Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Review Article - (2013) Volume 2, Issue 5
Currently, transportation fuels and many platform chemicals are derived primarily from petroleum. The use of this finite resource contributes substantially to the dangerous build-up of greenhouse gases in the earth’s atmosphere, which has led to an increasing demand for sustainably produced fuels and chemicals. Aromatic plants accumulate terpenoids and phenylpropanoids with physicochemical properties similar to those of petrochemical feedstocks. They are also a source of bioactive compounds used by the agrochemical and pharmaceutical industries. Recent successes with engineering the terpenoid pathway indicate that aromatic plants have the potential to play an important role in providing chemical and pharmaceutical feedstocks from renewable sources. In this review, we critically assess the current status of the field and discuss different biotechnological approaches to enhance the accumulation of commercially valuable target terpenoids.
Keywords: Biocrude; Essential oil; Glandular trichome; Metabolic engineering; Oleoresin; Terpene synthase; Terpenoid
Aromatic plants have a distinctive, usually fragrant smell or aroma, which is caused by the emission of volatiles. In this review, we are going to focus on a subset of aromatic plants where specialized anatomical structures for the storage of large volumes of volatile resins or essential oils have evolved. Archaeological evidence indicates that aromatic plants have been employed since pre-historic times as seasonings, herbal medicines and to prevent food spoilage [1-3]. The oldest written records of aromatic plants as sources for medicinal substances date back several thousand years to ancient Egypt, Sumer, Persia, China, India, and Mesoamerica [4]. The earliest systematic compilation of methods for the extraction and distillation of essential oils, the biologically active secretory products of aromatic plants, is found in a pharmacopoeia by Ibn al-Baitar (Arabic: 1198-1248 ??????? ??? ), a pharmacist and physician of the Islamic Golden Age [5]. The use of tar and pitch from pine trees as adhesives predates recorded history. There is even archaeological evidence that birch pitch was processed by Neanderthals more than 80,000 years ago, to manufacture an adhesive used to attach spear points to wooden shafts [6]. Tar and pitch, used for caulking wooden ships, preserving rigging and making torches, were already important commodities of ancient Mediterranean civilizations several hundred years B.C. [7]. Investigations into the chemical components of turpentine, obtained by distillation of conifer resin, were initiated in the 1800s by Durnas, Berthelot, Kékulé, Wallach and Bayer [8]. The naval stores industry then began building commercial plants to fractionate tall oil, which was recovered as a by-product of the Kraft process of conifer wood pulping (developed in the late 1800s), to supplement resin derived from the continued tapping of live pines [7]. Distillation products have had many uses. Turpentine, the highly volatile fraction of conifer wood extraction, has been employed broadly as paint solvent and thinner, component of polishes, as an insecticide, and for the production of specialty chemicals [9]. The less volatile rosin fraction was converted into synthetic resins, paper size (water repellent), soap and varnish [9]. With the emergence of the petrochemical industry in the 1920s, natural gas, coal and petroleum became the most important chemical feedstocks for the production of pharmaceuticals, solvents, fertilizers, pesticides, and plastics [10]. As a result, the naval stores industry declined over the following decades to a small fraction of its former production. However, there is some potential for a partial resurgence. The long-term environmental costs of fossil fuels have become apparent [11] and this has led to a rekindled interest in naval stores. Many industries in developed countries are implementing and developing more sustainable processes for the production of fuels and consumer goods. Aromatic plants have the potential to contribute to this development, especially as sources of chemical and pharmaceutical feedstocks.
The oleoresins and essential oils of aromatic plants, which are comprised of mostly terpenoids and phenylpropanoids, accumulate in particular anatomical structures, including resin ducts, secretory cavities, glandular trichomes, and isolated oil cell idioblasts [12,13]. These plant tissues contain specialized cells that are responsible for the biosynthesis and secretion of aromatic products into a storage cavity. Because of the commercial attractiveness of terpenoid essential oil components, several research groups have attempted to express biosynthetic genes in transgenic plant hosts. This review will use selected examples to outline the potential for using aromatic plants as sources of sustainable chemical and pharmaceutical feedstocks. A more in-depth discussion was provided by Lange and Ahkami [14]. There is relatively little literature on biotechnologically produced phenylpropanoids in aromatic plants, and the focus of this article will thus be on terpenoid pathway engineering. Metabolites are numbered sequentially when they first appear in the text and their structures are shown in Figure 1.
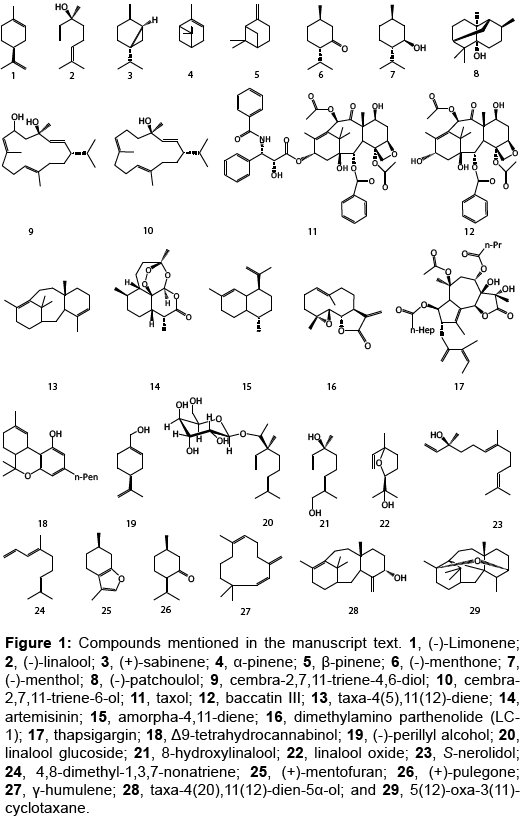
Figure 1: Compounds mentioned in the manuscript text. 1, (-)-Limonene; 2, (-)-linalool; 3, (+)-sabinene; 4, α-pinene; 5, β-pinene; 6, (-)-menthone; 7, (-)-menthol; 8, (-)-patchoulol; 9, cembra-2,7,11-triene-4,6-diol; 10, cembra-2,7,11-triene-6-ol; 11, taxol; 12, baccatin III; 13, taxa-4(5),11(12)-diene; 14, artemisinin; 15, amorpha-4,11-diene; 16, dimethylamino parthenolide (LC- 1); 17, thapsigargin; 18, Δ9-tetrahydrocannabinol; 19, (-)-perillyl alcohol; 20, linalool glucoside; 21, 8-hydroxylinalool; 22, linalool oxide; 23, S-nerolidol; 24, 4,8-dimethyl-1,3,7-nonatriene; 25, (+)-mentofuran; 26, (+)-pulegone; 27, γ-humulene; 28, taxa-4(20),11(12)-dien-5α-ol; and 29, 5(12)-oxa-3(11)- cyclotaxane.
