.50 ± 1.92 and 60.21 ± 2.19 μg/mL, respectively, (MTT assay).
Antiangiogenic properties of A3 and B3
Colony formation assay: The dose-dependent inhibitory effect of A3 and B3 on colony formation of HUVECs is shown in Figure 5a. Percentage of plating efficiency (PE) in vehicle (0.1% DMSO) treated cells was 88 ± 2.4% which was drastically reduced by the treatment of A3 and B3 with PE 34 ± 1.7 and 21 ± 3.1%, respectively. The results showed that A3 displayed significantly higher inhibitory effect than B3. The surviving fraction (SF) was determined (Figure 5b) to be 14.97 ± 1.37 and 54.22 ± 2.71% after A3 and B3 treatment at the concentrations 20 μg/mL, respectively (Figure 5b). These results can be compared with that of the standard reference, betulinic acid (20 μg/mL) which produced the PE of 11.13 ± 0.82%.
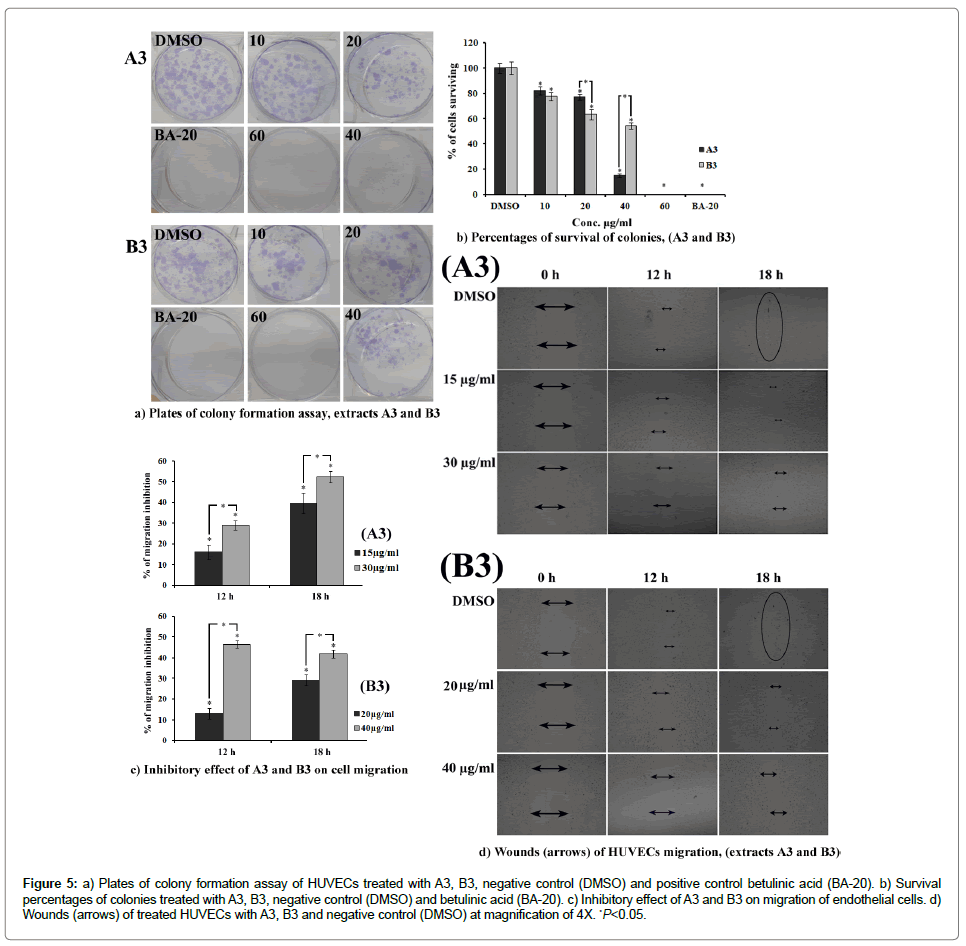
Figure 5: a) Plates of colony formation assay of HUVECs treated with A3, B3, negative control (DMSO) and positive control betulinic acid (BA-20). b) Survival percentages of colonies treated with A3, B3, negative control (DMSO) and betulinic acid (BA-20). c) Inhibitory effect of A3 and B3 on migration of endothelial cells. d) Wounds (arrows) of treated HUVECs with A3, B3 and negative control (DMSO) at magnification of 4X. *P<0.05.