Metabolites with relevance for the chemical industry
According to a recent assessment of the US Department of Energy, roughly 320 million dry tons of biomass per year is currently removed sustainably from US forestlands [15]. As markets for bioenergy feedstocks are evolving, there is considerable potential for harvesting additional biomass from non-reserve forestland by removing logging residues and by strategic thinning [15]. Biomass from conifer trees can contain high quantities of resins, while essential oils are more commonly accumulated by certain species of angiosperms. Both terpenoid oils and resins are characterized by a high volumetric energy density and high degree of reduction. Because of these physicochemical properties, terpenoid resins are viable bio crude feedstocks for heavier combustible liquids comparable to diesel, kerosene and jet fuel [16], while terpenoid oils are usable feedstocks for the manufacturing of solvents, fine chemicals and biopolymers [17,18]. The engineering of plants as renewable sources of specific terpenoid hydrocarbons that can be used in already existing industrial petroleum-based processes is thus highly desirable.
The first committed step in the biosynthesis of terpenoids is catalysed by chain length-specific terpene synthases that release mixtures of terpene hydrocarbons with, in many cases, a predominant terpenoid after which each of these enzymes is named [19] (Figure 2). The terpenoids of relevance for this review are derived from precursors with a chain length of C10 (monoterpenes), C15 (sesquiterpenes) or C20 (diterpenes). The two simplest approaches to produce plants that potentially accumulate elevated levels of hydrocarbons are: 1) overexpression of a particular terpene synthase gene or 2) blocking of a reaction generating oxygenated terpenoids. In the following we will highlight selected examples for both engineering avenues. Ohara et al. [20] transformed tobacco (Nicotiana tabacum L.) with constructs for the expression of a limonene synthase gene (from Perilla frutescens L. Britton) coding for either a cytosolic or plastidial isoform, but found that neither construct yielded significant amounts of the monoterpene limonene (Table 1). The overexpression of plastidtargeted monoterpene synthases, using constitutive promoters, in typical aromatic plants such as peppermint (Mentha x piperita L.) and spike lavender (Lavandula latifolia Medik.), has been more successful. The expression of linalool synthase (from bergamot mint (Mentha citrata Ehrh.)) in transgenic peppermint afforded S-linalool at 0.009% of dry weight biomass (Table 1; Supplementary Information). (+)-Sabinene or a mixture of α-pinene and β-pinene were accumulated to about 0.02% of dry weight biomass in transgenic peppermint plants expressing sabinene synthase (from garden sage (Salvia officinalis L.)) or pine synthase (from Grand fir (Abies grandis (Douglas ex D. Don) Lindley)) (Table 1; Supplementary Information). When limonene synthase (from spearmint (Mentha spicata L.) was expressed in transgenic spike lavender, (-)-limonene levels reached up to 0.09% of the dry weight biomass [21] (Table 1; Supplementary Information). Mahmoud et al. [22] reported that the expression of the gene encoding limonene 3-hydroxylase (L3H), which catalyzes the first oxygenation step of the p-menthane monoterpene pathway in peppermint, was cosuppressed in a transgenic peppermint line that accumulated up to 0.8% of dry weight biomass as (-)-limonene (1) (compared to 0.02% in control plants) [22,23]. This is noteworthy because natural peppermint essential oil is rich in oxygenated monoterpenes, which make up 90% (v/v) of the total oil (prominent examples are (-)-menthone (6) and (-)-menthol (7)). In contrast, the cosuppressed line accumulated 79% (v/v) of the oil as a single monoterpene hydrocarbon ((-)-limonene). Hydrogenated limonene has been employed successfully as an additive to diesel fuel [24] and as a source of fine chemicals [17], indicating that, by combining the metabolically engineered production of terpenoid hydrocarbons in aromatic plants with subsequent chemical processing, it might be possible to provide renewable feed stocks for refineries and chemical plants. Sallaud et al. [25] expressed various genes encoding copalyldiphosphate synthase (NtCPS2) and abienol synthase (NtABS), using a glandular trichome-specific promoter, in flowering tobacco (Nicotiana sylvestris), where the expected product, the diterpene Z-abienol, accumulated to 0.018% of dry weight biomass. This compound, which occurs in cultivated tobacco but not in flowering tobacco, can potentially be used as a precursor for the chemical synthesis of amber-like compounds, which are fairly costly perfumery raw materials. The biotechnological production of Z-abienol-derived chemical feedstocks could be employed as a substitute for the extraction of sclareol, a labdane diterpene accumulated in clary sage, which is currently the most common precursor for the production of amber-like scents.
| Target metabolite | Utility | Gene(s) | Source plant | Host plant | Amount of target metabolite | References | |||
|---|---|---|---|---|---|---|---|---|---|
| of gene | for transformation | [% of oil or exudate] | [% of dry weight biomass] | ||||||
| CONTROL | TRANSGENIC | CONTROL | TRANSGENIC | ||||||
| Metabolites with relevance for the chemical industry | |||||||||
| (-)-Limonene | solvent, biofuel | limonene 3-hydroxylase ↘ (cosuppression) | n.a. | Menthax piperita | 2.2 | 79 | 0.02 | 0.8 | [22] |
| limonene synthase ↗ (plast.) | Menthaspicata | Lavandulalatifolia | - | - | 0.06 | 0.09 | [21] | ||
| limonene synthase ↗ (plast.) | Perillafrutescens | Nicotianatabacum | - | - | 0 | 0.0001 | [20] | ||
| limonene synthase ↗ (cyt.) | Perillafrutescens | Nicotianatabacum | - | - | 0 | 0.000004 | [20] | ||
| S-Linalool | flavor/fragrance, biofuel | linalool synthase ↗ (plast.) | Menthacitrata | Menthax piperita | 0.4 | 0.7 | 0.005 | 0.009 | Suppl. Info |
| limonene 3-hydroxylase ↘ (cosuppression) | n.a. | ||||||||
| (+)-Sabinene | biofuel | sabinene synthase ↗ (plast.) | Salvia officinalis | Menthax piperita | 0.7 | 1.5 | 0.009 | 0.02 | Suppl. Info |
| limonene 3-hydroxylase ↘ (cosuppression) | n.a. | ||||||||
| α- and β-Pinene | solvent, biofuel | pinene synthase ↗ (plast.) | Abiesgrandis | Menthax piperita | 2.2 | 3.6 | 0.01 | 0.02 | Suppl. Info |
| Z-Abienol | flavor/fragrance | copalyldiphosphate synthase ↗ (plast.), | Nicotianatabacum | Nicotianasylvestris | 0 | 0.018 | [25] | ||
| abienol synthase ↗ (plast.) | |||||||||
| Metabolites with relevance for agribusiness industry | |||||||||
| Cembratrienol | disease resistance | cytochrome P450-dependent | Nicotianatabacum | Nicotianatabacum | 1.4 | 27 | 0.2 | 4.3 | [28] |
| oxygenase (cosuppressed) | |||||||||
| Patchoulol | disease resistance | patchoulol synthase ↗ (cyt.) | Pogostemoncabli | Nicotianatabacum | - | - | 0 | 0.005 | [27] |
| patchoulol synthase ↗ (plast.), | Pogostemoncabli | Nicotianatabacum | - | - | 0 | 0.015 | [27] | ||
| farnesyl-PP synthase ↗ (plast.) | Gallus gallusdomest. | ||||||||
| Metabolites with relevance for the pharmaceutical industry | |||||||||
| Taxa-4(5),11(12)-diene | precursor for taxol | taxadiene synthase ↗ (plast.) | Taxusbrevifolia | Physcomitrella patens | - | - | 0 | 0.2 | [37] |
| (anticancer treatment) | taxadiene synthase ↗ (plast.) | Taxusbrevifolia | Nicotianasylvestris | - | - | 0 | 0.005 | [34] | |
| taxadiene synthase ↗ (plast.), | Taxusbrevifolia | Arabidopsis thaliana | - | - | 0 | 0.00001 | [35, 36] | ||
| 1-deoxyxylulose 5-phosphate synthase ↗ (plast.) | Arabidopsis thaliana | ||||||||
| Artemisinin | antimalarial treatment | squalene synthase ↘ (antisense) | Artemisia annua | Artemisia annua | - | - | 1.0 | 3.1 | [43] |
| AaORA ↗ (nuc.) | Artemisia annua | Artemisia annua | - | - | 0.8 | 1.2 | [44] | ||
| (AP2/ERF transcription factor) | |||||||||
| AaERF1 and AaERF2 (nuc.) | Artemisia annua | Artemisia annua | - | - | 0.5 | 0.8 | [45] | ||
| (AP2/ERF transcription factors) | |||||||||
| amorpha-4,11-diene synthase ↗ (mit.) | Artemisia annua | Nicotianatabacum | - | - | 0 | 0.0007 | [48] | ||
| amorphadiene oxidase ↗ (ER), | Artemisia annua | ||||||||
| cytochrome P450 reductase (CPR) ↗ (ER), | Artemisia annua | ||||||||
| artemisinicald.Δ11(13) double-bond red. ↗ (cyt.), | Artemisia annua | ||||||||
| truncated HMG-CoA red. ↗ (cyt.) | Sacch. cerevisiae | ||||||||
| Amorpha-4,11-diene | precursor for artemisinin | amorphadiene synthase ↗ (mit.), | Artemisia annua | Nicotianabenthamiana | - | - | 0 | 0.02 | [47] |
| (antimalarial treatment) | farnesyl-PP synthase ↗ (mit.) | Arabidopsis thaliana | |||||||
| truncated HMG-CoA reductase ↗ (cyt.) | Sacch. cerevisiae | ||||||||
| amorphadiene synthase ↗ (plast.), | Artemisia annua | Menthax piperita | 0 | 4.9 | 0 | 0.1 | Suppl. Info | ||
| farnesyl-PP synthase ↗ (plast.) |
Mentha x piperita |
||||||||
Table 1: Metabolic engineering efforts with aromatic plants (and non-aromatic plants for reference).