HUVECs cell migration: A3 and B3 exhibited an obvious inhibition of migration of HUVECs. Both the extracts demonstrated time and dose-dependent inhibitory effect on migration of HUVECs. The extracts produced significant inhibition in cell motility (Figure 5c) at the concentration lower than its IC50 on proliferation of HUVECs. The percent inhibition of cell migration for A3 (30 μg/mL) calculated after 12 and 18 h was 29.04 ± 2.34 and 52.35 ± 2.65%, respectively (p<0.05). However, B3 resulted in 46.42 ± 1.79 and 41.88 ± 1.95% of inhibition at 12 and 18 h, respectively. Interestingly, at lower concentration (15 μg/ mL) A3 exhibited significant anti- migratory effect with (P<0.05). Figure 5d clearly depicts the comparative inhibitory effects of the extracts A3 and B3 on migration of endothelial cells with that of untreated cells.
Inhibition of tube formation: The extracts A3 and B3 showed strong abolishing effect on capillary tubules formation of HUVECs. At concentrations 40 and 60 μg/mL, A3 produced 27.06 ± 2.78 and 48.29 ± 0.87% inhibition, respectively. Similarly, B3 exhibited significant inhibitory effect on tube formation with 41.47 ± 2.70 and 57.86 ± 1.12% at 40 and 60 μg/mL concentration, respectively. The results can be compared with that of the positive control, betulinic acid as shown in Figure 6a and 6b.

Figure 6: a) Percentages of inhibition of tube formation of HUVECs by A3, B3, betulinic aicd (BA-10) and negative control (DMSO). b) Images of tube formation assay of HUVECs treated with A3, B3, betulinic acid (BA-10) and negative control (DMSO<0.5%), magnification was 4X. c) Content of VEGF (pg/mL of lysate) expressed by HUVECs treated with A3, B3 and negative control (DMSO). d) Photos of CAM assay after treatment (circles) with 100 μg/disk of A3 and B3. 50 μg/disk of Suramin (+ve cont) and untreated CAM (–ve cont). *P<0.05.
Inhibition of VEGF expression: In human VEGF-165 ELISA assay, HUVECs were treated separately with 20 μg/mL of each extract (A3 and B3) for 24 h treatment. The results showed remarkable inhibition of VEGF-165 expression in HUVECs by both the extracts. Content of VEGF-165 in vehicle (0.1% DMSO) treated cells was recorded up to 252.45 ± 7.34 pg/mL, which was drastically reduced to 77.18 ± 8.43 and 67.8 ± 5.25 pg/mL after treatment with A3 and B3, respectively. Figure 6c illustrates the comparative effects of A3 and B3.
CAM assay: Treatment of fertilized chick embryo with A3 or B3 produced a significant deformed vascular architecture. The inhibitory percentage of 100 μg/disk of A3 was 63.67 ± 2.55% which was significantly higher than that of B3 (51.75 ± 3.68%). The positive control (50 μg/disk Suramin) showed 63.47 ± 4.24% inhibitions. Figure 6d shows normal vasculature pattern in the vehicle (1.2% agarose) treated CAMs with prominent primary, secondary and tertiary vessels and dendritic branching pattern. Whereas, a distorted architecture in the vasculature can be seen clearly in the treated CAMs with the extracts (A3 and B3). The results of blood vessel count in treated CAMs showed that the treatment reduced the number of the blood vessels drastically.
GC-MS: The SC-CO2 extract of N. sativa seeds prepared in the present study were analysed using GC-MS to specify and quantify the major chemical constituents present in them. Figure 1 depicts the GC-MS chromatograms of the SC-CO2 extracts. The quantified data presented in the Figure 1 that illustrates the composition and proportion of the major chemical constituents in the extracts. The parameters such as retention time, % area peak, molecular formula and molecular weight obtained from the GC-MS of the extracts are given in Table 4. Detailed characteristics of the peaks identified in the GC-MS were described in the Table 4. Further GC-MS analysis of active and inactive extracts depicted in Tables 5 and 6, which represent the compounds present in active extracts and the compounds commonly, found in all the extracts. Mass spectra for each peak in the chromatogram are given in the Supplementary Information.