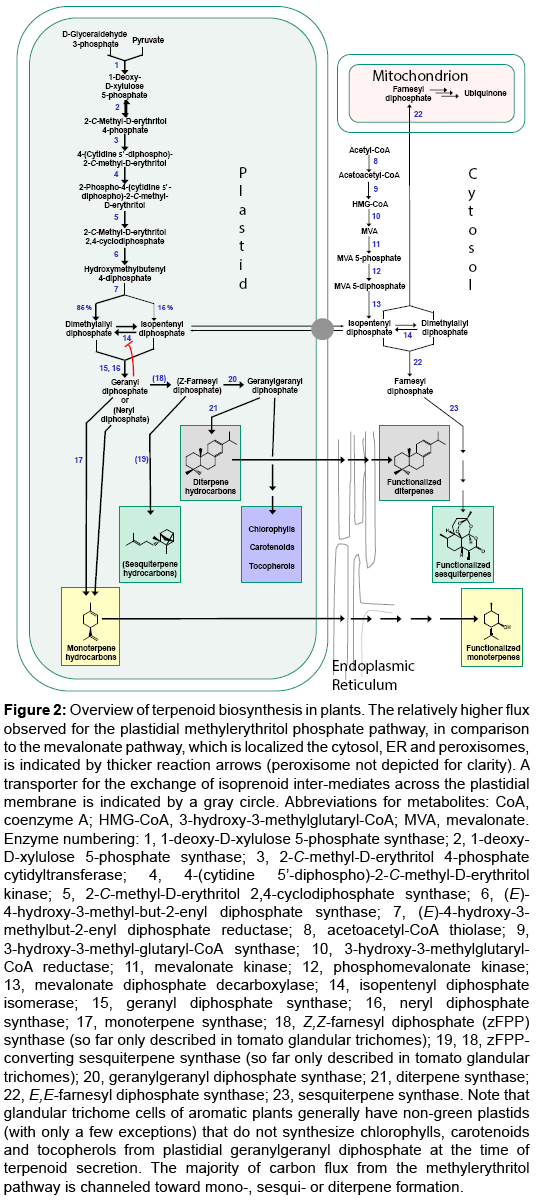
Figure 2: Overview of terpenoid biosynthesis in plants. The relatively higher flux observed for the plastidial methylerythritol phosphate pathway, in comparison to the mevalonate pathway, which is localized the cytosol, ER and peroxisomes, is indicated by thicker reaction arrows (peroxisome not depicted for clarity). A transporter for the exchange of isoprenoid inter-mediates across the plastidial membrane is indicated by a gray circle. Abbreviations for metabolites: CoA, coenzyme A; HMG-CoA, 3-hydroxy-3-methylglutaryl-CoA; MVA, mevalonate. Enzyme numbering: 1, 1-deoxy-D-xylulose 5-phosphate synthase; 2, 1-deoxy- D-xylulose 5-phosphate synthase; 3, 2-C-methyl-D-erythritol 4-phosphate cytidyltransferase; 4, 4-(cytidine 5’-diphospho)-2-C-methyl-D-erythritol kinase; 5, 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; 6, (E)- 4-hydroxy-3-methyl-but-2-enyl diphosphate synthase; 7, (E)-4-hydroxy-3- methylbut-2-enyl diphosphate reductase; 8, acetoacetyl-CoA thiolase; 9, 3-hydroxy-3-methyl-glutaryl-CoA synthase; 10, 3-hydroxy-3-methylglutaryl- CoA reductase; 11, mevalonate kinase; 12, phosphomevalonate kinase; 13, mevalonate diphosphate decarboxylase; 14, isopentenyl diphosphate isomerase; 15, geranyl diphosphate synthase; 16, neryl diphosphate synthase; 17, monoterpene synthase; 18, Z,Z-farnesyl diphosphate (zFPP) synthase (so far only described in tomato glandular trichomes); 19, 18, zFPPconverting sesquiterpene synthase (so far only described in tomato glandular trichomes); 20, geranylgeranyl diphosphate synthase; 21, diterpene synthase; 22, E,E-farnesyl diphosphate synthase; 23, sesquiterpene synthase. Note that glandular trichome cells of aromatic plants generally have non-green plastids (with only a few exceptions) that do not synthesize chlorophylls, carotenoids and tocopherols from plastidial geranylgeranyl diphosphate at the time of terpenoid secretion. The majority of carbon flux from the methylerythritol pathway is channeled toward mono-, sesqui- or diterpene formation.
Conclusions
Various genes encoding monterpene synthases have been overexpressed in aromatic plants for the increased accumulation of C10 hydrocarbons in essential oils, but the yields achieved thus far have been fairly low and further optimization is necessary. Tobacco is an emerging experimental system for the biotechnological production of diterpenes. The most promising approach for hydrocarbon accumulation appears to be the downregulation of the expression of endogenous genes that encode enzymes responsible for early oxygenation reactions.
Metabolites with relevance for the agribusiness industry
Oleoresins and essential oils of aromatic plants contain various bioactive metabolites, some of which repel insects [26]. This bears the potential of transferring genes involved in the biosynthesis of insect repellent metabolites into crop plants, thus potentially increasing disease resistance. Wu et al. [27] demonstrated that the expression of patchoulol synthase (PTS) from patchouli (Pogostemon cablin (Blanco) Benth.), using a constitutive promoter, in transgenic tobacco plants resulted in the low-level accumulation of the sesquiterpene patchoulol (8) (0.005% of dry weight biomass) [27]. The authors hypothesized that the precursor supply for the cytosolic PTS was limiting and a plastidial targeting of relevant enzyme activities could potentially increase sesquiterpene levels. Plastidial targeting sequences were thus added to the genes encoding farnesyl diphosphate (FPP) synthase and PTS, and both of these modified genes were fused to strong constitutive promoters. Transgenic tobacco plants with plastidial FPP synthase and PTS activities accumulated patchoulol at 0.015% of dry weight biomass [27], indicating that the plastid indeed appeared to support higher flux through the terpenoid pathway. It is important to note, however, that high patchoulol producers exhibited chlorotic symptoms and had reduced biomass, indicating that sesquiterpene production may have interfered with other plastidial terpenoid pathways involved in chlorophyll and carotenoid biosynthesis. Interestingly, transgenic patchoulol producers were significantly less affected by tobacco hornworm feeding when compared to wild-type controls, indicating that controlled patchoulol synthase expression has the potential to be used for generating more insect-resistant crop plants [27]. Wang et al. [28] showed that the suppression of the gene encoding a cytochrome P450-dependent monooxygenase (CYP71D16) in transgenic tobacco, by both antisense technology and cosuppression, resulted in a significant decrease in the amounts of the predominant diterpene cembratriene-diol (9) and a 19-fold increase of its biosynthetic precursor, cembratriene-ol (10), to 4.3% of dry weight biomass [28]. This is the highest accumulation level of an engineered terpenoid reported thus far and the work provides additional evidence for the notion that the downregulation of endogenous genes that encode oxygenating enzymes is currently the most promising bioengineering approach towardincreasing the accumulation of terpenoid hydrocarbons. They also found that suppression of CYP71D16 significantly reduced the plant’s resistance to colonization by the tobacco aphid (Myzus nicotiana) [28].