| Peak no. | RT min | Compounds | Molecular formula | Molecular weight |
|---|---|---|---|---|
| 1 | 6.27 | TQ | C10H12O2 | 164.08 |
| 2 | 6.86 | (+)-α-longipinene (sesquiterpene) | C15H24 | 204.19 |
| 3 | 7.25 | (+)-longifolene (sesquiterpene) | C15H24 | 204.19 |
| 4 | 8.36 | Thymohydroquinone | C10H14O2 | 166.1 |
| 5 | 8.52 | Cetane (hexadecane) | C16H34 | 226.27 |
| 6 | 9.1 | Heptadecane | C17H36 | 240.28 |
| 7 | 9.13 | Pristane (norphytane) (terpenoid) | C19H40 | 268.31 |
| 8 | 9.65 | Octadecane | C18H38 | 254.3 |
| 9 | 9.7 | Phytane | C20H42 | 282.33 |
| 10 | 10.17 | Eicosane | C20H42 | 282.33 |
| 11 | 10.15 | Methyl palmitate (metholene 2216) | C17H34O2 | 270.26 |
| 12 | 10.41 | Squalene (triterpene) | C30H50 | 410.39 |
| 13 | 10.59 | Palmitic acid (fatty acid) | C16H32O2 | 256.24 |
| 14 | 10.95 | Methyl octadeca-8,11-dienoate | C19H34O2 | 294.26 |
| 15 | 11.28 | Methyl stearate | C19H38O2 | 298.29 |
| 16 | 11.25 | Linoleic acid (linolic acid) (fatty acid) | C18H32O2 | 280.24 |
| 17 | 11.51 | 9-Eicosyne | C20H38 | 278.3 |
| 18 | 11.78 | Androstane-3,17-diol ((3α,5α)-androstane-3,17-diol) (neurosteroid) | C19H32O2 | 292.24 |
| 19 | 12.05 | Methyl (11E,13E)-11,13-icosadienoate | C21H38O2 | 322.29 |
| 20 | 13.29 | Hexacosane (cerane) | C26H54 | 366.42 |
| 21 | 13.82 | 2-Linoleoyl glycerol (monolinolein) | C21H38O4 | 354.28 |
| 22 | 14.22 | trans-squalene (E,E,E,E-squalene) | C30H50 | 410.39 |
Table 4: GC-MS data of the extracts by comparison of experimental with referenced data of NIST02 library.
| No. | Extract | Compound | % of Area |
|---|---|---|---|
| 1 | A3, B3, C3 and D3 | TQ | 1.20-2.10 |
| 2 | A3, B3, C3 and D3 | Cetane | 0.14-0.24 |
| 3 | A3, B3, C3 and D3 | Octadecane | 0.11-0.21 |
| 4 | A3, B3, C3 and D3 | Phytane | 0.22-0.30 |
| 5 | A3, B3, C3 and D3 | Methyl stearate | 1.13-2.30 |
| 6 | A3 and B3 | 2-Linoleoyl glycerol | 2.51-6.52 |
| C3 and D3 | 2-Linoleoyl glycerol | 12.60-16.13 | |
| 7 | A3 | Eicosane | 0.60 |
| 8 | B3 | Hexacosane | 0.26-0.38 |
| 9 | B3 | Thymohydroquinone | 0.19 |
| C3 and D3 | Thymohydroquinone | 0.56-0.62 | |
| 10 | B3, C3 and D3 | Heptadecane | 0.14-0.25 |
| 11 | B3, C3 and D3 | Methyl (11E,13E)-11,13-icosadienoate | 2.55-4.75 |
| 12 | C3 and D3 | Pristane | 0.28-0.30 |
Table 5: Compounds that were contained in the most active extracts (A3, B3, C3 and D3).
| No. | Extract | Compound | % of area |
|---|---|---|---|
| 1 | A3, B3, C3 and D3 | (+)-α-longipinene | 0.12-0.36 |
| A1, A2, B1, B2, C1, C2, D1 and D2 | (+)-α-longipinene | 0.32-1.51 | |
| 2 | A3, B3, C3 and D3 | (+)-Longifolene | 0.90-1.41 |
| A1, A2, B1,B2 and C2 | (+)-Longifolene | 1.77-2.95 | |
| C1, D1 and D2 | (+)-Longifolene | 5.37-7.76 | |
| 3 | A3 and B3 | Methyl palmitate | 0.51-1.50 |
| C3 and D3 | Methyl palmitate | 3.29-4.38 | |
| A1, A2 and B2 | Methyl palmitate | 0.11-0.21 | |
| 4 | A3 | Palmitic acid | 1.1 |
| A1 and A2 | Palmitic acid | 0.47-1.29 | |
| 5 | A3 and B3 | Methyl octadeca-8,11-dienoate | 7.86-13.45 |
| C3 and D3 | Methyl octadeca-8,11-dienoate | 21.26-21.55 | |
| A1, A2, B1, B2, C1, C2, D1 and D2 | Methyl octadeca-8,11-dienoate | 1.65-6.17 | |
| 6 | All 12 extracts | Linoleic acid | 33.97-44.13 |
| 7 | C3 and D3 | 9-Eicosyne | 2.44-6.01 |
| B1, C2, D1 and D2 | 9-Eicosyne | 1.68-11.79 | |
| 8 | A3 | Androstane-3,17-diol | 1.92 |
| A1 and A2 | Androstane-3,17-diol | 2.59-2.86 | |
| 9 | A3 and B3 | trans-squalene | 0.24-0.34 |
| A2 | trans-squalene | 0.56 | |
| 10 | A2 | Squalene | 8.24 |
Table 6: Compounds that were found in all the extracts commonly.
In the present study, an anti-angiogenic property of 12 SC-CO2 extracts of N. sativa seeds was evaluated. The anticancer effect of these extracts was measured by MTT test on five cancerous human cell lines: MCF 7, MDA-MB-231, HCT 116, Hep G2 and PC-3, as well as one normal human cell lines: CCD-18Co. It showed that the extract A3, which was extracted at the highest temperature (60°C) and the lowest pressure (2500 psi), was the most potent extract against breast cancer (MCF 7) with IC50 value of 53.34 ± 2.15 μg/mL as we reported previously [14]. The oil of N. sativa has showed significant anti-tumour potency [39,40]. Thymoquinone (TQ), the most active component of N. sativa, has resulted in an obvious anti-proliferative effect against HUVEC cell line. Also, it inhibited endothelial cell migration, invasion, tube formation and VEGF expression [41].
The anti-proliferative activity of the SC-CO2 extracts against HUVECs was reported. It showed that two extracts which were extracted at the higher temperature (60°C) and the lower pressures, A3 (2500 psi) and B3 (3000 psi) represented the most potencies. A3 (IC50 of 41.50 ± 1.92 μg/mL) showed stronger activity than B3 (IC50 of 60.21 ± 2.19 μg/mL) as tested by MTT assay. Angiogenic activity of the 12 different extracts of SC-CO2 of N. sativa revealed that four extracts (A3, B3, C3 and D3) significantly thwarted the micro vessel outgrowth as tested using ex-vivo isolated tissue assay. When the extraction pressure was increased, the level of antiangiogenic activity decreased. Closer inspection of the constituents present in these extracts indicates the presence of thymo-quinone and thymo-hydroquinone. The other eight extracts did not show promising anti-angiogenic effect and thymoquinone, thymo-hydroquinone was not detected.
The in vitro anti-angiogenic activities of A3 and B3 were evaluated by quantifying their inhibitory effect on endothelial cell proliferation, colonization, migration, tube formation and on VEGF expression. These assays provide a valuable data for anti-angiogenesis assessment [37,42]. Endothelial cells tend to be in contact with their neighbouring cells. These population characteristics lead to formation of colonies and enhance angiogenesis [43]. A3 and B3 inhibited colony formation by HUVECs in dose-response manner. This confirms the cytotoxic effect of the two extracts on HUVECs at higher concentration, whereas the lower concentrations showed cytostatic effect. The higher concentrations of both extracts resulted in irreversible inhibition of the cells viability. A3 showed stronger activity than B3 especially at 60 and 40 μg/mL. Endothelial cell migration and invasion are two integrated pathways required for normal embryonic morphogenesis, tissue regeneration in adults and they play important role in cancer development and tumour growth and neovascularization [44,45]. Migration signalling pathway is mediated by many interlinked positive feedback loops, such as Rho family GTPases, phosphoinositide 3-kinases, integrins, angiostatins and endostatins [46,47]. A3 and B3 elucidated potent suppression of HUVEC cell migration, especially after 18 h, where the untreated cells (DMSO) caused total closure of the scratch wounds. Previous study that looked into the effect of TQ reported inhibitory effect of the compound on endothelial cell migration [41]. in vitro tube formation assay demonstrates differentiation of HUVECs; it mimics in vivo model of angiogenesis [48]. The extracts A3 and B3 significantly altered the architectural capillary tube networks in dose-dependent manner. B3 showed more efficient inhibition of tube formation, where 60 and 40 μg/mL of A3 resulted in 48.29 ± 0.87 and 27.06 ± 2.78% of inhibition, respectively. Where as, 60 and 40 μg/mL of B3 resulted in 57.86 ± 1.12 and 41.47 ± 2.70% of inhibition. It has been reported that TQ resulted in a significant inhibition of endothelial cell tube formation [41]. Measuring of VEGF expression is one of the important methods for evaluating angiogenesis inhibition. There are many isoforms of VEGF, such as VEGF-121, VEGF-165 and VEGF-189, but VEGF-165 plays the predominant effective role in angiogenesis mechanism, its expression is regulated by hypoxic condition of tumour [49,50]. In this study, A3 and B3 stopped angiogenesis cascade, by counteracting VEGF-165 expression significantly. The inhibitory effect of these two extracts on VEGF expression pathway leads to down-regulation of hypoxia [51,52]. The inhibition of VEGF expression prevents binding of HIF-1 with HREs, so it antagonizes hypoxic conditions of tumour [53,54]. This activity of A3 and B3 might be due to their content of TQ, which has showed significant inhibition of VEGF expression [41].