Conclusions
The cytosolic precursor pathway for terpenoids appears to be highly regulated, and terpenoid precursors generally appear to be more abundant or available in plastids [29] (Figure 2). Therefore, using metabolic engineering constructs for the plastidial localization of terpenoid biosynthetic enzymes in transgenic plants is a viable strategy. However, high activities of introduced enzymes can interfere with essential plastid-localized terpenoid pathways, and a balancing of fluxes is thus required. The accumulation of specific terpenoids, by expressing genes from aromatic plants in transgenic crops, has significant potential for controlling insect pests, but more research is needed to test if this approach is more widely applicable.
Metabolites with relevance for the pharmaceutical industry
Thousands of metabolites extracted from aromatic plants have been evaluated for their medicinal properties. Many of these metabolites have shown promising biological activities in in vitro assays and early stage clinical trials, but only a few have entered advanced-stage clinical trials. Even fewer have been introduced to the pharmaceutical market. However, successfully commercialized drugs developed from aromatic plant metabolites have had a transformative impact on the industry.
The diterpene taxol® (generic name paclitaxel) (11) and its derivatives have been the best-selling antitumor agents for more than a decade [30]. Taxol differs from the other metabolites discussed in this review because it is not produced by specialized glandular cells and it does not accumulate within secretory ducts, cavities, or trichomes [31], but it is an extensively researched, medicinally important, terpenoid derivative that accumulates to only moderate levels within the secondary phloem of yew trees and has thus been a target for biotechnological production. The development of taxol as a drug has a history that illustrates generalizable challenges with plant natural products. During the clinical trial phases, taxol was extracted from bark strips of Pacific yew, a process that was soon recognized to be unsustainable [32]. A semi-synthetic route, which combined the extraction of a biosynthetic precursor, baccatin III (12), from leaves of the European yew (Taxus baccata L.) with subsequent modifications by chemical synthesis, was developed and adjusted for the commercial production of taxol. An alternative biotechnological approach, which employed Taxus cell suspension cultures, was developed by scientists at the US Department of Agriculture and later optimized for an industrial scale by Phyton Inc. [32]. Despite these successes, the market price of taxol is still very high and additional biotechnological approaches have been explored. Taxus lacks secretory structures associated with taxol production and the metabolite is accumulated within cell walls of the secondary phloem, vascular cambium, and xylem [31]. Tobacco leaves are densely populated with long glandular trichomes that synthesize an exudate rich in diterpenes (up to 10% of leaf dry weight in the experimental variety Nicotiana tabacum T.I. 1068) [33]. Tobacco is thus an attractive host for engineering diterpenoid biosynthesis, including the production of taxol precursors. Tissier et al. [34] expressed the gene encoding taxadiene synthase, the enzyme that catalyzes the first committed step of the taxol biosynthetic pathway, in transgenic tobacco (Nicotiana sylvestris Speg. &Comes). Relatively small amounts of taxa-4(5),11(12)-diene (13) (up to 0.005% of dry weight biomass) were obtained when either the 35S (constitutive) or the glandular trichome-specific NsTPS-02a promoters were used to drive transgene expression (Table 1). As a reference, taxa- 4(5),11(12)-diene accumulation in leaves of transgenic Arabidopsis thaliana (L.) plants, which do not have glandular trichomes, was even lower (0.00001% of dry weight biomass) when transformed with 35S promoter-driven constructs for the overexpression of the genes coding for 1-deoxy-D-xylulose 5-phosphate synthase (an enzyme involved in the synthesis of terpenoid precursors) and taxadiene synthase [35,36]. Interestingly, when Anterola and colleagues expressed taxadiene synthase constitutively in the moss Physcomitrella patens (Hedw.) Bruch & Schimp., taxa-4(5),11(12)-diene levels were as high as 0.2%, with apparently no negative effects on fitness [37] (Table 1).
Conclusions
Thus far, genes involved in only the early steps of the taxol biosynthetic pathway, which requires about 20 different enzymes [38], have been transformed into heterologous plant or microbial hosts [39]. Based on the very small number of published studies, aromatic plants may offer advantages over plants lacking glandular trichomes. Surprisingly, the experimental plant model system with the highest accumulation of the taxol precursor taxa-4(5),11(12)-diene, following transformation with the taxadiene synthase gene, was the moss Physcomitrella. It will now be important to assess if functionalized terpenoids can be accumulated at high levels in this moss, or if the expression of additional taxol pathway genes leads to dramatic yield penalties as described for engineered E. coli [39].
Artemisinin (Chinese: Qinghaosu; ???) (14), a sesquiterpene lactone occurring in the glandular trichomes of sweet wormwood (Artemisia annua L.), has become a critically important component of anti-malarial therapies [40]. To address the problem of supply shortages and quality inconsistencies, the Bill and Melinda Gates Foundation funded the development of improved Artemisia cultivars, which now accumulate artemisinin at up to 3% of dry weight biomass [41], and engineered microbial strains that accumulate a precursor for the semi-synthetic artemisinin production [42]. Independently, the utility of generating transgenic plants for this purpose has also been evaluated. An antisense-downregulation of the expression of the gene encoding squalene synthase, which is involved in the biosynthesis of triterpenes and competes for sesquiterpenoid pathway precursors, led to transgenic Artemisia plants with three-fold higher accumulation of artemisinin (3.1% of dry weight biomass), compared to untransformed controls [43] (Table 1). An alternative approach to overexpressing genes encoding structural proteins of a target biosynthetic pathway is the use of genes coding for regulatory proteins. The overexpression of the AP2/ERF transcription factor genes AaORA, AaERF1, and AaERF2 led to a more than 60% increase in artemisinin production in Artemisia [44,45]. The overexpression of the AaWRKY1 transcription factor gene in Artemisia resulted in elevated transcript levels for several genes involved in artemisinin biosynthesis but, unfortunately, the metabolite itself was not quantified [46]. Van Herpen and colleagues engineered tobacco (Nicotiana benthamiana Domin.) for the accumulation of amorpha-4,11-diene (15), a precursor of artemisinin biosynthesis [47]. The most successful construct contained a truncated 3-hydroxy-3-methylglutaryl-CoA reductase gene from Artemisia (gene product accumulates in cytosol), a modified FPP synthase gene from Arabidopsis (gene product accumulates in mitochondria), and a modified amorphadiene synthase gene from Artemisia (gene product accumulates in mitochondria). Transiently transformed tobacco leaves accumulated amorpha-4,11-diene to 0.02% of leaf dry weight [47] (Table 1), indicating that mitochondria are a suitable location for enzymes in transgenic plants engineered for terpenoid formation. Our laboratory expressed modified FPP synthase and amorphadiene synthase genes (both gene products targeted to plastids) in transgenic peppermint, where amorpha-4,11-diene accumulated to 0.1% of dry weight biomass (Table 1; Supplementary Information). The transfer of the entire artemisinin biosynthetic pathway into transgenic tobacco plants was recently achieved by Farhi et al. [48]. Five genes, each controlled by a different constitutive promoter, were placed on a single mega vector for transformation, and an artemisinin yield of 0.0007% of dry weight biomass was achieved [48].
Besides artemisinin, two additional sesquiterpene lactones have attracted the attention of clinical researchers: the parthenolide derivative LC-1 (16) and thapsigargin (17). These compounds are currently being tested in phase I clinical trials for the treatment of blood-lymph and advanced solid tumors, respectively [49]. Other examples include several cannabinoid preparations (Δ9-tetrahydrocannabinol (18) shown as an example in Figure 1), which have been introduced for the management of chronic pain [50]. Perillyl alcohol (19) has shown promising antitumor activities in in vitro assays, but chemopreventative effects were not confirmed in phase II clinical trials [51]. No metabolic engineering efforts for the biotechnological production of any of these aromatic plant-derived targets have been reported yet.
Conclusions
Metabolic engineering efforts with the native host (Artemisia) have been more successful for high level artemisinin production than those using heterologous hosts. The entire artemisinin biosynthetic pathway has been successfully transferred to tobacco, which was a demonstration of the feasibility of complex metabolic engineering approaches at the proof-of-concept stage, but the overall yields were disappointingly low. Metabolic precursors of artemisinin have been accumulated in heterologous hosts (Arabidopsis and peppermint) at much higher levels when appropriate biosynthetic enzymes were targeted to plastids or mitochondria. These results indicate that the supply of precursors is likely one of the limiting factors for producing pharmaceutically relevant sesquiterpenes in aromatic plants.
Unexpected outcomes of terpenoid metabolic engineering efforts
One of the common problems with transgenic host plants, which often do not have oil storing secretory structures, is that terpenoids are volatilized and, thus, cannot be collected efficiently on a commercial scale [14]. A second challenge is the propensity of transgenic plants to further metabolize novel terpenes, particularly when the gene introduced by metabolic engineering is expressed constitutively. Negative results or those that are difficult to explain are often not published in the primary literature. However, an appreciation of which approaches did not yield anticipated or desired outcomes is important for designing metabolic engineering strategies. We are thus including in this paragraph some as yet unpublished data from our own work that we believe are instructive in the context of this review
Lücker et al. [52] expressed a linalool synthase gene from Clarkia breweri in transgenic Petunia × hybrid plants and observed that the majority of S-linalool was converted to the corresponding glucoside (20). When the same gene was expressed in tobacco (Solanum lycopersicum L.), 8-hydroxylinalool (21) was detected [53], while carnation (Dianthus caryophyllus L.) accumulated linalool oxide (22) [54]. Aharoni et al. [55] reported that the expression of a multifunctional linalool/nerolidol synthase (FaNES) gene from strawberry (Fragariax ananassa; plastidial targeting of gene product) led to the formation of the monoterpene S-linalool, but also hydroxylated and glucosylated linalool metabolites in both Arabidopsis [55] and potato tubers [56]. The sesquiterpene S-nerolidol, in contrast, was only produced at trace levels, indicating that FPP, the precursor for sesquiterpene biosynthesis, was limiting in plastids. Plants that expressed FaNES at high levels were affected by stunted growth. The likely cause for this phenotype, as discussed by the authors, was a redirection of plastidial terpenoid precursors to monoterpene production, which depleted pools for the biosynthesis of other plastidial metabolites such as chlorophylls, carotenoids, and growth regulators [55] (Figure 2). To avoid interfering with plastidial terpenoid metabolism, Kappers et al. [57] transformed Arabidopsis with a construct for the accumulation of the FaNES gene product in mitochondria. Contrary to the results obtained with transgenic plants containing a plastidial FaNES activity, mitochondrial targeting of the enzyme led to the accumulation of S-nerolidol (23) but not S-linalool [57]. These results suggest that Arabidopsis mitochondria can naturally provide sufficient precursor for sesquiterpene but not for monoterpene biosynthesis. Interestingly, the mitochondrial localization of an FaNES activity also led to the formation of another new metabolite, the hemiterpene 4,8-dimethyl-1,3,7-nonatriene (24), in transgenic Arabidopsis plants [57]. The originally formed S-nerolidol appears to have been further metabolized by an endogenous activity, and enzymes acting on this metabolite, such as the ER-localized cytochrome P450 monooxygenase CYP82G1, have been described in the literature [58]. Kappers and colleagues then demonstrated that emitted S-nerolidol and 4,8-dimethyl-1,3,7-nonatriene were attractants for predatory mites [57], and transgenic plants might thus be able to “call upon” beneficial insects to control infestations with arthropods. Although not originally planned, the metabolic turnover of S-nerolidol had desirable effects on the pest resistance of transgenic plants.