GC-MC data showed that the 12 SC-CO2 extracts of N. sativa seeds resulted in clear differences in their constituents. This explained the reported variation in antiangiogenic activities of the extracts. GCMS qualitative analysis resulted in semi-quantitative identification of twenty-two compounds in the extracts. The most active extracts (A3, B3, C3 and D3) contained many compounds that were detected only in these active extracts at different ratios. They include TQ, cetane, octadecane, phytane, methyl stearate and 2-linoleoyl glycerol. Eicosane was detected in the extract A3 and hexacosane in B3. Three other compounds (thymohydroquinone, heptadecane and methyl (11E,13E)-11,13-icosadienoate) were obviously reported in the extracts B3, C3 and D3. In addition, the extracts C3 and D3 exhibited presence of pristane. Previous studies reported various bioactivities of these compounds, such as cytotoxic and antitumour potential of TQ [21,41], thymohydroquinone [55,56] and pristine [57].
Many studies have revealed the contribution of antioxidants in cancer prevention as apoptosis inducers or as antiangiogenic agents [58,59]. TQ has showed a good scavenging activity for superoxide radical [60]. The results of antioxidant activities of the extracts showed that the most active extract was D1 (EC50=0.86 ± 0.04 mg/mL); while, the least activity was recorded by the extract A3 (EC50=4.47 ± 0.16 mg/mL). The extraction pressure showed a positive correlation with the antioxidant activity significantly. On the other hand, there is a significant negative correlation between the extraction temperature and antioxidant activity. Antioxidant activity of the natural extracts is linked to the presence of phenolic constituents [60,61]. It is suggested that phenolic content has anticancer activity [62-64]. These extracts showed good content of polyphenolics and total flavonoids content as published in our article [14].
The extracts A3 and B3 resulted in a significant inhibition of sprouting of new blood vessels of CAM assay. A3 has significantly higher level of activity (percentage of inhibition was 63.67 ± 2.55%) than B3 (51.75 ± 3.68%) (P<0.05). However the level of TQ in A3 is not significantly different to B3. This suggests that the other compounds present in A3 may also contribute to its in vitro and in vivo antiangiogenic activity. Compounds such as eicosane, palmitic acid and androstane-3,17-diol were present in A3 but absent in B3. The effect of eicosane on angiogenesis has not been thoroughly investigated however palmitic acid has been found to increase angiogenesis by up-regulating IL8 [65], hence may not have contributed towards the antiangiogenic activity by A3. The other compound present is androstane-3,17-diol, where the related compound androsten-3,17-diol has been showed to have anti-inflammatory response by up-regulating IL-4 and IFN-γ. IL-4 and IFN-γ are potent inhibitors of angiogenesis [66,67]. Therefore it is probable that the additional potency in antiangiogenic activity by A3 may be due to the androstane-3,17-diol. Thymoquinone, an active principle of black seeds has resulted in antitumour effects [55] and suppressed proliferation, migration, invasion and tube formation of HUVECs. In addition, TQ restricted neovascularization on rat aortic ring assay [41].
In this study, the oil of N. sativa seeds was extracted using a SC-CO2 extraction method, which was developed to target angiogenesis cascade. Angiogenesis plays important role in tumourigenesis. The study reveals that SC-CO2 extraction can be useful to produce antiangiogenic extracts from N. sativa seeds. It is reported that the antiangiogenesis potency of this extract is highly dependent on extraction temperature and pressure. The higher temperature (60°C) and lower pressure (2500 and 3000 psi) produced the most potent antiangiogenic extracts against HUVECs. TQ is the key compound that has an important role in inhibiting angiogenesis. Another compound androstane-3,17-diol, is suspected to have acted synergistically with TQ to cause selective anticancer activity of the A3 extract. The extracts inhibited angiogenesis in vitro and in vivo by disrupting endothelial cell migration and tube formation by down-regulation of VEGF expression. It caused selective cytostatic activity against endothelial cells. The antiangiogenic activity could be due to the anti-oxidant nature of the extracts.
We would like to thank Universiti Sains Malaysia for supporting this study by a research university team (RUT) grant number 1001/PFARMASI/851001 and the postgraduate student research grant (1001/PFARMASI/834052). Our thanks to the Institute of Health Sciences in Seiyuon – Hadramaut and Ministry of Health in Yemen for awarding the financial scholarship to support this study.