Occasionally, metabolic engineering leads to the upregulation of enzymes unrelated to the target pathway. For example, we found that some transgenic peppermint lines transformed with two genes in order to generate a plastidial pool of amorpha-4,11-diene (precursor of the antimalarial sesquiterpene lactone artemisinin) accumulated high levels (up to 48% of the total essential oil) of (+)-menthofuran (25), a side product of the peppermint monoterpene pathway (Table 2; Supplementary Information). Similarly, the expression of a sabinene synthase from sage (Salvia officinalis) resulted in some transgenic lines with vastly elevated levels (up to 11% of the total essential oil) of (+)-pulegone (26), another intermediate of the peppermint monoterpene pathway (Table 2; Supplementary Information). The transgenic peppermint lines with elevated (+)-menthofuran or (+)-pulegone levels had a purple coloration akin to that described for plants accumulating stress-related anthocyanins [59]. We hypothesize that the observed changes in essential oil composition are the result of unspecific stress responses caused by the transformation process, but further research is necessary to elucidate the cause(s) of these surprising findings.
| Target metabolite | Utility | Gene(s) | Source plant | Host plant | Amount of target metabolite | References | |||
|---|---|---|---|---|---|---|---|---|---|
| of gene | for transformation | [% of oil or exudate] | [% of dry weight biomass] | ||||||
| CONTROL | TRANSGENIC | CONTROL | TRANSGENIC | ||||||
| S-Linalool | S-Linalool glucoside | linalool synthase ↗ (plast.) | Clarkia breweri |
Petunia hybrida |
- | - | 0 | 0.01 | [52] |
| 8-hydroxylinalool | linalool synthase ↗ (plast.) | Clarkia breweri |
Solanum lycopersicum |
0 | 0.00003 | [53] | |||
| Linalool oxide | linalool synthase ↗ (plast.) | Clarkia breweri |
Dianthus caryophyllus |
- | - | n.q. | [54] | ||
| S-Linalool, (3S)-Enerolidol |
8-hydroxylinalool, | linalool synthase ↗ (plast.) | Fragaria x ananassa |
Arabidopsis thaliana |
- | - | 0 | 0.1 | [55] |
| 8-Hydroxy-6,7- dihydrolinalool |
|||||||||
| terpene alcohol glucosides |
|||||||||
| 8-hydroxylinalool, | linalool/nerolidol synthase ↗ (plast.) |
Fragaria x ananassa |
Solanum tuberosum |
- | - | n.q. | n.q. | [56] | |
| 8-Hydroxy-6,7- dihydrolinalool |
|||||||||
| terpene alcohol glucosides |
|||||||||
| 4,8-dimethyl-1,3,7- nona-triene |
linalool/nerolidol synthase ↗ (plast.) |
Fragaria x ananassa |
Arabidopsis thaliana |
- | - | n.q. | n.q. | [57] | |
| (-)-Limonene, γ-humulene |
(-)-limonene | γ-humulene synthase ↗ (cyt.) |
Abies grandis | Mentha x piperita |
1.3 | 1.3 | 0.01 | 0.9 | Suppl. Info |
| limonene 3-hydroxylase ↘ (cosuppression) |
n.a. | ||||||||
| Amorpha- 4,11-diene |
(+)-menthofuran | amorphadiene synthase ↗ (plast.), |
Artemisia annua |
Mentha x piperita |
3.9 | 47.8 | 0.04 | 0.5 | Suppl. Info |
| farnesyl-PP synthase ↗ (plast.) |
Mentha x piperita |
||||||||
| (+)-Sabinene | (+)-pulegone | sabinene synthase ↗ (plast.) |
Salvia officinalis |
Mentha spicata (Erospicata) |
0.6 | 11.3 | 0.02 | 0.4 | Suppl. Info |
| Taxa- 4(20),11(12)- diene-5-ol |
5(12)-Oxa-3(11)- cyclotaxane |
taxadiene synthase ↗ (plast.), |
Taxus brevifolia, |
Nicotiana sylvestris |
- | - | 0 | 0.0002 | [61] |
| CYP725A4 ↗ (ER) | Taxus cuspidata |
||||||||
Table 2: Examples of unexpected outcomes of terpenoid engineering efforts with aromatic plants (and non-aromatic plants for reference).
Enzyme catalytic plasticity can cause additional unexpected results when employing metabolic engineering efforts. When we transformed a transgenic peppermint line with increased amounts of the monoterpene pathway precursor (-)-limonene (79% of the total essential oil) [21] with an additional gene encoding γ-humulene synthase from Grand fir (Abies grandis L), the sesquiterpene γ-humulene (27) did not accumulate to detectable levels. Instead, the monoterpene (-)-limonene accumulated to even higher levels (up to 90% of the total essential oil) (Table 2; Supplementary Information). Steele et al. had reported that γ-humulene synthase converts geranyl diphosphate to (-)-limonene, albeit with fairly low efficiency, in in vitro assays [60], but we had assumed that the available amounts of the monoterpene precursor geranyl diphosphate would be negligible in the cytosol, which is where the enzyme is localized. The observed increases in (-)-limonene concentration in transgenic plants could be caused by a side activity of the introduced γ-humulene synthase or an upregulation of limonene synthase, but further experimentation is needed to test these hypotheses. In an effort to produce a precursor for taxol biosynthesis, Rontein and colleagues transformed tobacco (Nicotiana sylvestris L.) with the genes encoding taxadiene synthase from Taxus brevifolia and CYP725A4 from Taxus cuspidata [61]. However, instead of the expected taxa-4(20),11(12)-dien-5α-ol (28), transgenic plants accumulated 5(12)-oxa-3(11)-cyclotaxane (29) [61], a metabolite that had not been observed in previous in vitro assays with recombinant CYP725A4 [62]. Both metabolites were later reported to occur in E. coli engineered for the expression of the same heterologous genes [39]. Taken together, these examples illustrate the uncertainties currently associated with terpenoid pathway engineering in plants, and serve as a reminder that more effort needs to be devoted to develop platform hosts in which introduced metabolic pathways operate more predictably than currently achievable.
Future directions of aromatic plant biotechnology
The constitutive expression of transgenes with the concomitant accumulation of novel enzyme activities can result in a competition for precursors that are also used in essential pathways. For example, when terpene synthases were expressed constitutively in transgenic plants, interference with the biosynthesis of sterols, chlorophylls or carotenoids has led to severe adverse effects ranging from dwarf phenotypes to bleaching [27,36,56,57]. Many aromatic plants accumulate essential oils and resins in specialized secretory (or glandular) structures, which allow for the localized accumulation of large quantities of secondary metabolites, while avoiding interference with primary pathways essential for plant growth. Thus, it would be desirable to biotechnologically introduce pathways to the glandular cells of these structures. An additional advantage of targeting secretory structures for metabolic engineering relates to the remarkable specialization of their cells for allowing high fluxes toward specific terpenoid or phenylpropanoid end products [12]. One engineering approach would be to employ glandular trichome-specific promoters, which have been isolated and characterized from various plants (reviewed in [63]). However, so far, the utility of these promoters for metabolic engineering of glandular trichome metabolism has been evaluated in only a couple of cases by the Tissier laboratory. They used the cembratrienol synthase (CBTS) promoter, to direct the expression of the NtCPS2 and NtABS genes to glandular trichomes of transgenic flowering tobacco plants, which accumulated the diterpene Z-abienol at fairly high levels (0.018% of dry weight biomass) [25]. In another study, Rontein et al. [61] expressed taxadiene synthase and CYP725A4 under control of the glandular trichome-specific CBTS and CYP71D16 promoters, respectively, but only small amounts of a cyclic taxane were produced in transgenic flowering tobacco. At a proof of concept level, glandular trichome-specific promoters confer cell type specificity and relatively high levels of transgene expression. Systematic studies, comparing glandular trichome-specific and constitutive promoter’s side-by-side for the purpose of obtaining high amounts of particular target metabolites, are now needed to evaluate the true potential of cell type-specific promoters for plant metabolic engineering.An attractive approach to upregulate the expression of several genes within a metabolic pathway is the overexpression of a master regulator, such as a transcription factor. Transcription factors with demonstrated attenuating effects on glandular trichome-specific pathways have been characterized in Artemisia, where the ectopic overexpression of AaORA, AaERF1 or AaERF2 resulted in increased expression levels of several genes involved in sesquiterpene biosynthesis and an associated increase in artemisinin concentrations [44,45]. It should be noted, however, that studies reporting high artemisinin yield increases [43-45] used Artemisia cultivars in which the artemisinin concentration after engineering was at best comparable to that reported for high natural producers [41].
Desirable effects on glandular trichome metabolism are potentially achievable by the ectopic expression of genes that affect relevant developmental programs. For example, the constitutive expression of NtLTP1 (lipid transfer protein required for secretion of hydrophobic materials), PIN2 (proteinase inhibitor) or GL1 (myb transcription factor) led to increases in glandular trichome density in transgenic tobacco [64-66]. Choi et al. [64] demonstrated that NtLTP1 expression increased the amounts of cembranoid and labdanoid diterpenes and alkanes, which indicates that, in additional to effects of glandular trichome development, the terpenoid pathway was also affected in transgenic plants [64].
We observed that, surprisingly, the antisense-based downregulation of the gene encoding menthofuran synthase, which catalyses a side branch reaction of the monoterpene pathway in peppermint, resulted in increased glandular trichome density and a shift toward earlier essential oil secretion, concomitant with significant increases (35%) in monoterpene formation [67]. This example indicates that modulating the developmental patterning programs of glandular trichomes has the potential of increasing the production of target metabolites in aromatic plants. However, the mechanisms underlying these intriguing findings are currently not understood and future research should be aimed at unraveling processes that can be used to enhance the capacity of engineered aromatic plants for accumulating valuable chemical and pharmaceutical feedstocks.
Essential oils and resins contain terpenoids as major components. The biosynthesis of this class of metabolites in plants is highly compartmentalized (Figure 2) and several metabolic engineering strategies have been developed to increase flux into target pathways. The mevalonate pathway for terpenoid precursor biosynthesis, which is localized in the cytosol endoplasmic reticulum and peroxisomes, appears to be highly regulated [29], and the targeting of biotechnologically introduced enzymes, such as terpene synthases, to the cytosol generally does not allow for the production of larger amounts of novel terpenoids [20,27]. The targeting of sesquiterpene synthases and FPP synthase (which are mostly localized to the cytosol and/or mitochondria) to plastids has led to much higher accumulation levels of sesquiterpenoids. If the transgenes are expressed constitutively at high levels, the accumulation of the corresponding enzymes in plastids can lead to adverse effects due to competition for precursors of chlorophyll and carotenoid biosynthesis [27,36]. No negative effects on plant fitness have as yet been reported for terpene synthases and additional FPP synthase copies that were targeted to mitochondria of transgenic plants [47,48]. However, further metabolic conversions of mitochondrially formed terpenoid products have been described [57], indicating that, although high terpenoid production levels are achievable by this approach, caution should be exerted to avoid undesirable effects. The least amount of metabolic turnover of chemical or pharmaceutical targets produced in aromatic plants appears to occur when the relevant pathways operate exclusively in glandular trichomes [14]. Research is needed to help us understand how to combine cell type specificity, appropriate transgene expression levels and favourable subcellular compartmentation in future metabolic engineering efforts.
In summary, aromatic plants have already been demonstrated to hold promise for accumulating valuable chemical and pharmaceutical feedstocks. It will now be important to develop an arsenal of tools, including efficient transformation protocols, glandular trichomespecific promoters and appropriate regulatory factors, which can be employed in metabolic engineering to more effectively modulate glandular trichome patterning and pathway flux balancing. In combination, such tools have the potential to enable the production of commercially relevant amounts of target compounds in aromatic plants.
The work presented here was supported by the Division of Chemical Sciences, Geosciences, and Biosciences, Office of Basic Energy Sciences, U.S. Department of Energy (DE-FG02-09ER16054), the Mint Industry Research Council, and the Washington Research Foundation. The authors would like to thank Lyuba Yurgel and William Cuthbertson for technical assistance and the greenhouse staff at the Institute of Biological Chemistry (Amy Hetrick, Susan Vogtman and Juliana Gothard-Szamosfalvi) for maintaining plant lines. We thank Dr. Rodney Croteau (WSU) for the donation of plasmids containing the sabinene synthase, pinene synthase, linalool synthase and γ-humulene synthase genes. We would also like to thank Dr. Peter Brodelius (University of Lund) for the donation of the amorphadiene synthase-containing plasmid.